代谢(植物)(卷名:生物学)
metabolism(plant)
植物利用太阳能和无机物质,形成体内的有机物,并用于各种生命活动,同时排除废物和多余的能量的过程。代谢为一切生物所共有,植物独特之处,在于自养性,即利用太阳光能,通过光合作用从二氧化碳、无机盐和水合成各种含能有机物质,供其生命活动之需。植物的非光合器官,或在黑暗中的光合器官的生命活动,也靠利用现成的有机物,即叶片白天形成的光合产物及其中的能量,因而具有与其他生物相似的呼吸代谢及承担呼吸的细胞器──线粒体。
植物的代谢产物,除二氧化碳和水向体外排出之外,其他废物一般不排出体外,而是累积于液泡之中。液泡由液泡膜与原生质隔开,因而其中的有毒物质和不适宜的pH值不致伤害原生质。某些植物形成并积累多种特殊产物,称为次生物质,其生理功能多不明。
植物的代谢是按照遗传基因的潜势,在环境条件的制约下有序地进行的,随着发育的进程而形成不同的器官,合成不同的物质。遗传基因的负载者──脱氧核糖核酸(DNA)和核糖核酸(RNA),以及响应外界条件、调节体内生理活动的植物激素,和催化各种生物化学反应的酶,既是代谢的产物,又反过来影响代谢。因此植物代谢的内容极其广泛。
光合、呼吸与代谢 光合作用是绿色植物有机物质与能量的源泉。非光合器官以及黑暗中的光合器官都通过呼吸作用来利用光合产物中贮存的能量,并利用呼吸作用的中间产物形成各种复杂的有机化合物。图1 显示了植物的代谢的主要途径和光合与呼吸在其中的地位。在生物化学的历史上,曾经把从葡萄糖、果糖或其磷酸酯开始,经糖酵解和三羧酸(TCA)循环释放CO2的整个过程看作呼吸作用,从而认为脂肪、氨基酸等类化合物都是从呼吸作用的中间产物衍生出来的。严格意义的呼吸则只包括二氧化碳的释放(脱羧反应)和氧的吸收(末端氧化反应),而将导向二氧化碳释放的反应看作降解代谢,其中的一些中间产物可转而走向合成代谢,形成各种更复杂的、往往还原程度更高的化合物。合成代谢中多处需要呼吸作用产生的腺苷三磷酸 (ATP)供能,因而整个代谢都与呼吸作用有密切的联系。
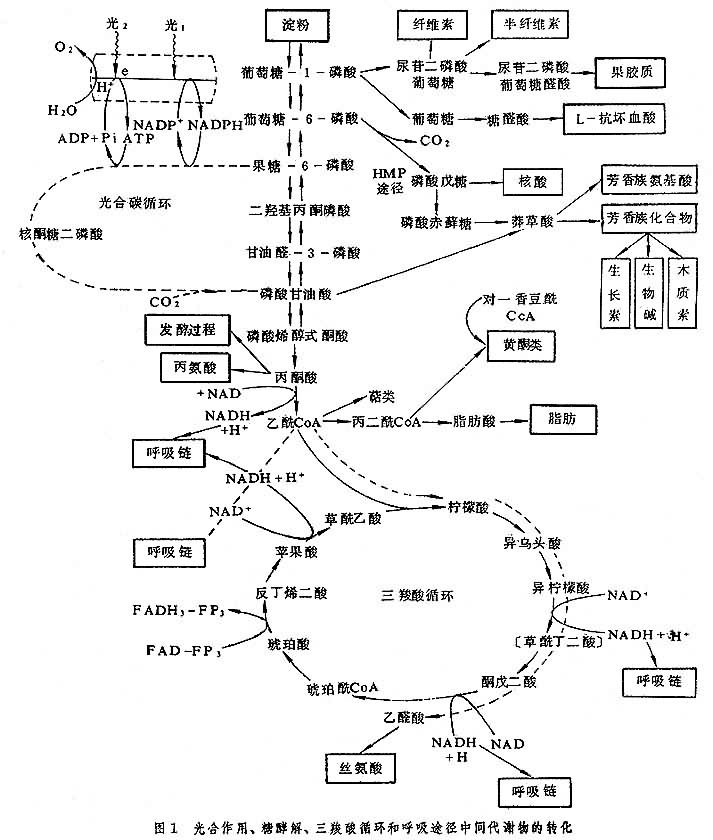
除以光合作用产物己糖磷酸和蔗糖的形式经呼吸作用为代谢提供有机物和能量以外,代谢和呼吸还有其他联系和方式。例如:①光合系统提供的ATP,NADPH和还原的铁氧还素可以推动绿色细胞中需要高能磷酸键或还原力的反应;②光合磷酸化提高能荷值后会抑制呼吸作用;③光合碳循环和光呼吸的中间产物经过转氨作用可以形成氨基酸,如天门冬氨酸、丙氨酸、丝氨酸、甘氨酸。
代谢的多条途径 植物代谢的各个部分,往往不止一条途径。例如碳水化合物降解的途径有糖酵解 (EMP)途径和磷酸戊糖(HMP)途径;呼吸链的电子传递有5条不同途径,并且可以通过多种末端氧化酶把电子传递给氧气。
不同的代谢途径同时存在于植物体内,但各途径运行的速率并不相等,速率间的比例也不固定。在特定环境中,植物的代谢以哪一途径运行,以及途径之间的变化,依植物的种类和器官、生长发育的状况及环境条件而定。例如:①地下组织(根系)的EMP途径-TCA循环比重较大,而地上组织 (茎、叶)则HMP途径占优势。②年幼的、生长旺盛的组织中,EMP途径-TCA循环占主要地位,而老的组织则以 HMP途径为主。这些不同途径的中间物是植物体内不同的物质的代谢基础,并且常与相应的生理过程密切相关。
植物代谢的多样性是植物在进化过程中对多变的环境的适应。
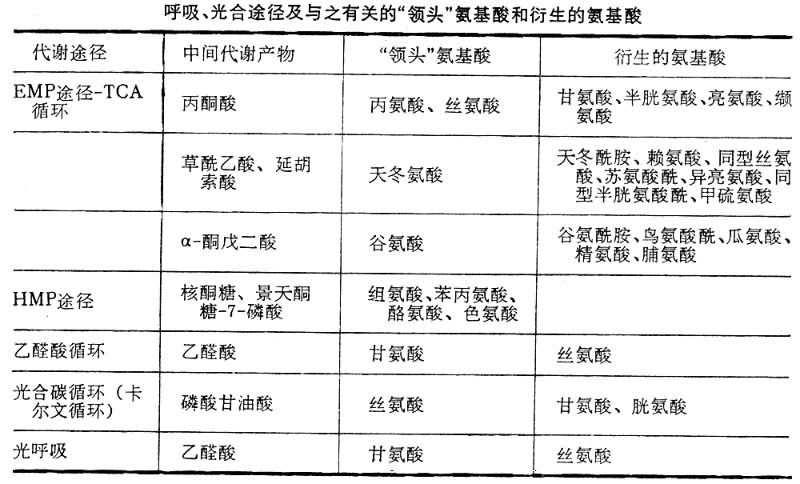
各类物质的代谢 氨基酸形成 其主要途径是呼吸代谢的中间产物有机酸通过加氨作用,形成“领头”氨基酸,后者在转氨酶催化下,通过转氨作用以及其他转化作用形成多种多样的氨基酸。不同的呼吸途径的中间代谢产物不同,形成的“领头”氨基酸和与之相适应的氨基酸代谢也不同(见表)。
脂肪代谢 脂肪的降解和合成的途径不同。脂肪的降解产物是甘油和脂肪酸。甘油经磷酸化形成α-磷酸甘油, 然后脱氢形成磷酸丙糖。后者既可循 EMP途径逆方向经1,6-二磷酸果糖转成蔗糖,也可经丙酮酸进入TCA环彻底氧化。脂肪酸则以β-氧化方式反复形成乙酰COA,后者经乙醛酸循环转变为琥珀酸,琥珀酸然后转移到线粒体上经TCA环彻底氧化;或者经草酰乙酸、磷酸烯醇式丙酮酸(PEP)循EMP途径逆方向转成蔗糖。
油料植物在种子形成过程中积累大量脂肪,这些脂肪是从糖转化而来的。脂肪的积累和 HMP途径的加强平行。
激素的形成 生长素 (吲哚乙酸,IAA)生物合成的前体是色氨酸。而色氨酸是通过HMP途径形成的,体内其他生长素类物质如反-肉桂酸和对-香豆酸也是通过 HMP途径的中间代谢形成莽草酸后进一步转化成的。
乙烯生物合成的前体是蛋氨酸,后者是由TCA环中草酰乙酸形成的天冬氨酸转化而成的。
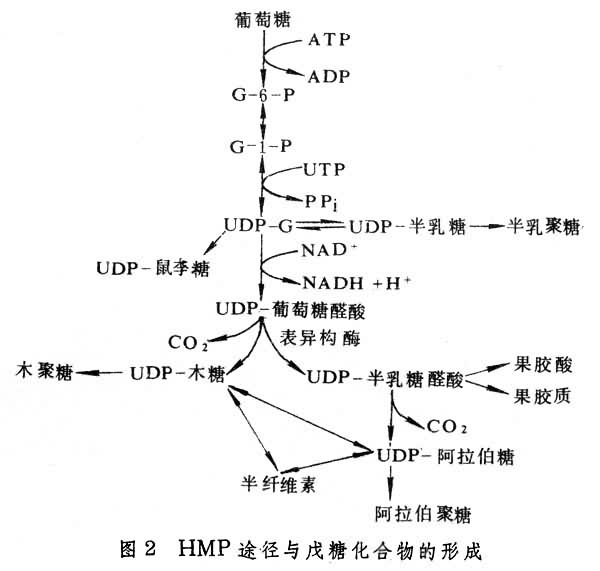
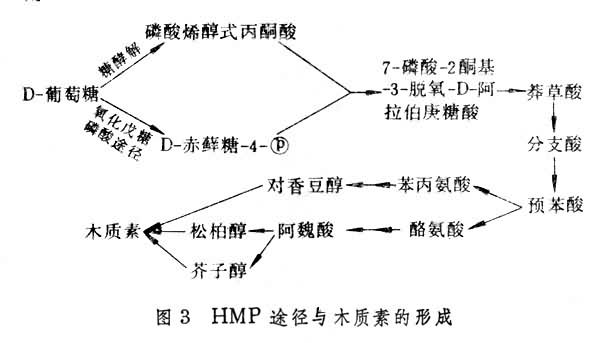
细胞壁结构物质的形成 细胞壁结构物质有纤维素、半纤维素、果胶质和木质素等, 而果胶质类中半乳糖醛-鼠李聚糖和阿拉伯-半乳聚糖是细胞壁的重要组分。细胞壁结构物质中除纤维素外,大部分物质的形成可能与HMP途径有关。其联系途径主要有两个:①通过HMP途径形成多种戊糖化合物(图2);②HMP途径的中间代谢通过莽草酸途径形成苯丙氨酸和酪氨酸,后者分别进一步转化成对-香豆醇及松柏醇和芥子醇,这些都是合成木质素的前体(图3)。
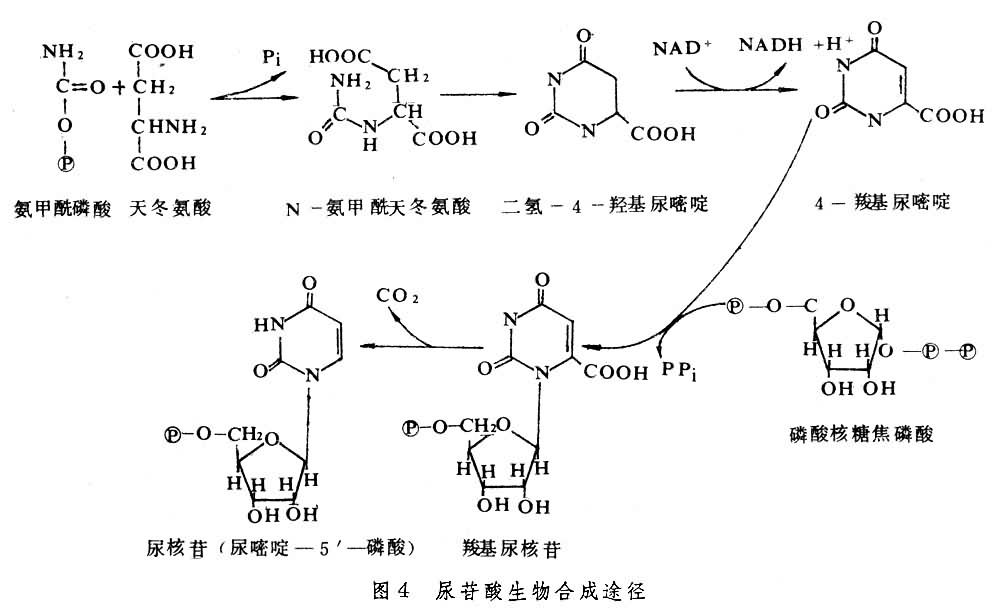
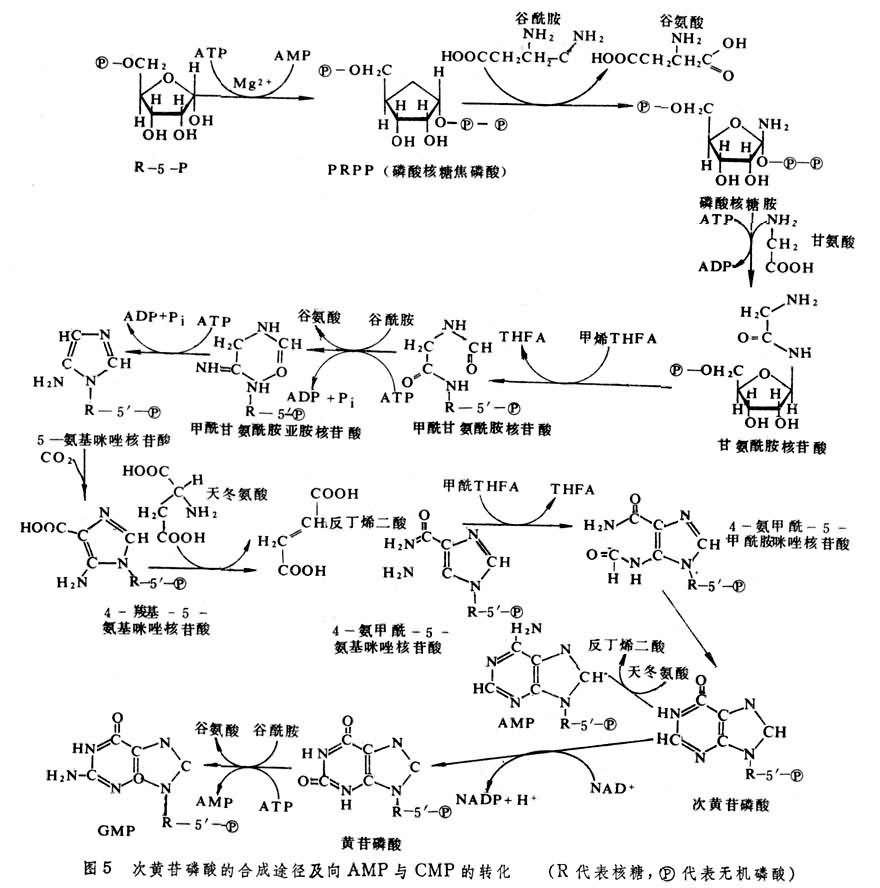
核酸类物质的形成 RNA与DNA在遗传性的传递与表达中起关键作用。 ATP是生物体内能量传递的通货。核酸类物质都由嘧啶碱或嘌呤碱、戊糖(核糖或脱氧核糖)和磷酸等 3部分组成。其中戊糖来自氧化戊糖磷酸途径。嘧啶碱与嘌呤碱的生物合成方式大不相同。嘧啶碱的合成过程是先合成嘧啶环,以后再与核苷酸结合,如图4所示(以尿苷酸为例)。嘌呤碱的合成过程则是从核糖核酸开始,先与谷酰胺反应,形成磷酸核糖胺,以后分步先后形成咪唑环和次黄 (嘌呤)苷磷酸(图5),后者经转化形成鸟核苷磷酸或腺核苷磷酸。
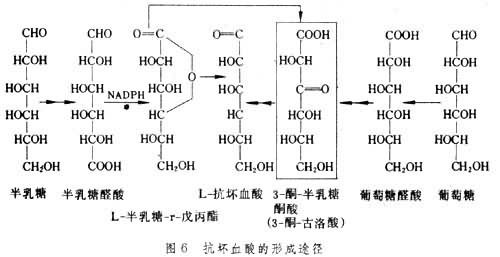
抗坏血酸形成 抗坏血酸是通过葡萄糖醛酸或半乳糖醛酸进一步转化而形成的(图6)。
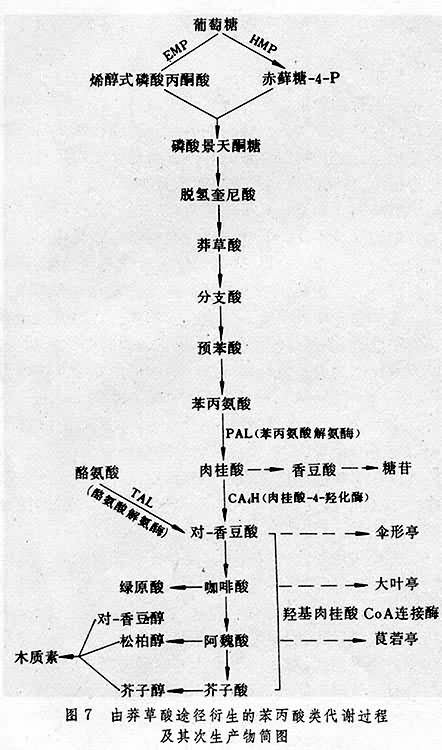
次生物质的形成 次生物质类别繁多,包括萜类、类黄酮、糖苷、酚类、生物碱、木质素等,在植物体内广泛存在。不同类化合物分别由降解代谢中的不同中间代谢物转化形成。例如由 EMP途径-TCA循环的中间代谢物乙酰 CoA通过甲羟戊酸产生次生代谢产物异戊二烯类(萜类),由EMP,HMP的中间代谢形成磷酸景天酮糖,通过莽草酸途径进一步进行苯丙酸类代谢,产生多种次生代谢产物(图7)。
能量代谢 植物首先通过光合作用把太阳的辐射能转变为化学能贮藏于有机化合物中。然后一部分通过呼吸代谢以及其他多酶系统把各种有机化合物中的化学能分解出来而加以利用。其中一部分用于作功,如生物合成、根瘤固氮、维持跨生物膜的浓度差、离子吸收和体内的逆浓度梯度输送。另一部分以热的形式放出,在某些情况下,可显著提高体温。
植物体内各种需能反应都以 ATP为直接能源。产生ATP的途径有两条,即光合磷酸化作用和氧化磷酸化作用,前者为植物所特有,后者则为动植物所共有。碳水化合物、脂肪和蛋白质等有机物在氧化时都可产生ATP,其机理也有两种。一种与底物的氧化步骤相联系,称为底物水平磷酸化;另一种与呼吸作用的电子传递(也称呼吸链)相联系,称为电子传递链水平磷酸化。后者是形成ATP的主要途径。不同电子传递途径中产生的ATP分子数不都相等,而其P/O比(即每吸收一个原子O所形成的ATP分子数)也不等。例如电子传递主路P/O=3,而抗氰电子支路P/O=1。 ATP产生的量因植物种类、生长发育时期和环境条件不同而异,ATP的数量反过来又可影响和调节代谢和生理过程。
代谢与种子萌发 种子萌发时,代谢活动非常旺盛。贮藏在胚乳或子叶中的有机物分解,成为合成新器官所需的蛋白质、核酸、纤维素等物质的原料;同时有一部分氧化,形成ATP,以满足物质运输和有机物合成时的能量需要。一般种子在短期吸胀后,ATP即成倍增加。如萝卜种子吸胀1小时后,每粒种子所含的ATP即从0.1纳摩尔增加到2.5纳摩尔,同时种子的胚根和鲜重增加。
种子萌发过程中,代谢途径也发生变化。如水稻、大麦种子发芽时,呼吸途径从早期的EMP途径-TCA循环转向HMP途径。
代谢与冷害 喜温植物受低温冷害时,呼吸强度先明显提高,然后下降。离体线粒体呼吸活性和氧化酶活动也表现类似变化。
低温还会引起代谢途径的改变。 例如马铃薯在低温(1.7℃)下贮藏时,HMP 途径的酶系活性增强,呼吸对碘代乙酸不敏感而对SHAM敏感,电子传递从细胞色素途径转向其他的途径,主要是抗氰途径。
零上低温对植物的影响与原生质膜变化有关。首先是生物膜脂质发生相变,膜透性增大,离子外渗,破坏了细胞内的离子平衡;同时膜的结合酶活力下降,破坏了与游离酶之间反应速率的平衡,从而使呼吸受阻,能量供应减少,体内积累有毒物质,最后导致植物死亡。
代谢与感病性 宿主植物在感染病菌或病毒后,呼吸强度明显升高,主要是含铜的氧化酶抗坏血酸氧化酶或多酚氧化酶的活动增强。同时 HMP途径有不同程度的增强。例如红花下胚轴感染锈病后呼吸对 NaF的敏感度下降,丙二酸的抑制也明显减弱,表明感病后EMP途径-TCA循环降低,而HMP途径明显增强,HMP途径的酶系G-6-P脱氢酶和6-PG脱氢酶活性也都增强。
感病后的植物组织中催化苯丙氨酸解氨酶 (PAL)活性明显增强,而苯丙氨酸类的代谢产物木质素起防止病原菌侵入的保护圈作用。苯丙氨酸解氨酶增强的程度与品种的抗病能力平行,因此在某些情况下该酶的活性可用作抗病的生理指标。
植保素是一类抗病物质,包括类黄酮、酚类(如绿原酸)、香豆素类、生物碱等,它们主要是PAL代谢的产物。很多植物感病后短时间内就产生植保素,它的产生和积累与PAL活性的增强平行。
调节与控制 调节代谢中的反应步骤的途径有两类,一类是改变反应底物和产物的浓度,使反应依照质量定律而加速、减缓或改变方向;另一类是改变催化各反应的酶的数量或活力。
酶活力的调节又可分为对酶蛋白形成的调节和对酶比活性的调节。前者是通过影响酶蛋白合成的速率,也就是影响基因表达中从转录到翻译的一段过程。其中有些酶在条件有利于反应时形成,被称为适应酶。例如在强光下叶片中RuBP羧化酶的数量增加3~4倍,这有利于利用强光。有些酶以不同的蛋白质分子形式存在,这些分子结构不同而功能相同的酶称为同功酶。在高温下最适温度较高的同功酶优先形成。另一些酶在有底物时形成,称为诱导酶。突出的例子是硝酸还原酶。它只有在植物体内有
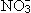 存在时才形成。通过增减酶的合成量或改变不同同工酶的相对合成量来调节,有利于对蛋白质的经济利用,但所需的时间较长,可称为粗调。对酶的比活性的调节则不改变酶蛋白的数量。这种调节比较迅速,也容易逆转,可称为细调。
存在时才形成。通过增减酶的合成量或改变不同同工酶的相对合成量来调节,有利于对蛋白质的经济利用,但所需的时间较长,可称为粗调。对酶的比活性的调节则不改变酶蛋白的数量。这种调节比较迅速,也容易逆转,可称为细调。酶活力的调节机理大致可以归纳成为以下几种:①变构效应:许多酶具有变构特性,当与效应剂结合时,空间构象发生变化,活性也因而改变。无机离子如Mg2+,Mn2+,K+,
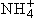 ,一些酶的辅因子和小分子代谢物都可以起效应剂作用。很多代谢途径的最终产物也以变构效应影响酶的活性,这是一种反馈调节,有正反馈即激活作用和负反馈即抑制作用两种。如柠檬酸对 F-6-P激酶的抑制就是负反馈。②改变酶分子聚合程度:有些酶分子可以以单体、二聚体、四聚体等多种形式存在,它们之间活力差异很大。有些几乎没有活力。当需要某一酶的活力时,这种酶分子就从无活力或低活力的聚合状态转向有活力或高活力的聚合状态。③能荷调节:能荷代表细胞腺苷酸库中高能磷酸键的相对饱和度,其数值高低影响多种酶(特别是各种激酶)的活性。 能荷值的增加(代表ATP积累)往往提高利用ATP途径的酶而抑制参与ATP再生途径的酶的活性,从而防止形成过多的ATP。前一类包括豌豆和菠菜中的磷酸甘油酸激酶和菠菜的核糖磷酸激酶,后一类包括 F-6-P激酶、磷酸甘油醛脱氢酶、柠檬酸合成酶和异柠檬酸脱氢酶。与此类似,氧化态辅酶I(NAD+)在辅酶I总量中的比例
,一些酶的辅因子和小分子代谢物都可以起效应剂作用。很多代谢途径的最终产物也以变构效应影响酶的活性,这是一种反馈调节,有正反馈即激活作用和负反馈即抑制作用两种。如柠檬酸对 F-6-P激酶的抑制就是负反馈。②改变酶分子聚合程度:有些酶分子可以以单体、二聚体、四聚体等多种形式存在,它们之间活力差异很大。有些几乎没有活力。当需要某一酶的活力时,这种酶分子就从无活力或低活力的聚合状态转向有活力或高活力的聚合状态。③能荷调节:能荷代表细胞腺苷酸库中高能磷酸键的相对饱和度,其数值高低影响多种酶(特别是各种激酶)的活性。 能荷值的增加(代表ATP积累)往往提高利用ATP途径的酶而抑制参与ATP再生途径的酶的活性,从而防止形成过多的ATP。前一类包括豌豆和菠菜中的磷酸甘油酸激酶和菠菜的核糖磷酸激酶,后一类包括 F-6-P激酶、磷酸甘油醛脱氢酶、柠檬酸合成酶和异柠檬酸脱氢酶。与此类似,氧化态辅酶I(NAD+)在辅酶I总量中的比例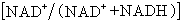 的变化影响各种脱氢酶的活性,如比例升高时抑制苹果酸脱氢酶的活性,降低时抑制异柠檬酸脱氢酶的活性。④酶的氧化或还原:这是对酶的共价键修饰。氧化磷酸戊糖途径中的 G-6-P脱氧酶、糖酵解途径中的磷酸果糖激酶等在被氧化或还原时酶活性都发生变化。在体内光可通过铁氧还蛋白/硫氧还蛋白系统调节它们的氧化还原状态。⑤酶的磷酸化或去磷酸化:这也是一种共价键修饰。蛋白激酶使酶磷酸化,而磷酸酯酶使酶去磷酸化。如豌豆叶线粒体的丙酮酸脱氢酶复合体经磷酸化或去磷酸化时,活性发生变化。⑥pH的调节:酶的活力依赖于pH值。植物体可通过改变pH来调节酶的活力。
的变化影响各种脱氢酶的活性,如比例升高时抑制苹果酸脱氢酶的活性,降低时抑制异柠檬酸脱氢酶的活性。④酶的氧化或还原:这是对酶的共价键修饰。氧化磷酸戊糖途径中的 G-6-P脱氧酶、糖酵解途径中的磷酸果糖激酶等在被氧化或还原时酶活性都发生变化。在体内光可通过铁氧还蛋白/硫氧还蛋白系统调节它们的氧化还原状态。⑤酶的磷酸化或去磷酸化:这也是一种共价键修饰。蛋白激酶使酶磷酸化,而磷酸酯酶使酶去磷酸化。如豌豆叶线粒体的丙酮酸脱氢酶复合体经磷酸化或去磷酸化时,活性发生变化。⑥pH的调节:酶的活力依赖于pH值。植物体可通过改变pH来调节酶的活力。 有些化合物(例如ATP和NAD+),既是某些酶反应的底物(或产物),又是另一些酶的效应剂,因而它们调节代谢的作用点不止一个,调节方式也不止一种。
除了对酶的数量和活性的调节以外,植物还可以以其他方式影响底物或产物的浓度,从而调节代谢中个别酶反应或整条代谢途径的速度。其调节方式有:①改变物质的运输速度或方向:对短距离的跨膜运输,可以通过对推动逆浓度梯度运输的呼吸的调节来改变各区隔中代谢物的浓度。对长距离运输,可通过对代谢物的源处的装入和壑处的卸出的影响来改变源壑关系;②激素的作用:植物激素能调节有机物质的运输,从而改变源壑关系;也能通过对生长、发育、离层形成等过程的影响而改变源壑关系;③气孔运动:气孔开闭影响二氧化碳的扩散,从而调节叶肉细胞二氧化碳的供应,影响光合作用的速率。
对植物代谢的认识有助于阐明各种生理过程的本质和机理。这对于提高植物的物质生产,特别是增加某些特殊物质包括多种次生物质的合成,和改进农林产品的品质,都极为重要。