主要组织相容性复合体(卷名:现代医学)
major histocompatibility complex,MHC
能诱导强排斥反应并参与免疫调节的基因组。
20世纪初,E.E.泰泽发现近交系小鼠的肿瘤移植到同一近交系小鼠体内后能够生长,而移植到另一近交系小鼠体内则遭排斥。随后P.B.梅达沃发现正常组织移植也出现类似的现象,认为移植排斥是一种免疫反应。进一步研究表明,在遗传性不同的个体间进行组织移植,细胞表面的同种异型抗原诱导了针对移植物的免疫反应。G.D.斯内尔(1948)将决定移植物是否被排斥的抗原称为组织相容性抗原(H抗原),控制它表达的基因称为组织相容性基因(H基因)。他随后发现有一个H基因能诱导强移植排斥反应,称之为主要组织相容性基因,后来发现它是由多个基因组成,并且位于同一染色体片段上,因此改称之为主要组织相容性复合体,而将那些诱导弱移植排斥反应的H基因称为非MHC基因或次要组织相容性基因。
1958年J.多塞发现多次输血者和多产妇血清中含有凝集人白细胞的抗体,其特异性不同于红细胞凝集抗体。随后的研究发现人类白细胞抗原 (HLA)是一个极为复杂的遗传系统的产物。这一遗传系统称为人类白细胞抗原复合体,就是人的主要组织相容性复合体。
60年代报道了对白血病病毒的抵抗力依赖于小鼠的H-2复合体。随后又发现人的一定HLA抗原的存在和对某些疾病的易感性间存在着惊人的相关性。这导致人们对MHC生物效应的广泛研究,取得关于免疫应答基因(Ir基因)和免疫细胞间相互作用的MHC限制性等的重大进展。
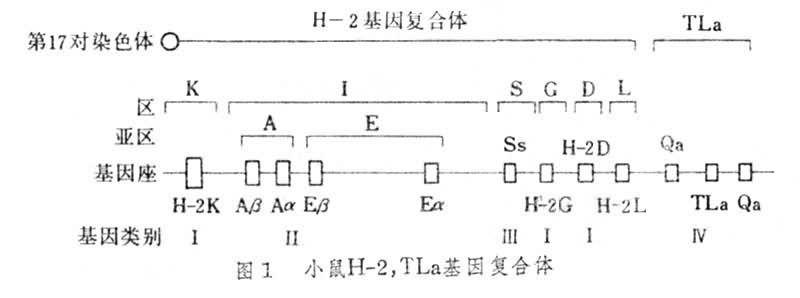
H-2基因复合体 小鼠主要组织相容性抗原受第17对常染色体 H-2基因群支配,可分为六个主要区,即K、I、S、G、D和L区。I区又可分为两个亚区,即I-A和I-E亚区。每个区或亚区至少包括一个基因座,分别称为H-2K座、H-2D 座、S座、H-2G座。I-A亚区含有Aα、Aβ二个座,I-E亚区含有Eα 和Eβ二个座。它们的功能各不相同, K区与D区基因可编码 H-2抗原系,I区基因编码的分子称为I区相关抗原或Ia抗原系(Ia), S区基因可编码血清因子和补体成分,G区基因与血型抗原有关。1981年J.克莱因将MHC基因座按功能分为四类(图1)。Ⅰ类座包括K、D和L座,其编码的分子称为Ⅰ类分子(即K、D和L分子);Ⅱ类座,其编码的分子称为Ⅱ类分子(Ia分子);Ⅲ类座,其编码的分子称为Ⅲ类分子(血清因子和补体分子);Ⅳ类座,位于D座右侧,是否属于H-2复合体尚未确定,但与H-2连锁,它包括TLa座和 Qa座,其编码的分子称为Ⅳ类分子(TL分子和Qa分子)。
HLA 基因复合体 1958年法国J.B.G.J.多塞首先在多次受血者的血清中发现能凝集人白细胞的抗异型白细胞抗原的抗体,并鉴定了第一个人白细胞抗原,定名为Mac(这是三个受检者名称的第一个字母)。其后近10年的研究证明,人组织相容性抗原也受多基因座控制,该基因座称为人主要组织相容性基因复合体(HLA复合体)。HLA复合体定位于第6对染色体短臂上,由4个基因区(HLA-A、B、C及HLA-D区)组成(图2)。D区相当于小鼠的Ⅰ区,由3个亚区,即HLA-DP、DQ和DR组成。HLA复合体也分为三类基因,Ⅰ类基因包括HLA-A、B、C三个座;Ⅱ类基因包括HLA-DR、DQ、DP三个座,每个基因座都表现为复等位基因,它们可编码 HLA抗原分子;Ⅲ类基因可编码部分补体分子。新发现属Ⅰ类基因的 HLA-E座和属于Ⅱ类基因的HLA-DZ、DO、DX等新出现的基因座,其定位及功能尚待进一步研究。
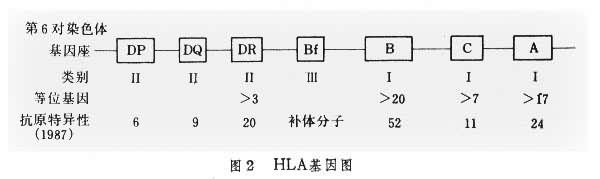
MHC的一个重要特征就是有高度的多态性,即在同一种属的群体中,同一个基因座可表达二种以上乃至几十种的基因产物。这是由于同一基因座存在着复等位基因。例如,1987年第10次国际组织相容性会议曾认为HLA-A座可表达24种抗原特异性,B座有52个,C座有11个,Dw有26个,DR座有20个,DQ座有9个,DP座有6个,共148个型别。这样可组成107种以上的单倍型,因此在无关人群中要找到HLA完全相同的个体,几乎是不可能的。HLA多态性为器官移植寻找合适的配型带来了很大困难,但在自然状态下,这可能是高等生物抵御病原体的一种适应性表现。
MHCⅠ类分子(HLA-A、B、C分子和H-2K/D分子)由两条肽链组成,一条为多态性糖蛋白,分子量为44000,由MHC编码称为重链(H)。另一条链为非多态性的β2-微球蛋白,分子量为12000,在人由第15对染色体编码,在小鼠由第2对染色体编码,称为轻链,以非共价键与H链结合,整个分子通过H链固定于细胞膜上。H链可分为三个区,自分子的氨基末端开始,依次是细胞外亲水区,穿膜的疏水区和胞内的亲水区。胞外区可分为三个功能区,每个功能区可包含约90个氨基酸残基,N-末端功能区具有寡糖侧链的附着部位,第二、三功能区各有一个二硫键环。抗原的多态性主要由这两个功能区的结构决定。胞内的亲水区能被磷酸化,这种反应可能与信号传递有关。
在小鼠的研究表明Ⅰ类分子见于除滋养层和绒毛膜外的组织,H-2K和H-2D抗原可存在于所有组织细胞表面,在小鼠胚胎第四天就可出现H-2抗原,在人胚胎组织也可发现HLA抗原。MHC的抗原密度依细胞种类而有显著不同,淋巴细胞密度最高,其次为肝、肾、骨骼肌和脑细胞。小鼠红细胞上有Ⅰ类抗原,而人红细胞上很难检测出Ⅰ类抗原。
MHCⅡ类分子(HLA-DR,DQ,DP分子及小鼠I-A、I-E分子)亦由两条大小不同的糖蛋白链组成,一条为多态性β链,分子量为33000,另一条为非多态性α 链,分子量为25000,亦为非共价结合,两条链都由MHC编码。每条链都由三个区组成,即胞外亲水区、穿膜疏水区和胞内亲水区。α 链胞外亲水区有两个功能区,都有一个寡糖附着部位,第二个功能区有一个链内二硫键,其胞内亲水区可被磷酸化。β链胞外亲水区也有两个功能区,都有链内二硫键。分析了抗原性不同的Ⅱ类分子的氨基酸序列,发现α 链都很相似,而β链则高度不同,这表明Ⅱ类抗原分子的多态性是由β链决定的。
Ⅱ类抗原分子的细胞分布与Ⅰ类抗原明显不同,后者可存在于多种组织细胞上,而前者只存在于少数种类的细胞,主要存在于B淋巴细胞,其次为巨噬细胞、血中单核细胞,上皮中的朗格汉斯氏细胞以及精子细胞等。不存在于静止状态的T细胞中,但可出现于活化T细胞上。
MHC的生理意义 MHC抗原最初是作为移植抗原而被发现的,是引起移植排斥的主要抗原系统。这种抗原不合,即可引起受体的免疫应答,排斥移植的供体组织。70年代后证明MHC分子还具有重要的免疫生理功能。
MHC分子在免疫应答过程中参与抗原识别。70年代R.M.津克纳泽尔等在小鼠实验中发现杀伤 T细胞在杀伤感染病毒的靶细胞时,只能杀伤同系感染靶细胞,而对不同系的感染靶细胞则无杀伤作用,称这种现象为遗传限制性。随后证明杀伤T细胞与靶细胞的MHC必需一致才有杀伤作用,因此又称此现象为MHC限制性。这揭示了MHC在T细胞识别异种抗原中的作用。进一步的研究证明T4+的T细胞在识别异种抗原时受MHCⅡ类分子限制,而T8+的T细胞识别异种抗原时受MHCⅠ类分子限制。这种限制性的机制是:T细胞通过其抗原识别受体,可同时识别异种抗原决定簇和自己MHC分子形成的新的复合抗原决定簇。
人们还发现外周血B细胞和单核细胞等非T细胞在体外能诱导某些自身反应性T细胞发生增殖反应,称这种现象为自身混合淋巴细胞反应(AMLR),并证明这是由非T细胞上MHCⅡ类抗原引起的。这种自身反应性T细胞在体内可能具有增强或抑制免疫功能的作用,借以维持机体的免疫稳定性,因此MHC分子也参予免疫调节作用。
研究证明,MHC分子对T细胞在胸腺内的分化成熟过程也起重要作用。体外研究发现:去除胸腺中MHCⅡ类抗原阳性的基质细胞,则T4+T细胞的发育受阻,在胸腺培养细胞中加入抗MHCⅡ类抗原的单克隆抗体,也能阻止T4+T细胞的发育。目前认为MHC分子在T细胞自身耐受的形成和T细胞库的产生中都起着重要作用。
MHC与疾病 研究发现许多疾病与某些HLA等位基因或HLA单倍型确实呈现明显的相关性。与HLA抗原相关的疾病有几个应当注意的特点:病因和病理生理机理未明,以遗传模式分布但为弱的外显率;与免疫异常相关;对生殖影响很少或没有影响。
可用群体和家系研究来证实 HLA复合体内标记基因与各种疾病状态的相关性。因为群体研究易于进行,所以有关资料多来源于这种研究。具体疾病与某些具体HLA抗原的相关性通过计算相对风险性(RR)来定量,相对风险性可表述为具有疾病相关 HLA抗原的个体与缺乏这种抗原的个体相比发生该种疾病的机会。RR越高,在病人群体中该抗原的频率越高。以具有 HLA-B27的强直性脊椎炎病人为例。美国患该病的高加索人种90%具有HLA-B27,美国高加索人种对照者的具有率接近9%。RR=(90×91)÷(10×9)=91。因此,HLA-B27阳性个体发生该病的风险性是 HLA-B27阴性个体的91倍。因为不同种族之间某种抗原的频率通常有明显的不同。所以有必要在同种族中比较病人和对照者。例如,HLA-B27见于48%的患强直性脊椎炎的美国黑人,只见于 2%美国黑人对照者,则RR=45。
已经发现很多疾病与某种抗原相关。 HLA与疾病的相关性可分为统计学上较为肯定的相关性、可能相关性及潜在相关性几组。具肯定相关性的有:强直性脊椎炎(抗原为B27)、赖特尔氏综合征(B27)、急性前葡萄膜炎(B27)、青少年风湿性关节炎(B27)、乳糜泻(B8)、突眼性甲状腺肿(B8,DW3)、重症肌无力(B8,DR3)、疱疹性皮炎(B8)、慢性活动性肝炎(BW6,DR8)、青少年糖尿病(B8,DR3)、多发性硬化(B7,DR2)。具可能相关性的有:系统性红斑狼疮(DR3)、天疱疮(B13)、自身溶血性贫血(A3)、脊髓灰质炎(A3,B7)、贝赫切特氏病(B5)。具有潜在相关性的有:急性淋巴细胞白血病(A2,B12)、慢性肾小球肾炎(A2)、麻风(B14)、霍奇金氏病(A1,B8)。
为了解释HLA与疾病的相关性,已提出多种假说。①结合假说:或称受体假说 HLA抗原为细胞膜的特异受体,可与致病因子(如病毒)结合,引起免疫性或非免疫性组织损伤。②连锁基因假说:HLA抗原伴随具体疾病,但其本身并不直接涉及致病作用,而只是与具体疾病的易感基因相连锁。③分子拟态假说:某些病原体(如病毒)抗原在分子结构上与特定HLA抗原相似,因此可形成交叉耐受,机体不能产生免疫攻击,以终止疾病过程。另一相反的观点认为机体对病原体可发生强的免疫应答,由于病原体与HLA抗原的相似性,免疫应答同时针对外来抗原和HLA抗原,这种自身免疫应答导致疾病。某些微生物与某些 HLA抗原间的交叉反应性的资料支持这种理论。④HLA基因与Ir基因连锁:Ir基因控制寄主对病毒或其他抗原的免疫反应性。人的Ir基因就是HLA复合体的D区基因。特定的 HLA-D区基因型可能导致特定的异常免疫反应性。这种异常免疫反应性将导致对某些疾病的易感性或抵抗性。
在不同的HLA-疾病相关中,可有不同的机理发挥作用,并且疾病的发生可能不止一种机理发挥作用。