生物膜离子通道(卷名:生物学)
ion channels of biomembrane
各种无机离子跨膜被动运输的通路。生物膜对无机离子的跨膜运输有被动运输(顺离子浓度梯度)和主动运输(逆离子浓度梯度)两种方式。被动运输的通路称离子通道,主动运输的离子载体称为离子泵。生物膜对离子的通透性与多种生命活动过程密切相关。例如,感受器电位的发生,神经兴奋与传导和中枢神经系统的调控功能,心脏搏动,平滑肌蠕动,骨骼肌收缩,激素分泌,光合作用和氧化磷酸化过程中跨膜质子梯度的形成等。
研究简史 在生物电产生机制的研究中发现了生物膜对离子通透性的变化。1902年J.伯恩斯坦在他的膜学说中提出神经细胞膜对钾离子有选择通透性。1939年A.L.霍奇金与A.F.赫胥黎用微电极插入枪乌贼巨神经纤维中,直接测量到膜内外电位差。1949年A.L.霍奇金和B.卡茨在一系列工作基础上提出膜电位离子假说,认为细胞膜动作电位的发生是膜对纳离子通透性快速而特异性地增加,称为“钠学说”。尤其重要的是,1952年A.L.霍奇金和A.F.赫胥黎用电压钳技术在枪乌贼巨神经轴突上对细胞膜的离子电流和电导进行了细致地定量研究,结果表明Na+和K+的电流和电导是膜电位和时间的函数,并首次提出了离子通道的概念。他们的模型 (H-H模型)认为,细胞膜的K+通道受膜上4个带电粒子的控制,当4个粒子在膜电场作用下同时移到某一位置时,K+才能穿过膜。设n是一个粒子位于通道位置的几率,则 K+电导为:
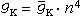 (1)假定参数n服从一阶反应方程,则有:
(1)假定参数n服从一阶反应方程,则有: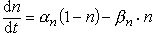 (2) 对Na+电导,认为有3个带电粒子同时控制通道的开放(活化),每个粒子位于活化位置的几率是m,一个粒子负责通道的失活,失活几率是1-h,则通道的开放几率为m3h,Na+电导可写为:
(2) 对Na+电导,认为有3个带电粒子同时控制通道的开放(活化),每个粒子位于活化位置的几率是m,一个粒子负责通道的失活,失活几率是1-h,则通道的开放几率为m3h,Na+电导可写为: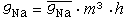 (3)同样,对m和h也采用一阶反应方程的形式,则有:
(3)同样,对m和h也采用一阶反应方程的形式,则有: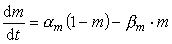 (4)
(4)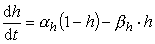 (5)以上各式中,
(5)以上各式中,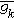 和
和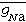 为最大K+和Na+电导,αn、βn、αm、βm、αh、βh分别表示相关过程的速率常数,它们是膜电位的函数。
为最大K+和Na+电导,αn、βn、αm、βm、αh、βh分别表示相关过程的速率常数,它们是膜电位的函数。另一方面,1955年,卡斯特罗和B.卡茨对神经-肌肉接头突触传递过程的研究发现:突触后膜终板电位的发生,是由于神经递质乙酰胆碱(Ach)作用于终板膜上受体的结果,从而确认了受化学递质调控的通道。60年代,用各种生物材料对不同离子通透性的研究表明,各种离子在膜上各自有专一性的运输机构,曾经提出运输机构是载体、洞孔和离子交换等模型。1973年和1974年,C.M.阿姆斯特朗、F.贝萨尼利亚及R.D.凯恩斯、E.罗贾斯两组分别在神经轴突上测量到与离子通道开放相关的膜内电荷的运动,称为门控电流,确认了离子通道的开放与膜中带电成分运动的依从性。1976年E.内尔和B.萨克曼创立了离子单通道电流记录技术,并迅速得到推广应用,近年用这种技术发现了一些新型离子通道,为深入研究通道的结构和功能提供了有力的工具。
80年代初,学者们先后从细胞膜上分离和纯化了一些运输离子的功能性蛋白质,并在人工膜上成功地重建了通道功能,从而肯定了离子通道实体就是膜上一些特殊蛋白质分子或其复合物。近年,科学家应用基因重组技术研究离子通道的结构,1982和1984年,纽莫及合作者先后测定了N型Ach受体和 Na+通道蛋白的氨基酸序列。
研究方法 离子通道结构和功能的研究需综合应用各种技术,包括:电压和电流钳位技术、单通道电流记录技术、通道蛋白分离、纯化等生化技术、人工膜离子通道重建技术、通道药物学、基因重组技术及一些物理和化学技术。
电压钳位技术 一般而言,膜对某种离子通透性的变化是膜电位和时间的函数。用玻璃微电极插入细胞内,利用电子学技术施加一跨膜电压并把膜电位固定于某一数值,可以测定该膜电位条件下离子电流随时间变化的动态过程。利用药物或改变细胞内外的溶液成分,使其他离子通道失效,即可测定被研究的某种离子通道的功能性参量,分析离子电流的稳态和动力学与膜电位、离子浓度等之间的关系,可推断该种通道的电导、活化和失活速率、离子选择性等,并能测量和分析通道的门控电流的特性。
单通道电流记录技术 又称膜片钳位技术,用特制的玻璃微吸管吸附于细胞表面,使之形成10~100GΩ的密封(giga-seal),被孤立的小膜片面积为μm2量级,内中仅有少数离子通道。然后对该膜片实行电压钳位,可测量单个离子通道开放产生的pA(10-12安培)量级的电流,这种通道开放是一种随机过程。通过观测单个通道开放和关闭的电流变化,可直接得到各种离子通道开放的电流幅值分布、开放几率、开放寿命分布等功能参量,并分析它们与膜电位、离子浓度等之间的关系。还可把吸管吸附的膜片从细胞膜上分离出来,以膜的外侧向外或膜的内侧向外等方式进行实验研究。这种技术对小细胞的电压钳位、改变膜内外溶液成分以及施加药物都很方便。
通道药物学研究 应用电压钳位或单通道电流记录技术,可分别于不同时间、不同部位(膜内侧或外侧)施用各种浓度的药物,研究它们对通道各种功能的影响。结合对药物分子结构的了解,不但可以深入了解药物和毒素对人和动物生理功能作用的机制,还可以从分子水平得到通道功能亚单位的类型和构象等信息。
通道蛋白分离、通道重建和基因重组技术 利用与通道特异结合的毒剂标记,可把通道蛋白质从膜上分离下来,经过纯化,可以测定各亚单位多肽的分子量。然后,把它们加入人工膜,可重新恢复通道功能。用于确定蛋白质氨基酸序列的基因重组技术的程序是:从细胞中分离出含有与该种通道蛋白相关的mRNA,置入某种细胞(如大肠杆菌),经逆转录得到cDNA。用限制性内切酶将cDNA切割成特定片段,再用核酸杂交方法钓出特定的DNA并克隆化。通过测定阳性克隆DNA的核苷酸顺序,推断出相应的蛋白质氨基酸序列。
类型和功能特征 离子通道依据其活化的方式不同,可分两类:一类是电压活化的通道,即通道的开放受膜电位的控制,如Na+、Ca+、Cl-和一些类型的K+通道;另一类是化学物活化的通道,即靠化学物与膜上受体相互作用而活化的通道,如 Ach受体通道、氨基酸受体通道、Ca+活化的K+通道等。
钠通道 各种生物材料中,与电兴奋相关的Na+通道有相似的基本特征。通道活化时间常数小于1毫秒,失活时间常数为数毫秒,Na+电流的反转电位约+55毫伏。单通道电流记录显示,Na+单通道电导为4~20pS,平均开放寿命数毫秒。Na+通道对一价碱金属离子的选择通透性比率是:
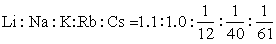 根据一些药物和毒素对Na+通道功能的不同影响,可分为4种类型:①通道阻断剂,如河豚毒素(TTX)、石房蛤毒素(STX)。②通道活化增强剂,如β-蝎毒、箭毒蛙毒素(BTX)、藜芦碱毒素(VER)等。③通道活化抑制剂,如一些局部麻醉剂及其衍生物。④通道失活抑制剂,如链霉蛋白酶、N-溴乙酰胺(NBA)等。
根据一些药物和毒素对Na+通道功能的不同影响,可分为4种类型:①通道阻断剂,如河豚毒素(TTX)、石房蛤毒素(STX)。②通道活化增强剂,如β-蝎毒、箭毒蛙毒素(BTX)、藜芦碱毒素(VER)等。③通道活化抑制剂,如一些局部麻醉剂及其衍生物。④通道失活抑制剂,如链霉蛋白酶、N-溴乙酰胺(NBA)等。钾通道 根据功能特性的不同,K+通道可分为以下类型:①慢(延迟)K+通道(K通道),也就是H-H模型中的K+通道。单通道电流记录显示,单个K通道电导在2~20pS,通道平均开放寿命为数十毫秒。该种通道可被四乙胺(TEA)等特异性阻断,通道对K+有高度选择性,但R姾和NH嬃亦能通过,这种通道在神经轴突和骨骼肌细胞膜中有较高密度。②快(早期)K+通道(A通道),该种通道外向的K+流在膜去极化的早期就出现,表明通道的活化时间常数比慢K+通道小得多,但在-40毫伏以上该通道即关闭。电压钳位实验表明,其宏观电流动力学与Na+电流相似。较低浓度的4-氨基吡啶即能阻断该通道,它也可被四乙胺阻断。③Ca2+活化的K+通道〔K(Ca)通道〕,该种通道的开放,不但与膜电位有关,而且依赖于细胞内Ca2+的浓度,每个通道需结合两个Ca2+才能活化。单通道电导可高达300pS,并有较长的开放寿命,这种通道与Ca2+通道协同作用,对调节细胞膜电兴奋性的节律有重要意义。它可被四乙胺、N'-四乙酸(EGTA)、奎尼丁和Ba2+阻断。④内向整流的K+通道,其特征是:在膜超极化时通道开放与膜电位和胞外K+浓度密切相关,通道开放时产生内向K+电流,单通道电导在5~10pS范围。
钙通道 Ca2+通道广泛存在于各种生物组织的细胞膜中。宏观的Ca2+电流动力学特征与Na2+电流相似,但峰值小且失活过程慢,可达数十到数百毫秒。Ca2+通道对Ca2+、Ba2+、Sr2+都有高通透性,但Ni2+、Cd2+、Co2+、Mn2+等离子能有效地阻断Ca2+通道。药物对Ca2+通道的作用可分为:①通道阻断或抑制剂,可分为苯烷基胺类(如异博定、甲基异博定D600)、苯硫氮类、双氢吡啶类等类型。②通道激活剂,一些双氢吡啶化合物如BayK8644等药物可活化Ca2+通道。近年,对小鸡背根神经节细胞的研究发现有3种类型的Ca2+通道:①L型,该种通道在膜电位大于-20毫伏时活化,电流失活缓慢。单通道电导约25pS。②T型,膜电位约-60毫伏时通道即活化,-10毫伏以上通道电流幅值反而下降,单通道电导约8pS。③N型,该种通道在膜电位不小于-10毫伏才能活化,但又必须超极化到-80毫伏以下才能克服通道的失活。电流动力学比 L型快但比T型慢,单通道电导约13pS。以上3类Ca2+通道在不同细胞膜上选择性分布及密度的差别,将影响各种细胞的生理功能。Ca2+通道除了对细胞电兴奋性有贡献外,它通过调节细胞内Ca2+浓度,可进一步调节许多细胞功能。
N型乙酰胆碱受体通道 它是由神经递质Ach活化的正离子通道。当突触前膜一次量子化释放数千个 Ach分子,它们作用于突触后膜上的N型受体时,受体通道开放,产生Na+和K+电流,引发突触后膜一个小终板电位(mEPP)。N-AchR单通道电导在20~60pS范围,平均开放寿命数毫秒,通道电流反转电位约-10毫伏,近年发现该种通道有多种电导态(见图)。
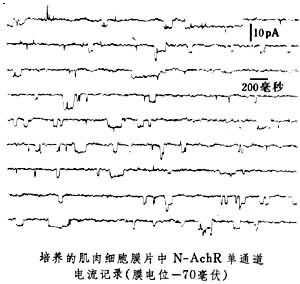 通道的离子选择性较差,可允许数十种无机和有机正离子通过,许多毒素和有机物能阻断或抑制该种通道,α-银环蛇毒(α-BGTX)是 N型Ach受体通道的特异性阻断剂。
通道的离子选择性较差,可允许数十种无机和有机正离子通过,许多毒素和有机物能阻断或抑制该种通道,α-银环蛇毒(α-BGTX)是 N型Ach受体通道的特异性阻断剂。80年代以来,已发现多种由神经递质和激素活化的受体通道,如谷氨酸受体通道、多巴胺受体通道、5-羟色胺受体通道、γ-氨基丁酸受体通道等。
分子构象和门控动力学 离子通道研究的前沿是试图从分子水平揭示通道蛋白的空间构象、构象变化与通道门控动力学之间的关系。
N-AchR通道 已测定了受体蛋白质分子量是250000,并测定了它的全部氨基酸序列,确证该受体通道由
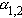 、α、γ和δ5个亚基组成,这4种亚基有相似的氨基酸顺序,但只有α亚基上有 α-BGTX的特异结合位点。一种构象模型是:5个亚基各有若干个α螺旋跨膜排列,共同形成五瓣状的蛋白质复合物,两个α亚基间是亲水的离子通道,通道开口约25埃,中间是6~7埃的狭窄孔道,其中排列有负电性氨基酸残基侧链。当两个 Ach分子分别结合于两个α亚基特定位点后,引起局部构象变化,使通道开放,可用以下模式描述其动力学过程:
、α、γ和δ5个亚基组成,这4种亚基有相似的氨基酸顺序,但只有α亚基上有 α-BGTX的特异结合位点。一种构象模型是:5个亚基各有若干个α螺旋跨膜排列,共同形成五瓣状的蛋白质复合物,两个α亚基间是亲水的离子通道,通道开口约25埃,中间是6~7埃的狭窄孔道,其中排列有负电性氨基酸残基侧链。当两个 Ach分子分别结合于两个α亚基特定位点后,引起局部构象变化,使通道开放,可用以下模式描述其动力学过程: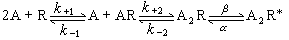 式中A是Ach分子,R是受体,
式中A是Ach分子,R是受体, 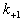 等分别表示每一步反应的速率常数。
等分别表示每一步反应的速率常数。钠通道 从电鳗电板分离的钠通道蛋白质分子量是208321,是由1820个氨基酸组成的多肽序列,可分为4个相似的区段,每个区段中分别有较集中的正电性和负电性的氨基酸序列节段。多种钠通道构象模型的共同特征是:由多个α螺旋跨膜排列组成通道,通道内侧应富含极性的氨基酸残基侧链,每个通道的控制部分由离子选择性滤器、活化闸门和失活闸门3部分组成,其实体是氨基酸侧链的极性基团。膜电位变化时,电场诱导极性基团运动,使通道局部构象发生变化,导致通道的开放、失活或关闭,并产生门控电流。一种动力学模式如下:
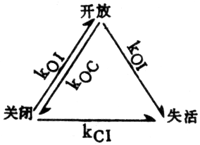 kCO等分别表示各种状态转化的速率常数。关于关闭、活化和失活3种状态之间的转化,有两种观点:一种认为通道从关闭态必须经活化态才能转化为失活态(偶联方式),另一种认为从关闭态可以直接转化为失活态(非偶联方式),目前非偶联方式得到较多的实验事实支持。
kCO等分别表示各种状态转化的速率常数。关于关闭、活化和失活3种状态之间的转化,有两种观点:一种认为通道从关闭态必须经活化态才能转化为失活态(偶联方式),另一种认为从关闭态可以直接转化为失活态(非偶联方式),目前非偶联方式得到较多的实验事实支持。参考书目
B.Hille,Ionic Channels of Excitable Membranes,Plenum Press,New York,1984.
R.Lalorre ed.,Ionic Channels in Cells and Model Systems, Plenum Press,New York,1986.
B.Sakmann, E.Neher,Single-Channel Recording, Plenum Press,New York,1983.