光合作用(卷名:生物学)
photosynthesis
━━━━━━━━━━━━━━━━━━━━━━━━
光合作用的发现
光合作用的机构及其演化
光合作用的机理
原初反应
同化力形成
光合电子传递
光合磷酸化
碳同化
光呼吸
四碳途径和景天科酸代谢
光合作用的效率及其影响因素
光合作用和植物生理状态的关系
光合作用和环境条件的关系
光
温度
水
气
肥
群体光能利用━━━━━━━━━━━━━━━━━━━━━━━━
植物利用光能将二氧化碳和水等无机物合成有机物并放出氧气的过程。通常以下式表示:
 式中(C啹O)代表合成的以碳水化合物为主的有机物。
式中(C啹O)代表合成的以碳水化合物为主的有机物。光合作用中合成的有机物是植物赖以生长的主要物质来源和全部能量来源,也是其他直接或间接依靠植物生活的生物的有机物和能量来源。光能及化能自养细菌可行光合作用的部分过程,也能合成有机物,但总量极微。地球上的植物每年通过光合作用合成近2000亿吨有机物,同时固定了3×1021焦耳的太阳能,相当于人类全部能耗的10倍。地层中埋藏的煤炭、石油和天然气是古代植物光合作用形成的有机物演变而成的。光合作用释放O2和固定CO2,使大气中的O2积累,CO2含量降低。
光合作用对地球演化、生物进化、现有大气环境的维持及人类的生活和生产都有非常重大的意义。人们除从各方面探讨和利用光合作用外,还模拟它的部分过程,借以研究光合作用的机理和对其的仿生应用。
光合作用的发现
古希腊哲学家亚里士多德认为,植物生长所需的物质全来源于土中。荷兰人范·埃尔蒙做了盆栽柳树称重实验,得出植物的重量主要不是来自土壤而是来自水的推论。他没有认识到空气中的物质参与了有机物的形成。在此前后,中国明末的宋应星在《论气》一书中说:“气从地下催腾一粒,种性小者为蓬,大者为蔽牛干霄之木,此一粒原本几何?其余则皆气所化也。”明确认识到植物(以及动物)身体的物质是由气转化而来。英国J.普里斯特利在1771年发现植物可以恢复因蜡烛燃烧而变“坏”了的空气。1773年荷兰J.英恩豪斯证明只有植物的绿色部分在光下才能起使空气变“好”的作用。1804年瑞士N.-T.de索绪尔通过定量研究进一步证实CO2和水是植物生长的原料。1845年德国J.R.迈尔发现植物把太阳能转化成了化学能。1860年左右,人们就已经用
表示植物利用光能的总过程。1897年首次在教科书中称它为光合作用。20世纪30年代,C.B.范尼尔发现有些细菌可在无氧条件下利用光能进行与光合作用类似的反应。但它们是从硫化氢等而不是从水取得还原二氧化碳的氢,也不释放氧气。他把这个反应称为细菌光合作用。此外,人们还发现有些细菌可通过氧化一些无机物获得能量进行有机物的合成反应,其过程和光合作用有许多类似之处,被称为化能合成作用。
光合作用的机构及其演化
光合作用是植物特有的功能。从藻类到被子植物,进行光合作用的机构有很多共同之处,但也存在着差别。植物的绿色部分都能进行光合作用。通常叶子是植物主要的光合器官。它形状扁平,有利于捕捉光能;表皮上有气孔,内部叶肉细胞间有不少空隙,便于气体交换;叶片内叶脉纵横,可进行频繁的水分及光合产物的运输。在具有真核细胞、能进行光合作用的植物 (包括大部分藻类)的细胞中,均有专门的细胞器──叶绿体。它外面包有双层被膜,以维持叶绿体内部特别适于光合作用进行的微环境;内部含大量扁平密闭的成囊状的膜片层──类囊体及无明显结构的间质。类囊体叠合成堆的部分称基粒。在原核生物蓝藻和原绿藻中,类囊体外无被膜,直接悬浮于细胞质内。主要的光合色素──叶绿素 a全部集中在类囊体上,在高等植物和一些藻类(如原绿藻和绿藻)中,类囊体上还有叶绿素b及类胡萝卜素;其他藻类有的无叶绿素b而有叶绿素c等;蓝藻、红藻则在类囊体表面有藻胆素。这些色素吸收的光能也能用于光合作用。
60年代以后,人们确证了叶绿体中存在着与其遗传有关的脱氧核糖核酸及其复制、转录、翻译系统,它们的性质和原核生物的类似。因此有人认为植物中的叶绿体是能营光合作用的类似原绿藻、蓝藻的原始原核生物进行真核细胞后内部共生、演化而成的。
关于光合作用的发生和演化,一般认为是于太古代的原始海洋中,在太阳辐射等作用下形成了一些非生命来源的有机物,经过长期相互作用变化,产生了原始生物,其中有的演化出了可利用光能合成有机物的功能,类似细菌光合作用;从中再逐渐演化出类似原绿藻和蓝藻那样能营放氧光合作用的原核生物;以后才出现具有叶绿体的营放氧光合作用的植物。
光合作用的机理
光合作用是一个很复杂的过程,它至少包含几十个步骤,大体上可分为原初反应、同化力形成和碳同化 3大阶段。原初反应包括光能的吸收、传递和电荷的分离;同化力形成是原初反应所引起的电荷分离,通过一系列电子传递和反应转变成生物代谢中的高能物质腺苷三磷酸(ATP)和还原辅酶Ⅱ(NADPH);碳同化是以同化力(ATP和NADPH)固定和还原 CO2形成有机物质。它们之间的衔接及与叶绿体结构的联系见图。
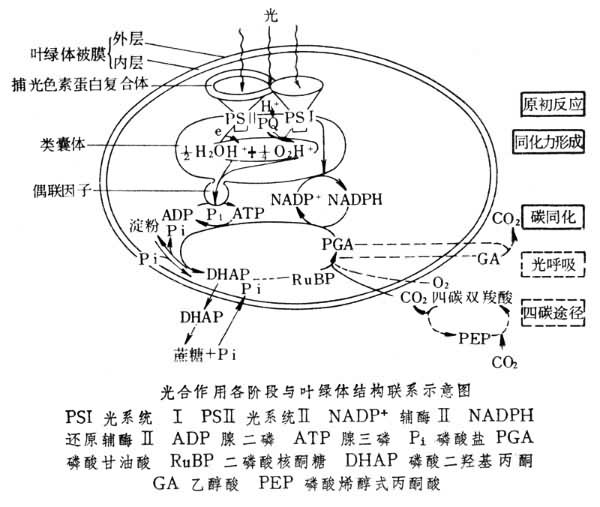
原初反应 光能被光合色素吸收、传递至反应中心并发生光化学反应,引起电荷分离。这些过程在1纳秒内就已完成。在类囊体膜上光合色素与蛋白质组成复合体,这些复合体中的光合色素可吸收不同波长的光能,但只有少数在反应中心处于特殊状态的叶绿素 a可进行光化学反应,其余色素所吸收的光能要传递给反应中心的叶绿素 a才有用。这些仅能吸收及传递光能不能引起电荷分离等光化学反应的色素起了类似无线电天线的收集能量的作用,称天线色素。一般情况下250~300个天线叶绿素分子和一个反应中心构成一个光合单位。将类囊体膜用机械的或化学的方法破碎,可分离出组成不同的 3大类色素蛋白复合体颗粒。其中两类能进行不同的光化学反应:一类颗粒的反应中心叶绿素a在发生光化学反应时,其吸收变化的峰值在700纳米左右,称光系统Ⅰ颗粒;另一类颗粒的反应中心叶绿素a进行光化学反应时,其吸收变化的峰值在680纳米左右,称光系统Ⅱ颗粒。第三类颗粒只含天线色素(在植物及绿藻等中包含部分叶绿素a 及绝大部分叶绿素 b,在蓝藻及红藻中则含全部藻胆素),称集光色素蛋白复合体,它吸收的光能可传与光系统Ⅱ也可传与光系统Ⅰ,其在两个光系统间的分配比例受一系列因素,尤其是膜表面所带的电荷特性的调节。
从光合细菌中,已分离到能进行光化学反应的色素蛋白复合体,将它的天线色素除去后,可得到纯净的反应中心。后者由 4个细菌叶绿素a及2个去镁细菌叶绿素a组成。
在这些色素蛋白复合体中,光合色素的排列很紧密,色素分子吸收光能后所产生的受激态可有效地通过共振传递给其他色素分子,最后传到反应中心,发生光化学反应(未传到反应中心的则变成荧光而散发或变为热能而耗散),即反应中心的叶绿素a把一个电子交给原初电子受体,使它带负电,而此叶绿素a则带正电,形成电荷分离。接着原初电子受体将电子交给次级电子受体,叶绿素a则从原初电子供体得到电子。这样,反应中心的叶绿素a及原初电子受体都恢复到原来状态,又可进行下一次光化学反应。
同化力形成 同化力的两个组成部分NADPH和ATP分别由光合电子传递和光合磷酸化作用形成,而光合磷酸化又偶联于电子传递链,并利用电子传递中的部分能量。
光合电子传递 叶绿素 a的电子在光化学反应中交给初级电子受体后,又经过一系列电子载体传递,最终交给辅酶Ⅱ,使之还原;与此同时,原初电子供体在交出电子给叶绿素后,又从次级电子供体取得电子。这样依次传递,最终从水中取得电子,使水氧化,释放出氧气。这一系列电子传递过程称光合电子传递。英国人R.希尔于20世纪30年代末首先发现离体叶绿体在光下可使外加电子受体如高铁盐或铁氰化钾还原,并从水中释放出氧气。此反应称希尔反应。
对于光合电子传递中光化学反应与电子传递衔接的机理,现在较普遍被接受的看法是两个光化学反应与一系列电子载体串联起来,形成电子传递链。按电子载体的电位高低作图,得到“Z”形图解(见光合电子传递)。
这些电子载体在类囊体膜上分布于不同纵深部位,电子在它们之间定向传递。水在类囊体膜内侧交出电子,电子经一系列传递后在膜外侧传给质醌,同时质醌从间质中取得氢离子并与之结合,被还原的质醌转移到膜内交出电子并释放出氢离子。电子经一系列传递最终在膜外侧交给辅酶Ⅱ。这样,定向电子传递除从水中释放出氧气和使辅酶Ⅱ还原外,还同时形成膜内外的电位差(膜内正,膜外负)及氢离子浓度差(膜内浓度高于膜外)。
光合磷酸化 叶绿体(或载色体)在光下将无机磷酸和腺苷二磷酸(ADP)转化为ATP的过程。50年代D.I.阿尔农发现离体叶绿体在光下可进行两种类型的光合磷酸化反应,一种与有氧的释放和NAD+的还原的非循环电子传递链相偶联,称非循环光合磷酸化,可以下式表示:
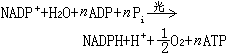 式中Pi为磷酸盐,n的数值尚无定论,最初认为是1,以后有人认为可以是 1.33或2。另一种是与循环电子传递偶联的 ATP形成不伴有氧的释放和还原态电子受体的积累,称循环光合磷酸化,可以下式表示:
式中Pi为磷酸盐,n的数值尚无定论,最初认为是1,以后有人认为可以是 1.33或2。另一种是与循环电子传递偶联的 ATP形成不伴有氧的释放和还原态电子受体的积累,称循环光合磷酸化,可以下式表示:
光合磷酸化与光合电子传递的偶联机理,P.D.米切尔的化学渗透假说认为:光合电子传递所产生的膜内外电位差和氢离子(又称质子)浓度差〔两者合称质子动子势(pmf)〕,即是光合磷酸化的动力。氢离子通过突出在类囊体膜上的蛋白复合体──偶联因子将 ADP和Pi转变成ATP。一般认为,偶联因子对ATP形成的催化与其构象变化有关。
碳同化 同化力(NADPH及ATP中所含的化学能)在一般情况下主要用于推动光合碳循环、同化 CO2形成蔗糖等碳水化合物或其他有机物,但也可有一部分用于同化硫和氢等无机化合物。光合碳循环又称卡尔文循环,此循环的大部分反应均在叶绿体的间质中进行,但从磷酸丙糖转化成蔗糖的一些步骤则是磷酸三糖通过叶绿体被膜转移到细胞质中后,在细胞质中完成的(见光合碳循环)。
植物的光合碳循环常伴随着光呼吸。有些植物中,在CO2由光合碳循环同化前,先通过四碳途径或景天科酸代谢途径固定在四碳双羧酸中。这些都是和碳同化密切关联着的反应。
光呼吸 植物在空气中进行光合作用时,碳同化的中间产物中有一部分被氧化成CO2,其过程和一般有机物氧化成CO2的呼吸作用不同。它是由于O2和CO2竞争,与二磷酸核酮糖反应生成磷酸乙醇酸而引起的。磷酸乙醇酸通过乙醇酸途径氧化而放出CO2。一般植物中,光呼吸所放出的 CO2量常可达当时光合作用同化CO2量的1/3左右。降低空气中氧气浓度,光呼吸就会减少。
四碳途径和景天科酸代谢 有些起源于热带的植物如玉米、甘蔗等进行光合作用时,CO2先在叶肉细胞内与磷酸烯醇式丙酮酸反应,被固定在四碳双羧酸中,后者转运到维管束鞘细胞中再释放出 CO2供光合碳循环同化。四碳双羧酸的形成和运输起CO2泵的作用,在鞘细胞中造成一个高于空气中CO2浓度的微环境,以利于光合碳循环的进行。这些植物的光合速率较高,光呼吸较低。此外,一些肉质植物(如景天、仙人掌、菠萝等)中存在景天科酸代谢,其反应与四碳途径相仿,但它是在夜间将CO2固定在四碳双羧酸中,白天释放出来供光合碳循环同化,起一个 CO2库的作用。因此使这些植物在白天因缺水而气孔关闭的情况下也能进行光合作用。有四碳途径的植物常被称为四碳植物,无四碳途径或景天科酸代谢的植物常被称为三碳植物。
光合作用的效率及其影响因素
在优良条件下测定光合作用效率的结果和对光合作用机理的分析表明,植物在光合作用中每同化一个CO2分子或碳水化合物需要 8~10个光量子。按此量子需要量推算,植物利用太阳总辐射的效率可达10%左右。实际上,大田中作物的光能利用率远低于此数。一般作物生物生长良好的田块,一季中光能利用率只有1~2%,其原因和作物的生理特性、环境条件和群体的结构有关。
光合作用和植物生理状态的关系 光合作用的机构和植物其他机构一样,有它的发生、成长和衰老过程。在发育初期光合速率较低;成长后较高,且稳定一段时期;到衰老时下降。植物的生理状态和外界条件对光合速率各阶段的持续时间和光合速率高低有显著影响。光合机构的光合产物大部分输出到植株其他部分,供各种生命活动之需。当植株其他部分对光合产物需要量增加时,光合速率常会提高。反之,当光合产物利用缓慢,输出受阻时,光合速率会逐渐下降。此外,植株的其他部分还能通过激素传递等方式影响光合机构的发育和运转。
光合作用和环境条件的关系 许多环境条件的变动可影响光合机构的运转,植物的光合机构对它的环境也有一定的适应能力。
光 是进行光合作用所需的能源。光照较弱时,光反应速率常是光合作用速率的限制因素。光合速率和光强变化成正比关系。光强较高时,光合作用受其他反应(统称暗反应)速率的限制,所以其速率不能随光强成正比地提高,并逐渐达到饱和。光强太高时,光合机构还会受到破坏。
不同类型的植物在不同光照强度下生长时,其光合速率达到饱和的光强有显著的差别。一般阳生植物(包括绝大部分农作物)的饱和光强接近晴天中午日光(约1kW/m2)的一半。生长良好的时候,其光合速率一般为10~20μmol m-2s-1(1μmol m-2s-1=1.58mgCO2dm-2h-1)。玉米、甘蔗等四碳植物的光合作用在中午日光下也未达到饱和,其速率可达25~35μmolm-2s-1。阴生植物如酢浆草等的光合作用在光强不到中午日光的1/10时便会饱和,光合速率小于6μmolm-2s-1。但其光补偿点(光合速率与呼吸速率相等时的光强)也很低。太阳辐射中植物光合作用所利用的波长范围为380~720纳米,有的细菌的光合作用的利用范围可延伸到1000纳米。不同光质的光对光合机构的发育及光合产物的种类有不同的影响。
温度 光合作用的暗反应受温度影响。低温下光合速率较低,温度上升10℃,光合速率可提高一倍左右,至最适温度以上再增加温度则光合速率下降。呌植物的光合作用最适温度一般为 20~30℃,C4植物一般为 35~40℃。温度过高或过低都会破坏光合机构。植物光合机构对所处温度条件有一定适应能力,不同种类植物的适应范围有显著差别。
水 作为光合作用的原料,每形成1克有机物约需0.6克水,这和陆生植物光合作用时所消耗水的数量相比是微不足道的,大量水分是蒸腾掉的。光合机构通过叶片等表皮上张开的气孔从空气中吸收CO2时,会大量蒸腾散失水分。一般呌植物蒸腾掉的水分重量和光合作用积累的有机物的重量的比值称蒸腾系数,常在300左右,C4植物因为有CO2泵,可低至200以下,景天科酸代谢的植物可小于100。植物缺水时,气孔关闭,光合作用受阻;进一步缺水则光合机构损坏。
气 空气中的CO2是光合作用的原料,它的浓度较低,一般为340vpm〔1vpm=1/1000000(体积比)〕,且在它到达同化部位的通路上,要经历周围大气─→叶片表皮
 叶肉细胞表面─→叶绿体内这 3大阶段的阻力(呌植物中,它们的比例大致为1:2~6:8)。所以CO2的供应常是植物在自然条件下光合速率的限制因素(C4植物因有CO2泵,在CO2浓度为0.03%时已接近饱和)。气流通过叶面的速度在一定范围内(一般小于2m/s)和光合速率成正相关。空气中CO2含量增至0.1%左右时,可显著提高光合速率。温室中CO2不易散失,可以增施CO2以提高产量。
叶肉细胞表面─→叶绿体内这 3大阶段的阻力(呌植物中,它们的比例大致为1:2~6:8)。所以CO2的供应常是植物在自然条件下光合速率的限制因素(C4植物因有CO2泵,在CO2浓度为0.03%时已接近饱和)。气流通过叶面的速度在一定范围内(一般小于2m/s)和光合速率成正相关。空气中CO2含量增至0.1%左右时,可显著提高光合速率。温室中CO2不易散失,可以增施CO2以提高产量。空气中所含浓度为21%的氧气是引起光呼吸的重要因素。空气中常含有 SO2、O3等污染物质,虽含量甚微,但超过限度就会引起气孔关闭,甚至损害光合机构。
肥 植物从土壤中吸收的无机元素在植株组成中所占的比重虽然比光合作用形成的有机物小,但其中许多种为光合所必需。例如,参与光合机构组成或运行的元素有 N,P,K,S,Mg,Fe,Mn,Cu,Zn,Cl等。其中N,P,Mg等还起着多重作用。当这些无机营养元素缺乏或过多时,光合机构的形成和运行就会受到影响。缺绿是多数元素缺乏时常出现的病征。
群体光能利用
在农田、森林、草地及各种自然植被中,常常不是单个的植物进行光合作用,而是由许多植株组成群体利用光能。群体内部形成了特殊的微环境,其中的光能分布、气体流通等都与外界显著不同。决定单位地面上太阳光的捕获量的,也不是单个叶片的大小,而是叶片总面积的多少。群体叶面积的多少以单位面积土地上叶面积的单位数来表示,称为叶面积指数(或系数)。它无论以亩(亩/亩)或平方米(m2/m2)计算时,都是无量纲数。当叶面积指数小于1时,对太阳光能的捕获和利用不充分。叶面积指数大于1时,虽然有一部分叶片被其他叶片遮蔽,所受光强低于自然光,但由于叶片交错排列,下层叶并不处于完全黑暗之中,仍能进行一定程度的光合作用。叶层过多,下部叶片受光极弱,可能低于补偿点,它们不能对群体干物质累积有所贡献。对每种特定条件,群体叶面积指数有一个最适值,多数情况下在3~5之间。
天空光特别是太阳直射光照射到叶片上时的入射角大小,影响受光的强弱。入射角小(光线与叶片接近垂直)则光强高。由于光饱和现象,光强高则光能利用率低。中午前后太阳光入射角较小,如叶面积指数足够大时,直立叶群体光能利用率高于水平叶,早晚则相反。禾本科的稻、麦等叶片常接近直立,而双子叶植物中的向日葵、棉花叶片接近水平。
植物的叶片等光合器官本身,也是用光合产物形成的。光合产物用于形成叶片的比例大,叶片薄,都有利于捕获光能和CO2。但叶片以外的其他部分,对叶片等光合器官起支持或支援作用,如根吸收水和无机养料,茎支撑叶片并输送养料等,也不宜过分削弱,因而叶片所占比例也有最适范围。
在农业生产上,常用高密度、高肥(特别是氮肥)或灌水来促进叶面积的形成和发展;用间作套种来提高地面覆盖率。温带早春气温低,是限制叶面积形成及光合作用进行的因素。除覆盖栽培(或称设施农业)以外,只能用选择能耐受低温的植物种(如冬小麦)或品种来适应。前后作茬口搭配得当,栽培条件适于维持旺盛的光合作用,可取得较多的总光合产物量(生物量)。
农作物除叶菜类和饲料作物外,收获物主要的常是非光合器官,如果实、种子、块根、块茎等。因而农业生产上既要求形成足够大的叶面积指数,并维持较长的有效期,也要求及时并大量地形成经济价值较大的构成产量的器官。所以生产上运筹栽培措施和茬口搭配时,需要将光合器官的数量和工作效率与光合产物在各器官间的分配结合起来考虑,求得最大的经济效益。
参考书目
殷宏章、沈允钢:光合作用研究进展,《自然杂志》1979/2。
沈允钢、王天铎:《光合作用──从机理到农业》,上海科学技术出版社,1978。
D.O.Hall and K.K.Rao, Photosynthesis, 4th ed., Edward Arnold Ltd.,London,1987.