下丘脑-垂体系统(卷名:生物学)
hypothalamus-hypophysis system
下丘脑与脑下垂体组成的一个完整的神经内分泌功能系统。此系统可分两部分: ①下丘脑-腺垂体系统。是神经、体液性联系,即指下丘脑促垂体区的肽能神经元通过所分泌的肽类神经激素(释放激素和释放抑制激素),经垂体门脉系统转运到腺垂体,调节相应的腺垂体激素的分泌;②下丘脑-神经垂体系统。有直接神经联系,下丘脑视上核和室旁核的神经内分泌细胞所分泌的肽类神经激素可以通过轴浆流动方式,经轴突直接到达神经垂体,并贮存于此(图1)。
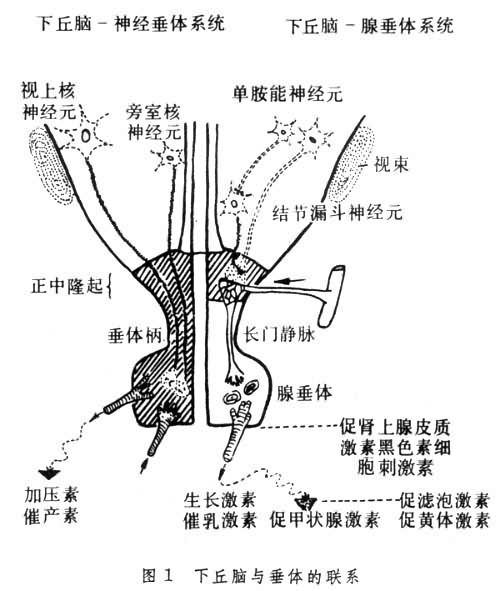
下丘脑是间脑的一部分,左右对称,形成第三脑室下部的侧壁和底部。上界以水平走行的下丘脑沟与丘脑为界,外侧以大脑基底神经节、底丘脑和视束为界,下方经垂体柄与垂体相连。下丘脑具有许多细胞核团和纤维束,与中枢神经系统的其他部位具有密切的相互联系。垂体又名脑下垂体,分泌的激素除直接调节某些靶器官的功能活动外,还有几种促激素作用于相应的靶腺(如甲状腺、性腺、肾上腺皮质等),调节靶腺的分泌功能,并经靶腺激素间接调节某些器官的生理功能。切断下丘脑与神经垂体的联系以后,神经垂体中的激素(催产素及抗利尿激素)会逐渐消失,而中枢端以上却贮留大量分泌颗粒。这说明,过去认为的两种神经垂体激素实际上是由下丘脑合成的,经神经轴突转运到神经垂体。下丘脑视上核和室旁核在合成两种神经激素的同时,还产生一种大分子蛋白质,是神经垂体激素的载体蛋白,称为神经垂体素,神经垂体素与两种神经垂体激素同时经轴突转运至神经垂体,释放入血。下丘脑促垂体区的某些神经元,还具有内分泌功能。这些神经内分泌细胞可以产生九种肽类神经激素(释放激素及释放抑制激素),经神经轴突转运到中隆区,并由此进入垂体门脉循环,再运到腺垂体,调节相应的腺垂体激素的分泌。因为脑下垂体在功能上与下丘脑紧密相连,所以一般将下丘脑与垂体看作为一个完整的功能系统。
垂体的功能结构 垂体位于大脑之下,有短柄(垂体柄)与脑底相连。它深藏于蝶鞍背面的小骨腔(垂体窝)内,成年人的垂体像一颗大豌豆,重约0.6克。垂体可分腺垂体和神经垂体两部分,它们在发生上、组织学上和功能上都不同(图2)。

腺垂体的血液供应主要来自垂体门脉系统。门静脉的两端都形成毛细血管网。自颈内动脉发出的分枝进入正中隆起和垂体柄近端细分为毛细血管网,随后毛细血管又汇合形成数条平行的静脉,叫垂体门静脉,门静脉沿垂体柄前面和腹侧面下行到垂体前部后,在腺细胞之间形成丰富的血窦,腺细胞与血液之间仅以窦壁内皮细胞层和窦周围间隙相隔。这个血管系统是下丘脑调节腺垂体的主要神经体液途径。神经垂体的血液供应来自垂体下动脉。两部分垂体的静脉血液通过数条静脉注入附近的静脉窦。
垂体的神经支配来自颈上交感神经节的节后纤维和面神经的岩神经分支(副交感神经纤维);以上两种纤维可能都是血管运动神经;此外,还有下丘脑纤维,主要来自下丘脑的视上核和室旁核以及结节区和乳头体等处,统称下丘脑-垂体束。根据解剖生理特点,将下丘脑-垂体束又分为视上-垂体束和结节-垂体束,它们是两个独立的神经分泌系统。前者的纤维主要来自视上核和室旁核,止于神经部。后者的纤维主要来自结节部,止于正中隆起和漏斗柄。上述所有不同来源的神经纤维,主要分布于神经垂体,仅有少数进入中间部,几乎没有分泌纤维支配垂体前部。
垂体的个体发生和种系发生 整个垂体起源于外胚层,但组织的起源是不同的。神经垂体起源于脑漏斗。脑漏斗是下丘脑的一个外突囊;前叶和中叶是由上颌的突起──拉克氏囊衍生而来的。在胚胎发育过程中,拉克氏囊迅速与脑漏斗相遇并失去它同颌上皮的联系,成为垂体残存的空腔。这个空腔在前叶和中叶之间;某些动物可以完全消失。中间部是由拉克氏囊壁分化来的,它同脑漏斗相接触,囊的其余部分大大变厚而成为远侧部,两侧的延伸物最后融合形成一层薄的组织板,叫做结节部,它被包在漏斗柄的周围并延长到下丘脑的正中隆起。
垂体在所有脊椎动物中都有类似的结构。从种系发生上看,有3个明显的变化趋势:①神经垂体趋于集中化;②由正中隆起流向腺垂体的垂体门脉系统的出现和完善化;③鸟类和某些哺乳动物垂体中叶的消失(图3)。
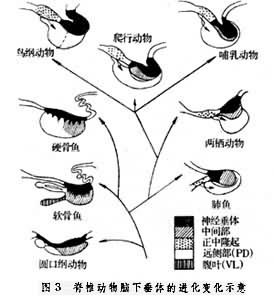
圆口纲动物的腺垂体为一长方体结构,由 3部组成:即前腺垂体、中腺垂体和后腺垂体。神经垂体是第三脑室底部稍加厚的部分,通过一层很薄的血管组织与腺垂体分开。某些下丘脑神经分泌轴突围绕着血管终止于神经垂体中,下丘脑合成的精氨酸血管加压素由此释出。
软骨鱼的神经垂体是散开的,并且与中间部交织在一起,通常合称神经中间部。视前核和下丘脑侧部的神经细胞分泌物,经过轴突释放到神经垂体,在神经垂体后部有一高度血管化的血管囊组织,功能还不清楚。远侧部位于漏斗部下面,可分近侧区和吻侧区;腹叶是板鳃鱼类腺垂体所特有的结构,大小和形状在不同种间差别很大,功能还不清楚。
硬骨鱼的腺垂体由中间部和前叶组成。前叶有时按其位置分为吻侧区和近侧区,大致含有哺乳动物腺垂体的各类细胞。硬骨鱼的神经垂体也像软骨鱼类的,是散开的,并同中间部和远侧部的细胞交织在一起。来自视前核和侧结节核的神经分泌纤维终止于神经垂体的各个部分,在远侧部也可以发现其分泌物。硬骨鱼是否存在正中隆起和垂体门脉系统的类似结构还有争论。
肺鱼的神经垂体和中间部已明显分开,没有结节部,远侧部和中间部之间有一裂缝,没有明显的血管囊。在肺鱼已出现正中隆起和一个原始的门脉系统,由正中隆起流向远侧部。
两栖纲动物的垂体很像肺鱼的垂体。无尾两栖纲动物的正中隆起较发达,门脉系统明显。某些两栖动物具有门脉系统,血液可流向中间部和神经部。
爬行动物没有结节部,但在中间部与远侧部之间存在着一条典型的缝隙;鸟类没有中间部,神经叶和前叶被结缔组织形成的膈分开。鸟类的神经垂体是高度特化的。有些鸟类,正中隆起外区及神经部是下丘脑神经元神经分泌物的贮藏所。
下丘脑-腺垂体系统 腺垂体的激素及其生理功能 从腺垂体中已经分离出8种蛋白质激素,即生长激素(CH),促肾上腺皮质激素(ACTH),促甲状腺激素(TSH),生乳素(PRH),卵泡刺激素(FSH),黄体生成素(LH),β-促脂激素(β-LPH)和黑素细胞刺激素(MSH)。
①生长激素(GH) 根据对GH分子量、氨基酸组成和部分氨基酸序列的比较研究已知各种动物生长激素的分子量非常接近。牛、羊、马、猪、鲸、狗、大鼠、家兔、 猴和人10种动物的GH分子量大致都在 22000左右。等电点有相当大的差异,其范围从4.9(人)到6.8(羊和牛)。各种动物GH末端的氨基酸也都十分相似。
GH的生理作用有相当大的种属差异性。灵长类的GH对人和猴都有效,而其他任何脊椎动物的GH对灵长类则无效;牛的GH对鱼有效,而鱼的GH对大鼠和其他哺乳动物则无效。大鼠对于除鱼以外的所有脊椎动物的GH皆有反应。GH分子结构上的免疫活性中心同生物活性中心可能不一致。
GH的主要生理功能是:刺激软骨及软组织增生,使骨骼面积增加,伴随内脏增大,肌肉、皮肤、结缔组织和淋巴器官增生;促进蛋白质合成;给哺乳动物反复注射GH时,可使血糖升高,长期注射,可导致永久性糖尿病;刺激胸腺淋巴细胞和一般淋巴细胞的繁殖;当将GH同其他几种“促激素”(如ACTH,TSH,FSH和LH)分别结合使用时,可显著加强这些激素的效能;对其他激素有“允许作用”,能产生为其他激素或因子充分发挥作用的生理环境等。
② 生乳素(PRH):已经从羊、牛和人的垂体中分离出高纯度的PRH,其分子结构、生物活性、免疫和电泳性质相似,分子量都接近约24000,但其溶解度和酪氨酸含量稍有差异。 羊的PRH和人的GH大部分分子片断相似。羊的PRH由198个氨基酸组成,比人的GH分子稍大。根据多方面分析(包括化学结构的相似性,生物活性的重叠性和免疫关系),各种脊椎动物的PRH和GH可能都是由同一种或几种原始分子演化而来。 PRH的主要生理功能是调节生殖活动和性行为,但随动物种属不同而异;可明显影响两栖动物的迁水现象,如用PRH处理去垂体的蝾螈,在4~10天之内便迁入水中;刺激鸟类羽毛突产生新羽毛,它与雌激素协同作用产生孵化斑;刺激嗉囊发育,使嗉汁充盈,抑制性活动,对雌、雄鸟都有抗性腺活动的作用;刺激大鼠和小鼠的黄体分泌孕酮;促进已发育好的乳腺分泌乳汁等。
③ 促肾上腺皮质激素(ACTH):从牛、羊、猪和人的垂体中已分离出高纯度的ACTH。这 4种来源的ACTH都是直链多肽,由39个氨基酸组成,N末端有一个丝氨酸,C末端有一个苯丙氨酸,分子量约4500。虽然在分子结构上有种属差异,但刺激肾上腺皮质的能力相同。种属差异限于分子第25到33位上的氨基酸,但分子的生物活性并不位于这些氨基酸上。去掉分子第25位以后的氨基酸,不影响激素的生物活性。ACTH氨基端1~24氨基酸为ACTH功能部分,没有种属差异。它的羧基末端25~39氨基酸与ACTH的生物活性无关。ACTH已能人工合成。ACTH主要作用于肾上腺皮质的束状带和网状带,使其细胞增生,并促进糖皮质激素的生物合成和分泌。此外,有人发现,ACTH也能促进肾上腺髓质激素的合成,这一作用部分是ACTH对酪氨酸羟化酶的直接作用,部分是通过糖皮质激素实现。
④ 促脂激素(LPH):β促脂激素(β-LPH)由91个氨基酸组成,分子量约9500。分子中41~58片断中的18个氨基酸与牛羊等动物的黑素细胞刺激素相同。α促脂激素(α-LPH)由58个氨基酸组成,与β-LPH前1~58个氨基酸相同。人的垂体能合成和分泌β-LPH和α-LPH。
LPH具有溶脂作用和轻微的黑素细胞的刺激作用。有人已从脑中提取出具有镇痛作用的五肽化合物,这些化合物中的一种和β-LPH的第61~65个氨基酸相同。已经发现β-LPH羧基末端的片断(其中包括61~65)较上述五肽的镇痛作用强约20倍,因而提出:LPH羧基末端片断可能是一种天然的具有镇痛作用的神经递质,而β-LPH可能是它的前身物。
⑤ 黑素细胞刺激素 (MSH):MSH有α和β两种。在鱼类、两栖类和爬行类是由垂体中间叶分泌,因此也叫垂体中叶激素。α-MSH由 13个氨基酸组成,与ACTH分子的前13个氢基酸相同。某些哺乳动物(包括人类)并不分泌α-MSH。MSH的主要功能是刺激黑素细胞色素体,使之扩散,导致皮肤变黑。
⑥ 糖蛋白质激素〔黄体生成素(LH)、卵泡刺激素(FSH)和促甲状腺激素(TSH)〕:在腺垂体中有3种糖蛋白质激素,即LH、FSH和TSH。这些激素都含有碳水化合物的侧链。分子量约为 3万。分子由两个化学结构不同的亚单位组成。两者间由非共价键结合。经特殊处理,两部分很易分开。分开的亚单位不具有生物活性。β亚单位具有决定激素分子特异性的构型特征。 3种糖蛋白质激素的 α亚单位相同。不同激素的 α和β亚单位的重组,并不破坏激素的活性,但是杂交分子的特性总是同β亚单位相关的。
在硬骨鱼垂体中发现有一种异促甲状腺因子 (HTF),与促性腺激素十分相近,HTF、LH和FSH对硬骨鱼的甲状腺都有刺激作用,这些发现提示,TSH、LH和FSH可能起源于同一种原始分子。LH和FSH对性腺均有刺激作用(见雌性生殖系统、雄性生殖系统)。TSH主要刺激甲状腺合成和分泌甲状腺激素(见甲状腺)。
下丘脑对垂体促激素的调节 下丘脑的释放激素(RH)可使相应的腺垂体激素合成及分泌增加,如促甲状腺激素释放激素可促进腺垂体促甲状腺激素的合成和分泌,下丘脑抑制激素能抑制腺垂体相应激素的合成及分泌;如生乳素抑制激素可使腺垂体生乳素合成及分泌减少,加压素可提高血压,促进肾小管对水的吸收。这 9种神经激素中,有4种已知其化学结构,其中3种已能人工合成,并用于生产实践,5种尚未弄清结构的叫做释放(或抑制)因子(见表)。已弄清结构并能人工合成的有 3种:促甲状腺激素释放激素(TRH,3肽):焦谷-组-脯-N啹。黄体生成素释放激素(LH-RH,10肽): 焦谷-组-色-丝-酪-甘-亮-精-脯-甘-N啹。生长激素释放的抑制激素(GH-RIH,14肽): H-丙-甘-半胱-赖-门酰-苯丙-苯丙-色-赖-苏-苯丙-苏-丝-半胱-OH。
利用已知结构并已人工合成的 3个肽类神经激素进行了广泛的研究,并且阐明这些神经激素具有以下一些共同特点:
① 化学性质。除PIF可能是多巴胺外,它们可能都是肽类激素,推测的分子量在1000~2500之间。
② 合成部位。它们都是在下丘脑促垂体区的神经元中合成的。合成每一种肽类神经激素的部位不全一致,但有相互重叠。
③ 含量。每种肽类神经激素在下丘脑中的含量都很少,大约每种肽类激素在每个下丘脑中的含量仅为20纳克左右。
④ 转运。这些肽类激素可经垂体门静脉转运至腺垂体,也可能被第三脑室的特化的脑室膜细胞摄取,经脑脊液再进入血液循环。 脑脊液中也含有高浓度的TRH和LH-RH。
⑤ 非下丘脑所特有。如在下丘脑以外的脑区中也存在TRH及LH-RH,提示它们可能也起到神经递质的作用。在胃肠道中也有TRH,并可能参与调节胃肠道的活动。此外,胎盘中也发现有TRH。下丘脑以外也有LH-RH,胎盘中存在的LH-RH可能参与调节人绒毛膜促性腺激素(HCG)的合成及分泌。最近报道在两性的性腺中可能也存在某种类似于LH-RH的肽。 生长素抑制激素广泛存在于胃、肠、胰系统中。甚至在某些神经元中肽类激素与经典的神经递质共存。
⑥ 作用并不专一。最初认为下丘脑的肽类激素作用十分专一,并依其作用而定名。实际上它们的作用并不专一,相反却十分广泛。如LH-RH不仅刺激腺垂体LH的合成和分泌,而且也刺激FSH的合成和分泌。TRH不仅促进TSH的合成和分泌,也有明显的促乳汁分泌作用。生长激素抑制激素的作用则更为广泛,它不仅抑制腺垂体生长激素的合成和释放,而且还抑制TSH、 ACTH、生乳素、胰高血糖素、胰岛素、胃泌素及促胰液素的分泌,因而有人称之为激素的抑制物。
下丘脑-神经垂体系统 神经垂体是神经组织,主要由大量无髓鞘神经纤维、神经胶质以及由后者演变而来的垂体细胞所组成。神经垂体在结构与功能上都与下丘脑密切相关。从下丘脑视上核和室旁核的神经元发出的神经纤维直接进入神经垂体,叫做下丘脑垂体束。由视上核和室旁核的神经元合成和和分泌的激素(抗利尿激素及催产素),即沿此束被运送至神经垂体贮藏,需要时再释放入血液循环。
激素及生理作用 视上核与室旁核如何合成抗利尿激素和催产素,其细节目前尚不十分清楚。一般认为,视上核可能以产生抗利尿激素为主,而室旁核以产生催产素为主。
抗利尿激素和催产素都是由九个氨基酸组成的小肽,除第3位和第8位上的两个氨基酸残基不同外,这两种激素的分子结构基本相同,因此两者在生理作用上也有交叉。
① 抗利尿激素:也叫加压素。其主要生理作用是促进肾脏集合管和远曲小管后段对水分子的重吸收。因此,抗利尿激素分泌不足、分泌过多以及肾小管对抗利尿激素反应失常都会影响尿量。例如,给身体内水分充足的人仅仅注射几百个微单位的抗利尿激素便可使尿量明显减少,尿液呈高渗。抗利尿激素缺乏时,每天尿量可多至十几升,称为尿崩症。
② 催产素:对妊娠子宫,催产素有强烈刺激子宫肌收缩的作用,但它对分娩过程究竟有何作用,至今尚无法定论(见妊娠)。催产素有促进乳腺排乳的作用(见泌乳)。此外,催产素对母鸡产卵,鱼生子的过程可能也有促进作用。
分泌的调节 ①抗利尿激素:在生理情况下,主要受血容量和细胞外液渗透压变化的调节。②催产素:刺激外生殖器、吸吮乳头与刺激子宫均可反射地引起催产素分泌,并伴有子宫收缩,对精子运行至输卵管有促进作用。
分娩时对子宫颈和阴道的牵拉可反射性地引起催产素的释放,使子宫肌收缩加强,一方面促进胎儿娩出,一方面又引起子宫颈和阴道更大的牵张,从而引起更多的催产素的释放,形成正反馈,直至胎儿娩出。
吸吮时,对乳头的触觉刺激,可反射地引起催产素的分泌和释放,从而引起排乳,与吸吮或哺乳有关的听或视觉刺激可通过“条件反射”引起催产素的释放。情绪反应如害怕、焦急、疼痛可以抑制催产素的释放,因而阻滞乳汁的排出。
下丘脑的神经递质 下丘脑有多种神经递质,主要为单胺类递质和肽类递质。
体内的单胺类主要包括儿茶酚胺和吲哚类两大部分。

下丘脑单胺能神经元分泌单胺类递质调节着产生释放或抑制激素的肽能神经元的功能。但是,每种单胺类递质的具体作用却存在着很大的种属差异。
下丘脑还含有高浓度的内源性吗啡样物质,它们可能也通过改变肽能神经元的功能活动,调节着多种腺垂体激素的分泌。例如,有实验表明,外源注射吗啡可使血中生乳素水平升高,而LH和FSH的水平下降。吗啡的这一作用是通过吗啡受体而实现的,用吗啡受体拮抗剂可以反转吗啡对垂体生乳素和促性腺激素分泌的影响。吗啡受体拮抗剂可降低血中生乳素的水平并使LH和 FSH的水平升高,这提示,内源性吗啡样物质正常即调节着生乳素及促性腺激素的分泌。
有人在脑内还发现了许多新的肽类物质, 如P物质、神经降压素、血管活性肠肽、胃泌素及胆囊收缩素等,它们可能也调节着垂体激素的释放,但其具体作用及其生理意义都有待进一步研究。
参考书目
程治平:《内分泌生理学》,人民卫生出版社,北京,1984。
C.D.特纳、J.T.巴格内尔著,刘以训等译:《普通内分泌学》,科学出版社,北京,1983。(C.D.Turner,J.T. Bagnara,General Endocrinology,6th ed., W.B.Saunders Co.,Philadelphia,1976.)