核型(卷名:生物学)
karyotype
动物、植物、真菌等真核生物的某一个体或某一分类群(亚种、种、属等)的细胞内具有的相对恒定特征的单倍或双倍染色体组 (见彩图)。染色体的特征以有丝分裂中期最为显著,包括染色体的数目、长度、着丝粒的位置、随体(指某些染色体末端的球形小体,由着色浅而狭细的副缢痕与染色体臂相连)与副缢痕的数目、大小、位置以及异染色质和常染色质在染色体上的分布等。
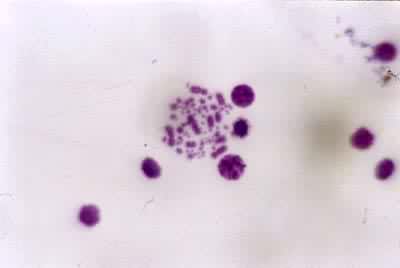
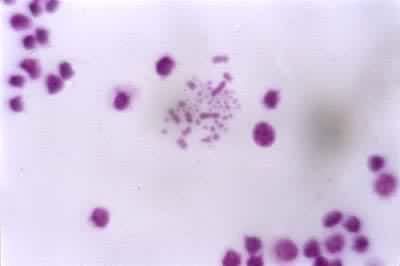
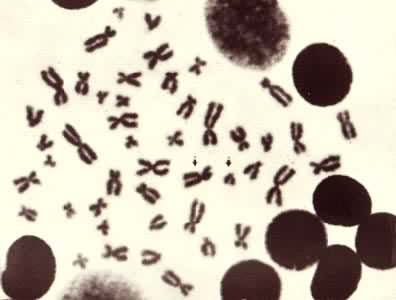
将一个染色体组的全部染色体逐条按其特征画下来,再按长短、形态等特征排列起来的图称为核型模式图,它代表一个物种的核型模式。
由于许多物种的各个染色体靠普通的制片染色方法不易精确地识别和区分,1968年以来发展起来的显带技术,即用各种特殊的处理和染色方法使各条染色体显示出各自的横纹特征(带型)的方法成为研究核型的有力工具。
核型及其各种带型是动物、植物、真菌在染色体水平上的表型。研究和比较各种动物、植物、真菌的核型和带型有助于对各个种、属、科的亲缘关系作出判断,揭示核型的进化过程和机制。此外,核型的研究又和人类自身利害密切相关,它的数目和结构的改变往往给人类带来遗传性疾病──染色体病;肿瘤细胞的核型分析已被应用于肿瘤的临床诊断、预后及药物疗效的观察;通过培养后的淋巴细胞或皮肤成纤维细胞的核型分析,可以对人的染色体病进行诊断,而对培养后的羊水中的胎儿脱屑细胞或胎盘绒毛膜细胞的核型分析则可用于对胎儿的性别和染色体病的产前诊断。
历史 核型一词首先由苏联学者T.A.列维茨基和JI.杰洛涅等在20世纪20年代提出。1952年美国细胞学家徐道觉首先采用低渗处理技术使细胞内的染色体分散而便于观察,以后秋水仙素的应用使增殖中的细胞停止于中期,从而便于获得大量供观察的中期分裂相,植物凝血素(简称 PHA)刺激白细胞分裂的发现使以血培养方法观察动物与人的染色体成为可能。随着各种培养、制片、染色技术的改进使核型的研究进入了蓬勃发展的新阶段。1956年瑞典细胞遗传学家庄有兴等报告了人的染色体数是46而不是过去认为的48。1959年以后在人类中发现越来越多的各种各样的染色体异常。1960年 4月在美国丹佛市召开的国际学术会议上对人的染色体分群和命名的术语、符号、方法等作了统一规定,在第五次国际人类遗传学会议上产生的人类染色体命名常务委员会又于1977年专门召开了会议进行修订,会后公布了《人类细胞遗传学命名国际体制(ISCN)(1978)》。1981年该委员会又公布了《人类细胞遗传学高分辨显带命名国际体制》,在1977年所制订的中期染色体带型命名规定的基础上提出了高分辨的晚前期和早中期染色体带型命名规定和模式图。这些规定目前为世界各国学者所普遍采用。
方法 核型研究所用的材料或是自然条件下活体中正在旺盛分裂的细胞(如植物的根尖、嫩叶、茎尖等细胞,以及动物的胚胎细胞、骨髓细胞、睾丸中的精原细胞等)或是离体培养的旺盛分裂的细胞。植物细胞一般不经低渗处理,如需经低渗处理则需用酶溶去细胞壁。动物细胞则往往经低渗处理后再行固定、染色。
常用的显带技术所显示的带有Q带、G带、C带、R带、T带等。Q带技术即喹吖因荧光染色技术,是1968年瑞典细胞化学家T.O.卡斯珀松建立的,所显示的是中期染色体经氮芥喹吖因或双盐酸喹吖因染色后在紫外线照射下所呈现的明亮的荧光带,这些区带相当于DNA分子中A:T碱基对成分丰富的部分。G带即吉姆萨带,是将处于分裂中期的细胞经过胰酶或碱、热、尿素、去污剂等处理后再经吉姆萨染料染色后所呈现的区带。C 带又称着丝粒异染色质带,是着丝粒邻近的异染色质部分。C带技术是M.L.帕多等于1970年建立的。R带由 B.迪特里约于1971年所首创,是中期染色体不经盐酸水解或胰酶处理,只在磷酸缓冲液中保温处理后就用吉姆萨等染料染色后所呈现的区带,也是G带染色后的带间不着色区,所以又称反带。T带又称端粒带,是染色体的端粒部位经吖啶橙染色后所呈现的区带,典型的T带呈绿色,由B.迪特里约1973年首先报道。染色体银染法系用硝酸银 (AgNO3)使染色体上的核仁形成区部位呈现黑色的一种特殊染色法。1975年以来,美国细胞遗传学家J.J.尤尼斯等又建立了高分辨显带法,方法是先用氨甲喋呤使细胞分裂同步化,然后用秋水酰胺进行短时间的处理,使出现大量晚前期和早中期的分裂相。在这样处理过的人的早中期细胞的染色体组中可以看到555~842条带。在晚前期细胞中可以看到843~1256条带,而已往在中期染色体上只能观察到320~554条带。后来又用放线菌素D作用于DNA合成后期(G2期)的细胞以阻碍染色体浓缩时特殊蛋白质与染色体的结合,从而使染色体更为细长,使所显示的带纹多达5000条。这样就可以更精确地观察染色体的各种变异,甚至在各种生物的正常个体细胞中也可以看到染色体的各种区带的宽窄、位置等存在着一些变化,这些变化称带的多态现象。
人类染色体命名符号与核型式 《人类细胞遗传学命名国际体制(ISCN)(1978)》中的命名符号:
A、B、C、D、E、F、G染色体群的符号
1~22 常染色体的顺序号
X、Y 性染色体
/ 嵌合体或异源嵌合体的不同细胞系之间的分隔
符号
+ 增加
- 丢失
→ 从→到
: 断裂
∷ 断裂和重接
( ) ( )内为结构改变的染色体
? 染色体或其结构的鉴别有疑问
; 在涉及两个以上染色体的结构重排中用来分隔
染色体或染色体区段的符号
= 总数为
(
 ) 用于区别同源染色体
) 用于区别同源染色体* 作为×号用,其前是母本,其后是父本。
AⅠ(first meiotic anaphase) 减数分裂Ⅰ后期
AⅡ(second meiotic anaphase) 减数分裂Ⅱ后期
ace(acentric fragment) 无着丝粒断片
b(break) 断裂
cen(centromere) 着丝粒
chi(chimera) 异源嵌合体
cs(chromosome) 染色体
ct(chromatid) 染色单体
cx(complex) (染色体)群
del(deletion) 缺失
der(derivative chromosome) 衍生染色体
dia(diakinesis) 浓缩期
dic(dicentric) 双着丝粒
dip(diplotene) 双线(期)
dir(direct) 直接(分裂)
dis(distal) 远端
dit(dictyotene) 核网(期)
dmin(double minute) 双微小体
dup(duplication) 重复
e(exchange) 互换
end(endoreduplication) 内复制
f(fragment) 断片
fem(female) 女性
g(gap) 裂隙
h(secondary constriction) 副缢痕(或次缢痕)
i(isochromosome) 等臂染色体
ins(insertion) 插入
inv(inversion) 倒位
lep(leptotene)细线(期)
MⅠ(first meiotic metaphase) 减数分裂Ⅰ中期
MⅡ(second meiotic metaphase) 减数分裂Ⅱ中期
mal(male) 男性
mar(marker chromosome) 标记染色体
mat(maternal origin) 来自母亲
med(median) 中央
min(minute) 微小体
mn(modal number) 众数
mos(mosaic) 嵌合体
oom(oogonial metaphase) 卵原细胞中期
p(short arm of chromosome) 染色体短臂
PⅠ(first meiotic prophase) 减数分裂Ⅰ前期
pac(pachytene) 粗线(期)
pat(paternal origin) 来自父亲
pcc(premature chromosome condensation) 染色
体提前浓缩
Ph(Philadelphia chromosome) 费城染色体
prx(proximal) 近端
psu(pseudo) 假
prz(pulverization) 粉碎
q(long arm of chromosome)染色体长臂
qr(qua lriradial) 四射体
r(ring chromosome) 环状染色体
rcp(reciprocal) 相互(易位)
rea(rearrangement) 重排
rec(recombinant chromosome) 重组染色体
rob(Robertsonian translocation) 罗伯逊易位
s(satellite) 随体
SCE(sister chromatid exchange) 姐妹染色单体
互换
sdl(sub-line,side line) 亚系、旁系
sl(stem line) 干系
spm(spermatogonial metaphase) 精原细胞中期
t(translocation) 易位
tan(tandem translocation) 衔接易位
ter(terminal,end of chromosome) 末端、染色体
端部
tr(triradial) 三射体
tri(tricentric) 三着丝粒
var(variable chromosome region) 染色体可变区
xma(ta)[chiasma(ta)] 交叉
zyg(zygotene) 偶线(期)
在用核型式描述一个核型时,第一项是染色体总数(包括性染色体),然后是一个逗号,最后是性染色体。下面是一些核型式的举例:
46,XX 正常女性;
46,XY 正常男性;
45,X 特纳氏综合征(Turner's syndrome);
47,XXY 克氏综合征(Klinefelter's syndrome);
47,XY,+21 男性21三体;
46,XY,1q+ 具46条染色体的男性,第1号染色体长臂
延长;
chi46,XX/46,XY具有XX和XY细胞系的异源嵌合体;
46,XX,t(Xq+;16q-) 具46条染色体的女性,X染色体
长臂与第16号染色体短臂之间相互易位(X染色体
长臂延长,16号染色体短臂缺失);
46,XX,del(1) (pter→q21∷q31→qter) 具有46条染
色体的女性,1号染色体长臂上1q21和1q31带间断裂
和重接,这些带间的片段缺失。
应用 医学诊断 不少恶性肿瘤的核型中常出现不规则的非整倍体、多倍体或标记染色体。例如在绝大多数慢性粒细胞性白血病人的骨髓细胞中都可以发现有一个小的特殊染色体。这一标记染色体称为Ph1染色体,是一个22号染色体的长臂第一区第一带以远末端部分缺失的结果,22号染色体长臂丢失的部分易位到9号染色体长臂上第三区第四带的末端。又如视网膜母细胞瘤患者的核型具有13号染色体的长臂部分缺失特征(见染色体畸变、肿瘤遗传学)。由于高分辨显带技术的应用,新近又发现几十种常染色体综合征与染色体臂的部分缺失或重复有关。
遗传学研究 核型分析广泛应用于动植物染色体倍性、数目和结构变异的分析和染色体来源的鉴定,通过细胞融合所得来的杂种细胞的研究以及基因定位研究中单个染色体的识别等方面。在动植物分类和生物进化研究中也得到广泛的应用。
对越来越多的动、植物物种所进行的核型及带型分析,使原来以形态学和解剖学指标为依据的分类学提高到了一个新的水平,并不断地丰富了对染色体进化规律与机制的了解。例如灵长类动物核型的研究表明黑猩猩及大猩猩的核型和人的十分相似,如果它们的两对端着丝粒染色体融合在一起成为一个,就相当于人的2号染色体,此外的区别是人的4、5、9、12、17、18号染色体中各有一段倒位。
灵长类染色体进化的主要机制是染色体在着丝粒处的融合,其次是倒位与衔接等。鹿亚科核型的研究则表明其进化机制主要是一条中部或亚中部着丝粒染色体在其着丝粒部位断裂成为两条端着丝粒染色体。