分类学(卷名:生物学)
taxonomy
研究生物分类的方法和原理的生物学分支。分类就是遵循分类学原理和方法,对生物的各种类群进行命名和等级划分。分类学曾被称为系统分类学,但它与系统学很易混淆,系统学是研究生物的分异度(多样性)以及它们中间的任何一个类群和其他所有类群的各种关系的科学,曾称为分类系统学。三者的共同目的是从理论上和实践上,阐明种类之间的关系(或亲缘关系),建立自然系统,确定各类群的命名和排序,总结其进化历史。
地球上现生的物种以百万计,千变万化,各不相同,如果不予分类,不立系统,便无从认识,难以利用。分类系统是生物种类的查找系统,可借以认识和查取有关资料。分类的对象是形形色色的种类,都是进化的产物;分类学在于阐明种类之间的历史渊源,使建立的分类系统反映进化历史。因而从理论意义上说,分类学是生物进化的历史总结。
分类学是综合性学科。生物学的各个分支,从古老的形态学到现代分子生物学的新成就,都可吸取为分类依据。分类学亦有其自己的分支学科,如以染色体为依据的细胞分类学(或染色体分类学),以血清反应为依据的血清分类学,以化学成分为依据的化学分类学,等等。动物、植物和细菌,作为三门分类学,各有其特点;病毒分类则尚未正式采用双名制和阶元系统。
分类学历史 人类在很早以前就能识别物类,给以名称。汉初的《尔雅》把动物分为虫、鱼、鸟、兽 4类:虫包括大部分无脊椎动物;鱼包括鱼类、两栖类、爬行类等低级脊椎动物及鲸和虾、蟹、贝类等;鸟是鸟类;兽是哺乳动物。这是中国古代最早的动物分类,四类名称的产生时期看来不晚于西周。这个分类,和林奈(1758)的六纲系统比较,只少了两个纲(两栖纲和蠕虫纲)。
古希腊哲学家亚里士多德采取性状对比的方法区分物类,如把热血动物归为一类,以与冷血动物相区别。他把动物按构造的完善程度依次排列,给人以自然阶梯的概念。
17世纪末叶,英国植物学者J.雷曾把当时所知的植物种类,作了属和种的描述,所著《植物研究的新方法》(1682) 是C.von林奈以前的一本最全面的植物分类总结。J.雷还提出“杂交不育”作为区分物种的标准。
近代分类学诞生于18世纪,它的奠基人是瑞典植物学者林奈。林奈为分类学解决了两个关键问题:①建立了双名制,每一物种都给以一个学名,由两个拉丁化名词所组成,第一个代表属名,第二个代表种名。例如Panthera tigris是虎的学名,Bombyx mori是蚕的学名。②确立了阶元系统。林奈把自然界分为植物、动物和矿物三界,在动植物界下,又设有纲、目、属、种四个级别,从而确立了分类的阶元系统。每一物种都隶属于一定的分类系统,占有一定的分类地位,可以按阶元查对检索。并首先在1753年印行的《植物种志》和1758年第10版《自然系统》中应用于植物和动物。这两部经典著作,标志着近代分类学的诞生。
林奈相信物种不变,他的《自然系统》没有亲缘概念,其中 6个动物纲是按哺乳类、鸟类、两栖类、鱼类、昆虫、蠕虫的顺序排列的。J.-B.de拉马克把这个颠倒了的系统拨正过来,从低级到高级列成进化系统。他还把动物区分为脊椎动物和无脊椎动物两类,并沿用至今。但是,由于他的进化观点在当时没有得到公认,因而对分类学影响不大。直到1859年,C.R.达尔文的《物种起源》出版以后,进化思想才在分类学中得到贯彻,明确了分类研究在于探索生物之间的亲缘关系,使分类系统成为生物系谱──系统分类学由此诞生。
分类阶元、命名模本方法和鉴定 分类系统是阶元系统,通常包括7个主要级别:种、属、科、目、纲、门、界。种(物种)是基本单元,近缘的种归合为属,近缘的属归合为科,科隶于目,目隶于纲,纲隶于门,门隶于界。随着研究的进展,分类层次不断增加,单元上下可以附加次生单元,如总纲(超纲)、亚纲、次纲、总目(超目)、亚目、次目、总科(超科)、亚科等等。此外,还可增设新的单元,如股、群、族、组等等,其中最常设的是族,介于亚科和属之间。林奈晚年承认种内有变种。通常种下分类,动物只设亚种单元;植物设亚种、变种、变型等单元;细菌设品系、菌株等单元。
列入阶元系统中的各级单元都有一个科学名称,表1是两个简化的例子。命名的单元──表上从家蚕到动物界,从马尾松到植物界,不再是抽象的单元,而是含有实际种类的物类单元taxa。分类工作的基本程序就是把研究对象归入一定的系统和级别,成为物类单元。所以分类和命名是分不开的。

种和属的学名后常附命名人姓氏,以标明来源,便于查找文献。表上家蚕,蚕蛾属学名后的L.,是命名人林奈Linnaeus的缩写,马尾松学名后的Lamb.,是命名人兰伯特Lambert的缩写,通常除林奈等少数之外,一般命名者姓名引用时不应缩写。亚种命名采取3名制,即在属名、种名后再加亚种名。例如华南虎和东北虎都是虎的亚种,前者的学名是Panthera tigris amoyensis Hilzheimer,后者是Panthera tigris altaica Temminck。变种学名亦采取 3名制,只是在种名与变种之间加上拉丁文变种varietas的缩写var.,例如中华按蚊原先鉴定为赫坎按蚊的变种(现已分出为独立的种)学名曾是 Anopheleshyrcanes var.sinensis Wedemann。
分类名称要求稳定,一个属或种(包括种下单元)只能有一个学名,如果有两个或多个时,便是“同物异名”,必须于其中核定最早提出的学名,摒弃较后的异名。一个学名只能用于一个对象(属或种),如果有两个或多个对象者,便是“异物同名”,必须于其中核定最早的命名对象,而其他的同名对象则另取新名。这叫做“优先律”,动物和植物分类学界各自制订了《命名法规》,所以在动物界和植物界间不存在异物同名问题,如Cystiphyllum既是褐藻又是古生代珊瑚的属名。“优先律”是稳定学名的重要措施。优先律的起始日期,动物是1758年,植物是1820年,细菌则起始于1980年1月1日。
法规的另一重要规定是模本方法。凡发表新种(包括新属和新亚种),必须指定一个标本作为命名的模式标本,叫做“正模”,简称模本。“副模”是原描述所根据的其他标本,特别是和正模同时同地采得的标本;“配模”是副模中选取的与正模异性的标本。在模本遗失或未定的情况下,可以选取合适标本作为“新模”,以代替正模,一般要从原产地和原层位采集。与模本产地相同而在以后采得的标本可立为“地模”。模式标本是鉴定的对照标本。
鉴定学名是取得物种有关资料的手段,即使是前所未知的新种类,只要鉴定出其分类隶属,亦可预见其一定特征。分类系统是检索系统,也是信息存取系统。许多分类著作,如基于区系调查的动植物志,记述某一国家或地区的动植物种类情况,作为基本资料,都是为鉴定、查考服务的。
物种概念与物种标准 物种指一个动物或植物群,其所有成员在形态上极为相似,以至可以认为他们是一些变异很小的相同的有机体,它们中的各个成员间可以正常交配并繁育出有生殖能力的后代,物种是生物分类的基本单元,也是生物繁殖的基本单元。物种概念反映时代思潮。在林奈时代,人们相信物种是不变的,同种个体符合于同一“模式”。模式概念(不同于模式标本),渊源于古希腊哲学的古老的概念,应用到整个分类系统,概念假定所有阶元系统中的各级物类单元,都各自符合于一个模式。对神创论者来讲,由各级模式所组成的物类阶元,正好说明了“造物主”的智慧设计。模式概念亦是形态学概念,所谓模式是指形态模式──物种是由形态相似的个体所组成的,同种个体符合于同一形态模式。这是物种的形态学定义,也是形态学标准。另一物种标准是杂交不育。早在林奈以前,J.雷就明确指出,物种是繁殖单元,同种个体可以交配繁殖,异种之间则杂交不育,形成生殖隔离。生殖隔离和模式概念一样,符合于物种不变的观点,因为它突出了种间间断,无视历史连续。
林奈时代的物种概念,包含两个基本内容:①不变,②客观存在。由于不变,而且是“上帝”所创造,物种当然是客观存在。种内形态同一,种间生殖隔离则是物种不变的两个标准。达尔文的物种概念与此相反,它的基本内容是:①变,②人为单元。进化论说明了物种是变的,变化中的物种否定了自身的存在。达尔文在《物种起源》内说:“可见‘物种’这名词,我认为是为了方便起见,任意地用来表示一群很相似的个体的,它在本质上和‘变种’没有区别”。达尔文不考虑物种定义,因为物种既是人为单元,便不可能有客观标准,更不需要定义。他的进化论证明了种间的历史连续,却忽视了种间的间断意义。
物种之变与不变曾经是进化论和特创论的斗争焦点,是势不两立的观点。但是,分类学的事实说明,每一物种各有自己的特征,没有两个物种完全相同;而每个物种又保持一系列祖传的特征,据之可以决定其界、门、纲、目、科、属的分类地位,并反映其进化历史。整个阶元系统内各级物类之间的特征关系,表现为祖征与新征的对立统一。界的特征,如细胞壁、叶绿体和光合作用之作为植物特征,对界下单元(门、纲、目、科、属、种)来讲,是祖征;而在界间对比,如植物界与动物界或真菌界对比,则又表现为各界的新征。界下的各级单元,从门到种,亦都如此。
林奈和达尔文的物种概念是个体概念──物种是一群相似的个体。20世纪30、40年代,随着“新系统学”的发展,强调了群体概念。物种不是毫不相干的个体,而是以个体集合为大大小小的种群单元而存在的,物种是“种群”集团,种群是种内的繁殖单元。分类学上当前流行的物种定义,就是以种群为单元的E.迈尔定义:“物种是由自然种群所组成的集团,种群之间可以相互交流繁殖(实际的或潜在的),而与其他这样的集团在生殖上是隔离的”。定义沿用了生殖隔离的标准,它所突出的是群体概念。生殖隔离作为标准,只适用于有性物种,不适用于无性物种和化石物种。对于无性和化石物种,一般是从特征的间断程度,以判断种类划分。一个比较笼统的定义:“物种是生命系统线上的基本间断”,可以适用于一切物种。
分类工作的基本内容是区分物种和归合物种,前者是种级和种下分类,后者是种上分类。种群概念提高了种级分类水平,改进了种下分类,其要点是以亚种代替变种。亚种一般是指地理亚种,是种群的地理分化,具有一定的区别特征和分布范围。亚种分类反映物种分化,突出了物种的空间概念。含有两个或多个亚种的物种称多型种,不分亚种的物种称单型种。形态十分相似但生殖上隔离的物种称姐妹种,又称隐种。分布区域重叠的物种称同地种,分布不重叠的物种称异地种。地理亚种都是异地亚种,在交接区域常常有中间类型。
变种这一术语过去用得很杂,有的指个体变异,有的指群体类型,意义很不明确,在动物分类中已废除不用。在植物分类中,一般用以区分居群内部的不连续变体。生态型是生活在一定生境而具有一定生态特征的种内类型,常用于植物分类。人工选育的动植物种下单元称为品种。
由于种内种间变异错综复杂,分类学者对种的划分有时分歧很大。根据外部形态的异同程度作为划分物种依据而划分的称为形态种,由于对各种形态特征的重要性认识不一,使划分的种因人而异,尤其是分类学者对某些特征的“加权”常使它们比其他特征更具重要性,而造成主观偏见。古生物种或化石种中的大多数是以形态特征为依据,甚至以硬组织,如外壳、骨骼等部分形态特征为依据而建立的,应归于形态种的范畴。植物的各个部分常单独保存为化石,某些不属于同一科的生物,由于单独保存的叶、根等,很难确立其系统关系,仅以形态相似而建立的属为形态属,如栉羊齿(Pecopteris)等;其单独保存的器官可构成器官属,如鳞木类的孢子叶化石叫鳞孢叶属(Lepidostrobophyllum),孢子叶穗化石称鳞孢穗属(Lepidostrobus),而根座常被称为Stigmaria。
另外,强调染色体数目(如二倍体、多倍体、无融合生殖种等)作为分种依据而将外部形态作为次要特征依据的称为生物学种。
植物分类有大种和小种之分:“归并派”主张大种概念,又称“林奈种”,把种的范围划得较大,有时甚至很大,包括近缘种在内;“细分派”主张小种概念,又称“约尔丹种”,常把亚种、变种、生态型等划分为种,种下不再分类。动物分类学亦有归并和细分两派,但不作大种和小种的区分。归并派和细分派的存在,说明了分类学家对物种标准还缺少一致的认识。作为动物种,不同种间不能繁殖,但作为植物种则可以杂交而产生有繁殖能力的后代,如小麦。因此倾向于把二倍体作为种的代表。
这一切说明物种的概念及定义并未真正解决,同时,种上分类亦同样有归并与细分之争,例如细分派把猫科分为28个属,而极端的归并派只承认一个属。
系统原理和分类方法 一个物种或物类,以至整个植物界和动物界,都有自己的历史。研究系统发育就是探索种类之间历史渊源,以阐明亲缘关系,为分类提供理论依据。尽管在分类学派中有综合(进化)分类学、分支系统学和数值分类学3大流派,但在其基本原理上都有许多共同之处,不过各自强调不同的方面而已。系统分类的要求是:①共同起源。每一个物类都有一个起源的问题,要求同一系统内的物种或物类必须是起源于共同的祖先的,这样才能反映自然系谱。这叫做建立单系系统,单系是指一个物类起源于一个祖种。复系则指起源于两个或多个祖种。分类学要求建立单系系统,不取复系系统。②分支发展。一个新物类的产生最初总是少数,在理论上该是一个物种。从这个最初的祖种,通过分支发展,从少到多,形成为许多不同支系(图1)。
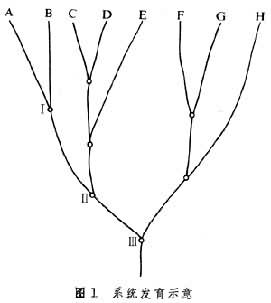 系统发展是分支发展的过程,分支关系反映亲缘关系。图1是简化的系统树,从A到H代表8个物种或物类,A和B的共同祖先在Ⅰ处,A和C的在Ⅱ处,A和F的在Ⅲ处。根据这种分支关系,W.亨尼希对亲缘关系下了一个定义:当物种A和B来源于某一个共同祖先,而这个祖先却不是物种C的祖先时,则A和B的关系要比A和C为近。因此,共同祖先的远近,决定亲缘关系的远近。他指出,所谓单系类群“是一些或大或小的类群,其群内的物种之间的相互关系比群外的任何物种为近”。例如,图1中AB、CD、FG、CDE、FGH、ABCDE都是这样的单系类群,而BCDE由于A的存在,DE由于C的存在,GH由于 F的存在,都不是单系类群。这是亨尼希的、亦即分支系统学的严格的单系概念。③级进发展(或级序发展)。分支是横的分化,级进是纵的上升,是生物从低级到高级的发展。分支与级进是相辅相成的。例如:硬骨鱼类在分支发展中有一支演进为两栖类,两栖类在分支中有一支演进为爬行类,另一支演进为哺乳类,而鸟类和哺乳类,又再各自分支发展。所以分支中有级进,级进后再分支,系统发育是分支与级进的统一发展过程(分支与级进是相对的,大分支中有大级进,小分支中有小级进)。
系统发展是分支发展的过程,分支关系反映亲缘关系。图1是简化的系统树,从A到H代表8个物种或物类,A和B的共同祖先在Ⅰ处,A和C的在Ⅱ处,A和F的在Ⅲ处。根据这种分支关系,W.亨尼希对亲缘关系下了一个定义:当物种A和B来源于某一个共同祖先,而这个祖先却不是物种C的祖先时,则A和B的关系要比A和C为近。因此,共同祖先的远近,决定亲缘关系的远近。他指出,所谓单系类群“是一些或大或小的类群,其群内的物种之间的相互关系比群外的任何物种为近”。例如,图1中AB、CD、FG、CDE、FGH、ABCDE都是这样的单系类群,而BCDE由于A的存在,DE由于C的存在,GH由于 F的存在,都不是单系类群。这是亨尼希的、亦即分支系统学的严格的单系概念。③级进发展(或级序发展)。分支是横的分化,级进是纵的上升,是生物从低级到高级的发展。分支与级进是相辅相成的。例如:硬骨鱼类在分支发展中有一支演进为两栖类,两栖类在分支中有一支演进为爬行类,另一支演进为哺乳类,而鸟类和哺乳类,又再各自分支发展。所以分支中有级进,级进后再分支,系统发育是分支与级进的统一发展过程(分支与级进是相对的,大分支中有大级进,小分支中有小级进)。假定图1上的BCDE代表爬行动物的4个支系,A是由此分出的鸟类分支,按照阶段级进的观点,鸟类和爬行类的四支比较,代表进化的一个新的阶段,必须分列为独立的一个生物群,以与其他4支相区别。这就是一般所采取的鸟纲与爬行纲并立的传统分类,也就是所谓进化分类学的分类。进化分类学派认为这个分类并不违反单系系统的原则,因为鸟类(A)作为一个单元,源出于同一祖种,而爬行类的四个支(BCDE)作为另一单元,同样是出于一个祖种。
分支学派把系统图上从同一基点所分出的支系称为姊妹群,图1上如Ⅰ处分出的A和B两支,Ⅱ处分出的AB和CDE两支,以及Ⅲ处分出的 ABCDE和FGH两支都是姊妹群。姊妹群在分类上应列为同等级别,如果图上 A代表鸟类,BCDE代表爬行类 4个支系,则A支应与B支同级,不能与BCDE四支同级。
这是当前系统分类学上的两个学派的分歧:分支系统学派以宗谱分支为衡量亲缘的唯一标准;而进化分类学派则考虑两个标准,宗谱分支和进化水平(阶段级进)。分支与级进交错发展,因而在分类工作中经常会遇到一个和两个标准的分歧,成为两个学派的争论交点。
特征对比是分类的基本方法。所谓对比是异同的对比:“异”(差异性)是区分种类的根据,“同”(相似性)是合并种类的根据。分析分类特征,首先要考虑反映共同起源的共同特征。但有同源和非同源的不同。例如鸟类的翼和兽类的前肢是同源器管,可以追溯到共同的祖先,是“同源特征”。恒温在鸟兽是各别起源,并非来自共同祖先,是“非同源特征”。系统分类采用同源特征,不取非同源性状。亨尼希把同源特征分为祖征和裔征两类:祖征是来自较早祖先而在后代保持不变的特征;裔征是两个姊妹支系来自最近祖先的派生特征,不见于较早祖先。每一物种或物类都是祖征和裔征镶嵌,共同祖征是追溯历史渊源的根据,共同裔征是建立单系的根据。进化分类学派亦要求采用同源特征,但不作特征分类。他们从共同性探索共同祖先,又从差异性衡量进化水平。上面提到鸟类和爬行类的问题,已知鸟类和鳄类最近缘,按照分支学派的分析,两者具有共同的裔征(如内鼻孔具次生腭,其他爬行类内鼻孔无次生腭),在系谱上是一对姊妹支系,组成为一个单系群。但进化分类学派重视特征差距,他们根据鸟类具有羽毛、前肢特化为翼以及恒温等特征,认为两者差距很大,反映不同的进化阶段,主张鸟类应与鳄目分开,而与爬行类(包括鳄目)并立为纲。这是两个学派对特征衡量的分歧,也就是宗谱分支与进化水平的分歧。
分类特征反映进化历史,系统分类要求从历史观点衡量特征价值。但特征衡量常会出现分歧,重视这一或那一特征,会得出不同的分类。在某些情况下,由于同源和非同源不易区分,衡量又会陷入循环论,如:共同的特征反映共同祖先,共同的祖先产生共同特征。基于这些情况,为了促进分类的稳定性和重复性,数量分类学主张“总体相似性”的观点,其方法是:采用大量能以数值表示的特征,编成符号,运用电子计算机。求得相似性系数,作出分类。数量分类不先考虑亲缘,但得出的结果在很多情况下和传统分类近似。
生命现象的各个方面都可为分类提供特征,最常用的是形态,尤其是外部形态。但目前趋向愈来愈重视生理、生化、遗传等方面的特征,如DNA(脱氧核糖核酸)的含量比较,蛋白质的成分分析,染色体的祖型分析,以及植物化学成分、动物交配行为等等。如以蟋蟀鸣声为特征,使原先单凭形态特征所未能区分的近缘种类得以鉴别。
生物的界级分类 林奈把生物分为两大类群:固着的植物和行动的动物。两百多年来,随着科学的发展,人们逐渐发现,这个两界系统存在着不少问题,但直到20世纪50年代,仍为一般教本所遵从,基本没有变动。
最初的问题产生于中间类型,如眼虫Euglena综合了动植物两界的双重特征,既有叶绿体而营光合作用,又能行动而摄取食物。植物学者把它们列为藻类,称为裸藻;动物学者把它们列为原生动物,称为眼虫。中间类型是进化的证据,却成为分类的难题。为了解决这个难题,早在19世纪60年代,人们建议成立一个由低等生物所组成的第三界,取名为原生生物界(Protista),包括细菌、藻类、真菌和原生动物。这个三界系统解决了动植物界限难分的问题,但未被接受,整整100年后,直到20世纪60年代,才开始流行了一段时间,为不少教科书所采用。
生命的历史经历了几个重要阶段,最初的生命应是非细胞形态的生命,当然,在细胞出现之前,必须有个“非细胞”或“前细胞”的阶段。病毒就是一类非细胞生物,只是关于它们的来历,是原始类型,还是次生类型,仍未定论。从非细胞到细胞是生物发展的第二个重要阶段。早期的细胞是原核细胞,早期的生物称为原核生物(细菌、蓝藻)。原核细胞构造简单,没有核膜,没有复杂的细胞器。从原核到真核是生物发展的第三个重要阶段。真核细胞具有核膜,整个细胞分化为细胞核和细胞质两个部分:细胞核内具有复杂的染色体装置,成为遗传中心;细胞质内具有复杂的细胞器结构,成为代谢中心。由核质分化的真核细胞,其机体水平远远高出于原核细胞。从单细胞真核生物到多细胞生物是生命史上的第四个重要阶段。随着多细胞体形的出现,发展了复杂的组织结构和器官系统,最后产生了高级的被子植物和哺乳动物。
植物、菌类和动物组成为生态系统的 3个环节。绿色植物(真核植物和原核蓝藻)是自养生物,是自然界的生产者。它们通过中绿素进行光合作用,把无机物质合成有机养料,供应自己,又供应异养生物。菌类(细菌和真菌)是异养生物(自养细菌除外),是自然界的分解者。它们从植物得到食料,又把有机食料分解为无机物质,反过来为植物供应生产原料。动物亦是异养生物,它们是消费者,是地球上最后出现的一类生物。即使没有动物,植物和菌类仍然可以存在,因为它们已经具备了自然界物质循环的两个基本环节,完成循环过程中合成与分解的统一。但是,没有动物,生物界不可能这样丰富多彩,更不可能产生人类。植物、菌类和动物代表生物进化的3条路线或3大方向。
以上是目前对生物进化的阶段和方向的认识,是现代界级分类的根据。当前最流行的分类是一种五界系统(表2)。

五界系统反映了生物进化的 3个阶段和多细胞阶段的 3个分支,是有纵有横的分类。它没有包括非细胞形态的病毒在内,也许是因为病毒系统地位不明之故。它的原生生物界内容庞杂,包括全部原生动物和红藻、褐藻、绿藻以外的其他真核藻类,包括了不同的动物和植物。不少学者认为,不必成立原生生物界,而把藻类和原生动物分别划归植物界和动物界,成为比较紧凑的四界系统。反之,亦还有不少人主张扩大原生生物界,把真菌划归在内,成为另一种四界系统。
本卷采用六界系统 (表2)。因为任何生物系统都应把病毒(连同类病毒)包括在内。病毒是非细胞生物,但不一定代表生命的前细胞阶段。六界分类系统,在病毒界和细菌界之间还加有问号;因为病毒如果是原始生物,细菌作为早期的原核生物,很可能起源于“原病毒”。原核生物的最早化石记录,包括细菌和蓝藻,见于30多亿年前的地层中;真核生物的最早记录,包括金藻和绿藻,见于14~15亿年前的地层中。
自养的生产者蓝藻和异养的分解者细菌组成为原核时代的两大生态环节,正如植物、真菌和动物组成为真核生物的三大生态环节。这五类生物,基于它们的重要时空地位,都应独立为界,连同病毒,综合为六界分类系统(图2)。
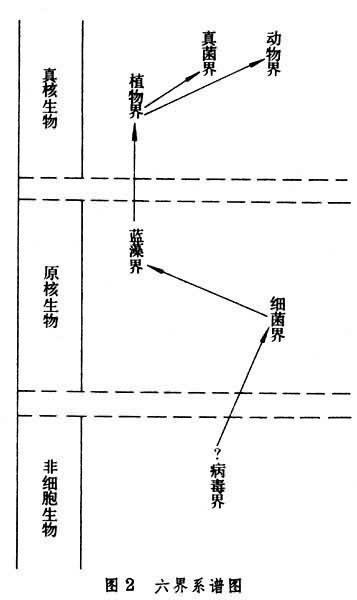
蓝藻显然起源于细菌,是细菌的一支;真核植物大概起源于原核蓝藻,是蓝藻的一支(见细胞起源)。从真核植物又分支发展为真菌和动物 (图2)。一般说来,植物细胞有细胞壁和叶绿体,真菌细胞有壁无叶绿体,动物细胞无壁无叶绿体。如果把原核的蓝藻和动植物的中间类型裸藻都归在植物界内,列为蓝藻门和裸藻门。真菌和动物的中间类型粘菌,以及真菌和藻类共生的地衣,归在真菌界内,则可成为一种四界系统。