胚胎发育(无脊椎动物)(卷名:生物学)
embryonic development of invertebrate
━━━━━━━━━━━━━━━━━━━━━━━━
基本模式
原生动物模式
两胚层动物模式
无体腔动物模式
假体腔动物模式
裂腔动物模式
肠腔动物模式
发育类型
自由幼虫发育类型
直接发育类型
非自由幼虫发育类型
发育方式
有性方式的发育
无性方式的发育
再生━━━━━━━━━━━━━━━━━━━━━━━━
胚胎发育一词通常是指从受精卵起到胚胎出离卵膜的一段过程。而无脊椎动物胚胎学家则常把其概念扩展到胎后发育直到性成熟,甚至整个生活史。
无脊椎动物包括30多个门类,在种数上约为脊索动物的22倍,几乎存在于地球的各种环境中。由于不同种动物具有不同发展历史、不同形态和不同繁殖方式等,它们的胚胎发育表现了高度的多样性和复杂性。在这种情形下,不同门类无脊椎动物胚胎发育之间所显示的各种联系都需从不同的角度予以归纳和分析。
众所周知,无脊椎动物胚胎发育表现出的首要特点是其过程并不限在卵膜或母体内进行,在很多种类中有幼虫期。由幼虫到成体要经过变态。用形态发生的程度来衡量,无脊椎动物幼虫期仍属胚胎期。胚胎学者称不经幼虫期的发育为直接发育,经过幼虫期的为间接发育。
绝大多数无脊椎动物的胚胎发育是在有性生殖基础上进行的,有少数种类兼行孤雌生殖式的胚胎发育。腹毛虫纲中的大多数鼬虫属动物因缺少雄性生殖器官而只进行后一种胚胎发育。此外,有些种类的无性生殖和再生都与胚胎发育相似。
无脊椎动物胚胎发育除普遍经过卵裂、囊胚、原肠胚、幼虫和成体器官发生等阶段外,有的在幼虫期之后和未进入成体之前还有一个称为后幼虫期的过渡阶段,如对虾类的后幼虫期已具所有附肢,但在体躯比例、附肢长短和外部生殖器官等方面尚未达到成体水平。又如蟹的后幼虫期(大眼幼虫)具有与成体相同的头胸部,但宽大的腹部尚未弯折其下。
由于进化过程的不同,无脊椎动物胚胎期较脊椎动物为短,但各发育阶段明显。同理,在各门无脊椎动物内部,凡进化水平越低的种类,其胚胎发育历程越短,各个发育阶段越明显;幼虫的出现越早,历时相对越长,是以对系统发育各阶段的反映越清楚。
无脊椎动物的胚胎发育,与脊椎动物相同处在:也是按阶段循序进行生物合成和细胞分化;由组织发生到器官发生,终而在形态建立的基础上出现功能分化。以上过程,不仅要具体通过细胞间和组织间的相互作用,而且还必须具备一定的环境条件才能实现,如光照、干旱和寒冷对胚胎或幼虫的蜕皮和滞育有直接影响,底质的理化和生物因素可决定水生幼虫的沉落、附着和变态,而营养、盐度、温度、光照、空间和外激素等条件都可影响性腺的发生和分化。
基本模式
直接发育和间接发育,作为两种模式,虽能够概括所有多细胞动物的胚胎发育,但还不足以表达出无脊椎动物各门类胚胎发育的特点及其相互联系。各门无脊椎动物的后期胚胎发育或变态期的形态发生过程,都各具发展方向,当然难以归纳为少数模式。但如以早期胚胎发育过程为准,则无论在进化水平相近门类之间或不同进化水平门类之间,都存在着一些形态发生上的共性或规律性的内容。这方面内容不仅体现着个体发育与系统发育间相互制约关系,体现着进化地位的高低,也体现着亲缘关系的远近。20世纪40年代以来,以上内容已成为确定各门无脊椎动物分类位置和说明其进化方向的重要依据,因而这里也以之试作建立无脊椎动物胚胎发育基本模式的理论基础。下述几种胚胎发育基本模式除原生动物模式和两胚层动物模式外,其他几种模式与分类上无体腔动物、假体腔动物、裂腔动物和腔肠动物的划分是一致的。
原生动物模式 原生动物体制结构简单,传代迅速。通常仅用“生殖”一词概括其个体发育。其实这类动物的生殖过程也有繁简不同。单裂生殖最简单,而有性生殖和世代交替都较复杂。后两者的整个过程常称为生活史或发育环。重要的是在发育环中也有过渡形态出现,致使原生动物与多细胞动物的个体发育有了共同点。
原生动物的无性生殖通过个体直接分裂来完成。包括二裂和复裂两种基本形式。例如红球虫的复裂(图1),
 开始时先失去鞭毛并在细胞表面分泌出胞壳,壳内虫体则变为孢母细胞。此细胞经两次分裂成为 4个子细胞。后者在生出鞭毛后即称为游走孢子。由此孢子长成新个体。孢母细胞和后来子细胞因形态上不同于亲体而相当于多细胞动物个体发育中的胚胎期,由游走孢子到成体相当于胚后生长期。
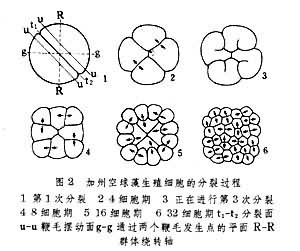
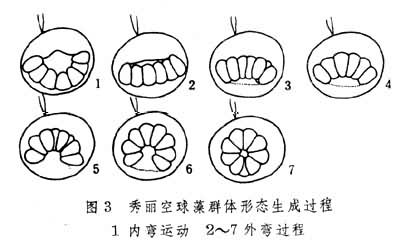
开始时先失去鞭毛并在细胞表面分泌出胞壳,壳内虫体则变为孢母细胞。此细胞经两次分裂成为 4个子细胞。后者在生出鞭毛后即称为游走孢子。由此孢子长成新个体。孢母细胞和后来子细胞因形态上不同于亲体而相当于多细胞动物个体发育中的胚胎期,由游走孢子到成体相当于胚后生长期。有一种群体鞭毛虫(真球藻)的无性分裂生殖比红球虫有所演进。首先,其复裂开始于无性生殖母细胞而非始自一般营养个员。肥大的生殖母细胞在发生中经过生长阶段,很象有性生殖中的卵子。其后,复裂即以类似腔肠动物中表格式卵裂方式在两度空间内进行(图2)。结果形成板状细胞层。该细胞层经弯卷后形成新的球形的群体(图3)。此球状群体本身堪与多细胞动物的鞭毛式囊胚相比拟,这意味着原生动物发育环中所出现的最高形态仅能达到多细胞动物囊胚期的水平。
在原生动物发育模式方面有两点最值得注意:①如前所述,其过程中有过渡形态出现,是可作为有胚胎发育期的证据;②从生殖母细胞的出现到精、卵的成熟与结合表明。这类动物的性表现基本上已接近多细胞动物的水平。
两胚层动物模式 多孔动物和腔肠动物属此模式,其特点是发育过程中只形成原始外胚层和原始内胚层而无中胚层,于是所有成体结构都来自这两个胚层。再则,由于被列为侧生动物的多孔动物与获有辐射动物之称的腔肠动物采取不同进化方向,所以两者的胚胎发育必然还有次要内容的差异。
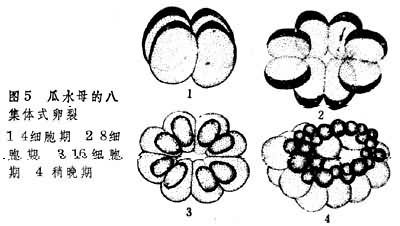
就卵裂来说,等裂和不等裂皆出现于多孔动物中,但有更多类型,如不规则式、辐射式和具有螺旋性质的卵裂,皆见于腔肠动物中的有刺泡类中(图4)。8集体式定型卵裂(图5)仅为栉水母类动物所有。
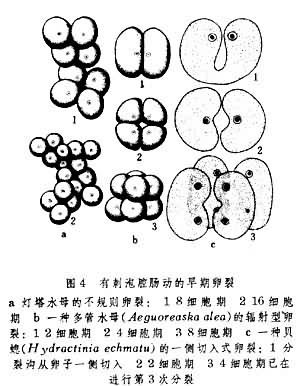
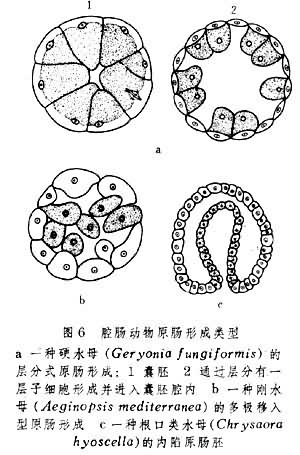
关于原肠形成,多孔动物和腔肠动物同有单一的移入式、层分式与内陷式(图6)。这在演化上属较低级形式。唯有在栉水母类中才有内陷与外包相结合的情形。
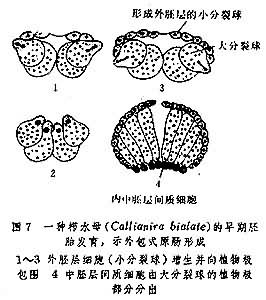
对两胚层动物来说,介于内外两胚层之间的基质被统称为间充质,多孔动物间充质中的胶细胞和有刺泡类动物中的间隙细胞都来源于外胚层,故被称为外中胚层细胞,栉水母胶细胞因来源于内胚层而被称为内中胚层细胞(图7)。
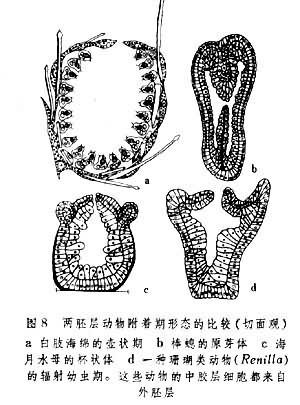
两胚层动物胚胎发育在进入原肠胚期的同时,也多半是幼虫期的开始,但也有早自囊胚阶段即已进入幼虫期的。幼虫经过一定时间和一定形态发生过程后即进行附着。刚附着的幼体呈杯状或筒状,在顶端有开孔,内部有腔隙,围壁由内外两胚层共同组成。海绵动物的此一发育期称壶状期,水螅虫者称原芽体,真水母者称杯状体,珊瑚虫者称辐射幼虫(图8)。由这一阶段再分别发育为上述各类动物的新个体。
无体腔动物模式 扁形动物和纽形动物在胚胎发育期间都有过原始体腔──囊胚腔的形成,只是由于间叶细胞在腔内的增加和分化,至成体时此腔才又失去。这是该两类动物胚胎发育同归于无体腔动物模式的唯一基础。
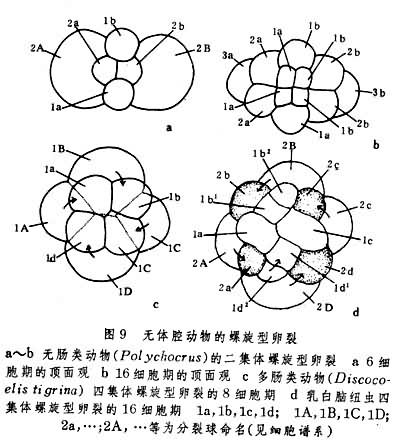
由于进化道路和环境需要的不相同,上述两门动物胚胎发育之间以及扁形动物内各纲胚胎发育之间都有很多差异。首先,在卵裂方面,纽形动物都取螺旋型(图9);而在扁形动物中,固然许多海生无肠类、多肠类和由内黄卵发育的单肠类皆取螺旋型,但由外黄卵发育的三肠类和吸虫类却取不规则形式(图10)。在绦虫类中,经过等全裂或不等全裂而生成的分裂球并不全部参加胚体形成,其中只有位于中央部分的小裂球分化为胚胎细胞,居于表面的大裂球则形成胎膜。
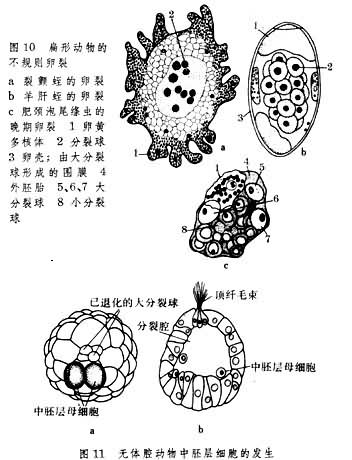
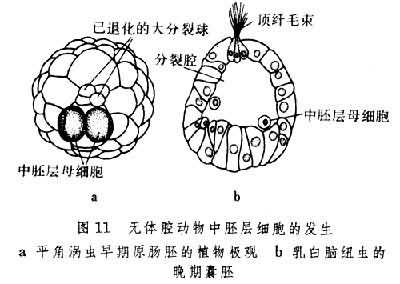
其次,无体腔动物的胚层形成与分化情况也很复杂。在无肠类、多肠类和纽形动物中都有中胚层母细胞的出现(图11),但在某些单肠类和三肠类动物中,卵裂后形成的细胞团只分化为内胚层和外胚层。它们的中胚层细胞到胚胎后期才能出现。吸虫类和绦虫类也属此种情形。
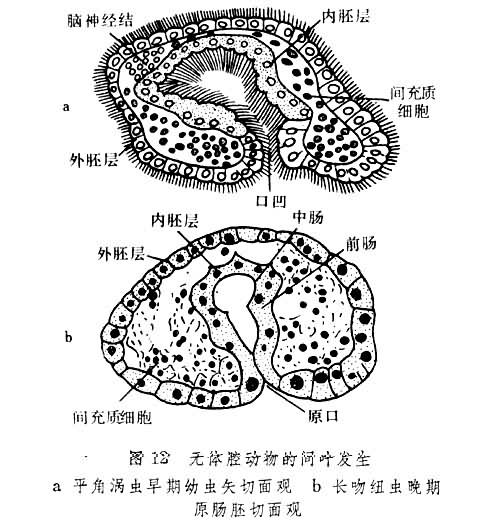
无体腔动物的中胚层细胞出生后总是以间叶细胞的形态不断增多(图12)。其中有的长时期保持固有形态,有的变成肌肉纤维并分布于肠壁(内胚层)和体壁(外胚层)之间。
无体腔动物的后期胚胎发育,在大多数种类中仍属幼虫期发育。其中少数是自由生活幼虫(多肠类动物),多数是寄生幼虫(吸蛭、绦虫)。它们的变态过程各有独特表现。
假体腔动物模式 轮形动物(Rotifera)、线虫(Ne-matoda)、线形动物(Nematomorpha)、腹毛虫 (Gastro-tricha)、棘头虫(Acanthocephala)和内肛动物 (Ento-procta) 在胚胎发育过程中都把囊胚腔保留下来形成假体腔。这是本模式的基本特点。
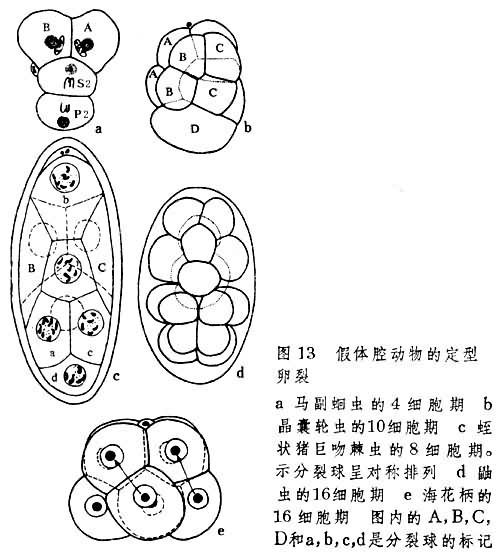
各门假体腔动物的卵裂不但普遍严格地按一定形式进行定型卵裂,并且各有独自特点(图13)。其中轮虫和棘头虫的早期卵裂具螺旋型性质。
假体腔动物的原肠形成多取内陷式,但是,线形动物和内肛动物中的内陷是指囊胚的植物极细胞层同时向动物极一侧的陷入,而轮虫、线虫和腹毛虫的内陷过程却皆从少数分裂球进入囊胚腔开始(图14c、d)。
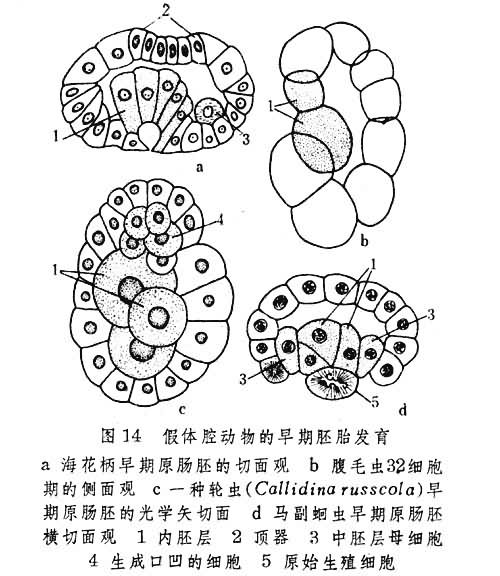
中胚层的发生,大致有3种情况:
①在早期胚胎发育期间无中胚层母细胞或细胞团的出现,如轮虫和腹毛虫(图14b、c)。棘头虫虽在此时期有内细胞团生成,但其组成细胞尚无中胚层和内胚层之分。
②在原肠形成期间,先有部分外胚层细胞进入囊胚腔,形成外中胚层间叶细胞;继之,再在原口附近生出内中胚层细胞,如线形动物中的铁线虫。
③出现中胚层母细胞。它们通过持续分裂,形成胚体左右两侧的中胚层带,如内肛动物中的海花柄(图14a)和线虫类中的马副蛔虫(图15)。
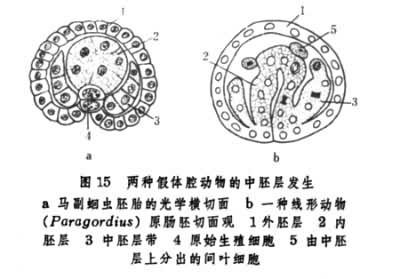
多数假体腔动物的中胚层细胞主要分化为肌肉纤维,只有轮虫是由外胚层细胞在胚胎晚期分化成肌纤维细胞。这种肌纤维分布在体壁外胚层内表面的情形不仅使体壁的组成得到加固,同时也使假体腔本身具有了处于肠壁内胚层和体壁中胚层之间的特点。
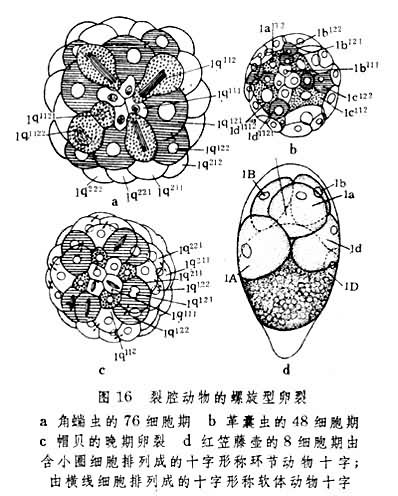
裂腔动物模式 外肛动物(Ectoprocta)、帚虫(Pho-ronida)、星虫(Sipunculoidea)、螠虫(Echiuroidea)、环节动物(Annelida)、软体动物(Mollusca)和节肢动物(Arthropoda)的胚胎发育同属此模式。它们的体腔都以裂隙形式出现于中胚层带或中胚层细胞团内。
除均有裂生体腔外,裂腔动物模式的特点还表现在卵裂类型、中胚层分化形式和幼虫形态等方面。此模式中各类动物彼此间所显示的进化关系较其他模式中所显示的要明显得多。
典型螺旋卵裂普遍出现于大多数裂腔动物门类中。环节动物和软体动物卵裂的相似甚至表现在分裂球的排列形式上(图16a、c)。节肢动物的卵裂虽在向着表面卵裂方向发展(图17b),但在其低等种类中仍有螺旋性质出现。头足类中的盘状卵裂和外肛动物中的辐射型卵裂在本模式中均属少见。

关于原始胚层的生成,不同门类间颇有差异,此表现在原肠形成的方式有取单一形式的内陷、外包、分层或移入的,如卤虫限于移入,藻苔虫(Flustrellidra)限于分层;也有结合数种形式的,如头足类(图18)。
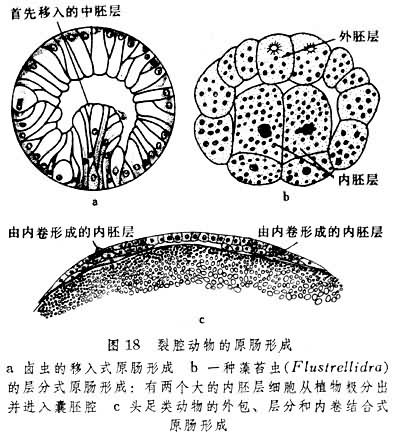
随着原肠的形成,中胚层的来源及其的产生方式在各类动物中有所不同。星虫、外肛动物的裸唇类、帚虫和螠虫的都来自内胚层,而环节动物、节肢动物和软体动物的一部分中胚层来自外胚层,其主要部分来自内胚层。前者称外中胚层,后者称内中胚层。裂腔动物中不少门类的内中胚层起源于一个中胚层母细胞(图19),但有个别门类(帚虫)起源于多个细胞向囊胚腔中的陷入。
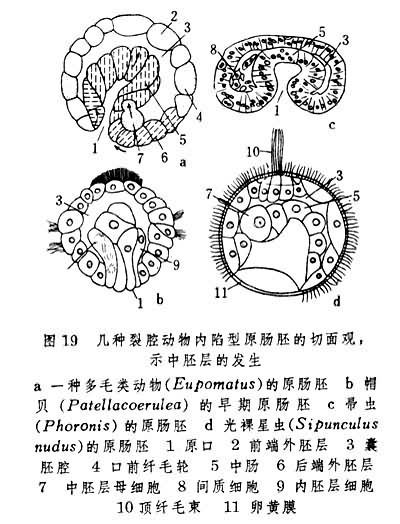
在发育过程中,外中胚层主要分化为幼虫肌肉,内中胚层发展为间叶细胞群或细胞纵带(图20)。帚虫的体腔即出现在间叶细胞群内;在外肛动物中由间叶细胞组成体腔膜。
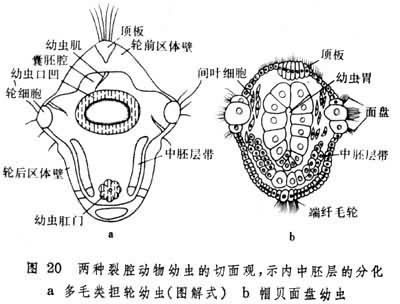
至于细胞带的发展,情况很不一致。星虫的中胚层带不经分节即分化为体腔和肌肉,而环节动物者却是随着胚体的生长,自前向后进行分节并在各节中生出腔隙,此一系列体节和体腔保留到成体并得到发展。在节肢动物中成对的体节及所含腔隙仅仅是过渡形态,不久即分别形成肌肉和血腔。
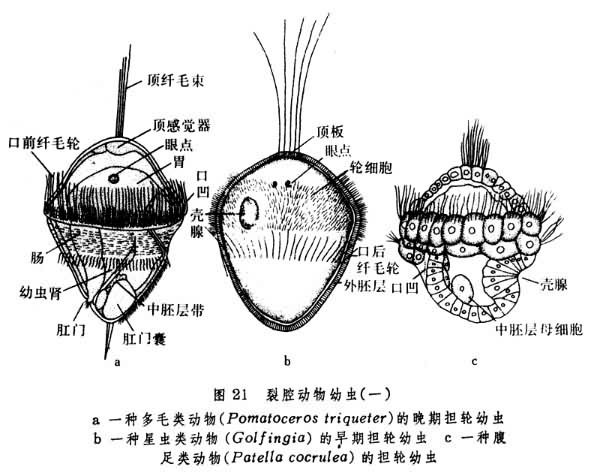
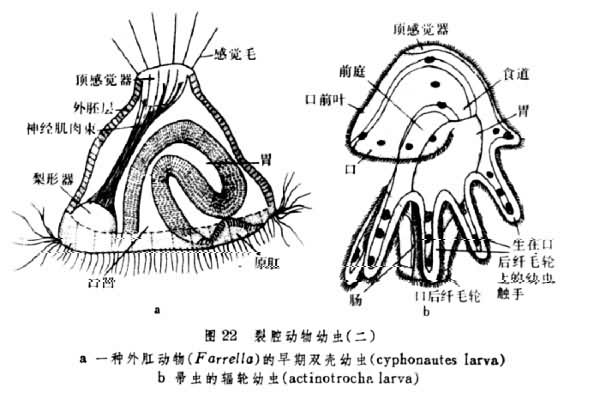
经过胚层的形成和初步分化,裂腔动物各门的发育普遍进入幼虫期。星虫、螠虫、环节动物和软体动物都有形态大致相同的担轮幼虫期(图21)。外肛动物中的后湾幼虫和帚虫中的辐轮幼虫,都与担轮幼虫有一定的相似(图22)。这些幼虫的口都在原口基础上生成,然后在变态过程中再变为成体口孔。这一情形与在无体腔动物和假体腔动物发育中所见一致。
肠腔动物模式 腕足动物(Brachipoda)、棘皮动物(Echinodermata)和毛颚动物 (Chaetognatha)都由原肠生出体腔(图23),这是肠腔动物模式的主要特点。此外,辐射型卵裂、内陷式原肠形成(图24)和体腔分节(图25)在这类动物胚胎发育中也普遍出现。
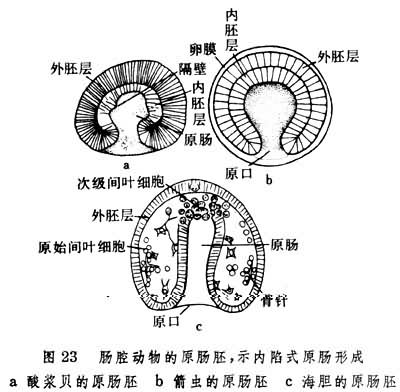
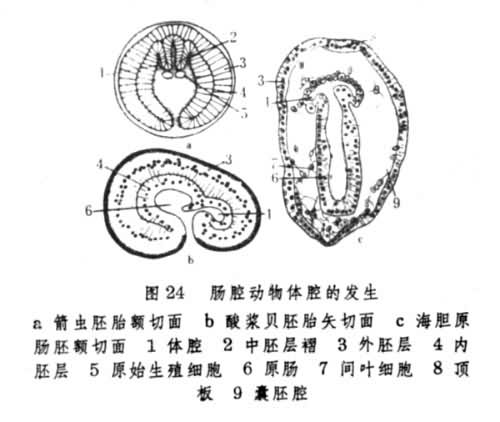
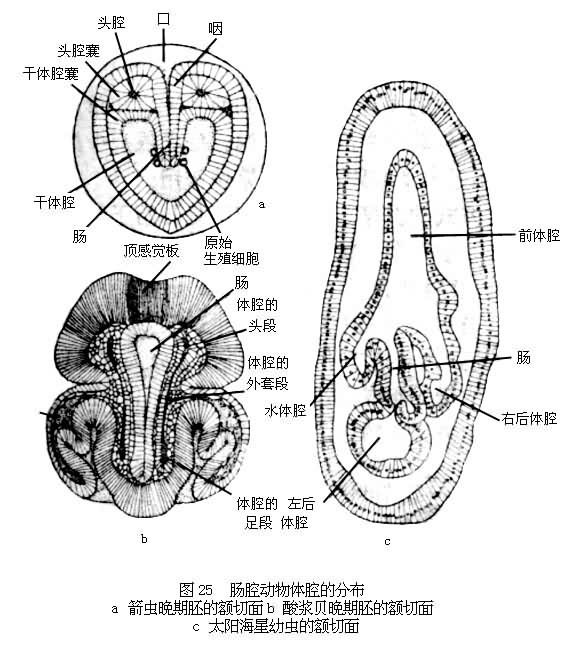
棘皮动物和毛颚动物的体腔在发育过程中分为前后两部分,某些腕足类动物如酸浆贝则分前中后3部分。各部分体腔的进一步发展在不同门类间有相当差异。毛颚动物的干腔和尾腔分别由中、后两部分体腔形成,其头腔完全分化为头部肌肉。腕足动物的体腔有分枝通入触手冠和外套膜中。棘皮动物的左前体腔演变为水管系统。只有其左后体腔才是成体体腔的发生基础。
在肠腔动物中,腕足类和棘皮动物的后期胚胎发育仍表现为幼虫阶段。棘皮动物成体为辐射对称体制,但其幼虫期都为左右对称体制。棘皮动物变态的复杂化与此有直接联系。
上述6种模式中,除原生动物模式外,其余5种模式都为多个门类所共属。各模式之间不仅有内容上的不同,而且还有进化水平上的差别。诸如由无胚层到有胚层,由两胚层到三胚层,由无体腔到有体腔,由假体腔到真体腔,以及由裂生体腔到肠生体腔等这一系列由简单到复杂、由低级到高级的演化历程都在无脊椎动物胚胎发育中有充分显示。
发育类型
各类无脊椎动物在其历史发展基础上,不断提高适应能力,克服种种环境影响以求得个体生存和种族的延续。正是在此长期进化过程中,它们逐渐各自形成了独有形式的生活环(动物)。
动物生活环包括多个阶段,其中最敏感最脆弱的当是胚胎发育阶段。为顺利渡过这一阶段需要解决多方面困难,其中主要是营养物质来源。概括说来,无脊椎动物胚胎发育期的营养有3种来源:①由胚体或幼虫自己取得;②由母体先为之储备于卵内;③由母体直接供给。有些动物的胚胎发育期在脱离卵膜的情况下进行,有的在卵膜或卵包囊内进行,更有的在母体内进行,这都与上述 3种营养方式有直接联系。主要根据有无幼虫期的出现,无脊椎动物的胚胎发育分为以下几种类型。
自由幼虫发育类型 发育过程中出现自由幼虫者称之。此型遍及多细胞无脊椎动物各个门类。海生无脊椎动物中,有70%以上种类具有浮游幼虫。一般说来,自由幼虫较多出现于低等无脊椎动物门类和某一门类内较原始种类中,而较少出现于高等门类和特化种类中。这一情形与卵子含卵黄量多少有直接关系,是物种在历史发展中与环境条件相统一的结果。
自由幼虫不仅具有临时性执行感觉、行动和营养功能的幼虫器官,有独立生活方式,甚至不同于其成体所属生态类群。
自由幼虫发育到一定程度之后即进行变态。此一过程随物种不同而有差异。如有些甲壳动物由结构简单的早期幼虫开始,中间经过若干不同演化水平的幼虫期,最后达到成体形态,是称演化性变态。与此相反,有些动物的幼虫如帽状幼虫和后湾幼虫,在变态期间几乎要失去所有幼虫器官和身体的很大部分,是称坏死性变态。
这一发育类型的优点在于能够节省亲体营养供应,以便亲体多产卵子。对底栖动物来说,通过幼虫的活动,种的分布可以得到扩大。其不利之点是容易受到环境条件变化的影响,以致幼虫死亡或不能完成变态。
直接发育类型 特点是形态发生主要在卵膜内完成。在此期间,胚体依靠亲体预先准备好的大量卵黄作为营养。最后由卵膜中孵出的不是幼虫,而是幼体。
许多例子说明:直接发育类型是在自由幼虫发育类型基础上演化而成。如有些多毛类动物的早期胚胎发育是在卵膜内渡过的,可是在此期间仍有形态改变的幼虫器官。又如高等虾类已失去了某些自由幼虫期,但实际上在卵膜内还要出现与无节幼虫和水蚤幼虫相当的发育阶段。
从自由幼虫发育类型向直接发育类型过渡的直接原因是卵黄量的增加,根本原因是为适应环境的需要。如生活在潮下带的乳白纵沟纽虫具有典型帽状幼虫期,而生活在潮间带的德沙纵沟纽虫,由于受干燥的影响失去了自由幼虫期。在这种情形下,卵黄含量增加,卵子相应变大,胚胎期延长,幼虫形态减少。与帽状幼虫相比较下,德沙纵沟纽虫的胚胎期缺少了顶感觉器和口叶等幼虫器官,但仍出现胚盘,因之称为德沙氏幼虫。这种幼虫在卵膜内完成变态,所以德沙纵沟纽虫的胚胎发育应归入直接发育类型。
对现存昆虫类动物来说,发育类型的演化与上述情况不同。无翅类和某些低等有翅类都进行直接发育,但到高等有翅类反而又有了幼虫和变态,从而成为自由幼虫发育类型。这种幼虫有后成自由幼虫或次级自由幼虫之称。
非自由幼虫发育类型 在此类型中,幼虫形态仍有不同程度存在,但与自由幼虫类型相较,显然有所不同。可分为以下3个亚型:
寄生幼虫 不仅见之于寄生动物,也见之于某些营自由生活的动物。吸虫类的雷叠幼虫和绦虫类的囊尾幼虫属前者;刚水母的实囊幼虫和无齿蚌的钩介幼虫属于后者。寄生幼虫的特点是从寄主体上获得营养。
包囊幼虫 此种幼虫因在由亲体制成的包囊中发育而得名(图26)。
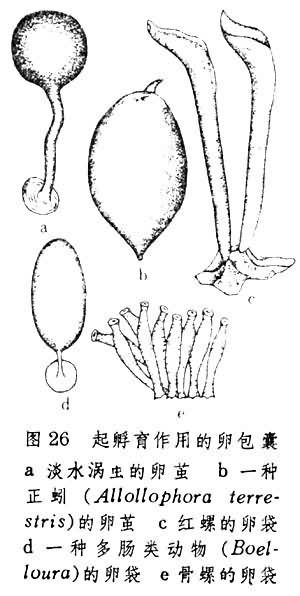 包囊一般是指具有三级卵膜意义的卵袋。此外,如涡虫类卵子外面的硬壳,后鳃类和多毛类中的胶质卵块和寡毛类动物中卵囊等也都具有包囊意义。包囊内除含有正常卵子外,还可储存营养物质或不能发育的卵子(败育卵或称营养细胞)。发育中的胚胎即以此为营养。有些陆生寡毛类动物(Oligochaeta megadrili)和颚蛭类动物(gnathobde-llide nephelis) 的卵袋中含有蛋白质团块和液体。由正常卵发育成的胚胎可利用暂时性纤毛构造游动于团块之间并以之为食。
包囊一般是指具有三级卵膜意义的卵袋。此外,如涡虫类卵子外面的硬壳,后鳃类和多毛类中的胶质卵块和寡毛类动物中卵囊等也都具有包囊意义。包囊内除含有正常卵子外,还可储存营养物质或不能发育的卵子(败育卵或称营养细胞)。发育中的胚胎即以此为营养。有些陆生寡毛类动物(Oligochaeta megadrili)和颚蛭类动物(gnathobde-llide nephelis) 的卵袋中含有蛋白质团块和液体。由正常卵发育成的胚胎可利用暂时性纤毛构造游动于团块之间并以之为食。胎生 这种发育在广大无脊椎门类中都有出现。优点是胚胎能够直接得到亲体的护养。怀胎的地点和方式因种类而异。如樽海绵中胶层内的间质细胞可围成胎腔以纳卵子;一种海葵(Siderastraea)的早期胚胎在隔间腔中发育;轮虫类(如晶囊轮虫 Asplanchna 和轮虫Rotaria)卵子留在输卵管内发育;一种单肠类动物 (Me-sostoma)和田螺的卵子都在子宫中发育。在上述各例中,胚胎都是以亲体分泌出的物质或本身原有卵黄作为营养,所以这种胎生被称为卵胎生。
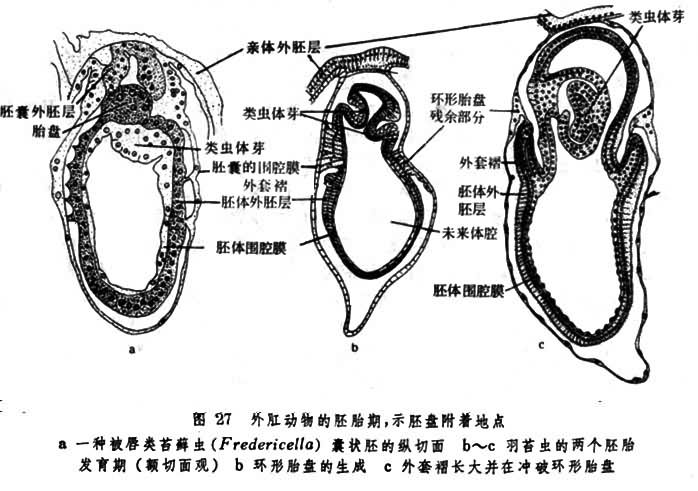
真胎生仅出现于少数无脊椎动物门类中。胚胎通过胎盘或其他结构直接自亲体得到营养的情形称真胎生,这也是和卵胎生相区别的唯一标准。无胎盘的真胎生极少见。一种多毛类动物(Ctenodrilus)的胚胎依靠亲体血管供给营养是其中一例。在有胎盘的情形下,其构造也简单,只不过是由胚体表面生出的外胚层皱褶而已。胎盘与亲体相接触的地点因种而异。一种有爪类动物(Epi-peripatus trinidadensis)的胎盘附着于亲体输卵管壁上,有些被唇动物(Fredericella,Plumatella)的胎盘种植在亲体卵房内(图27)。
发育方式
有性方式的发育 又可分为两性发育和单性发育。前者在无脊椎动物中正统而普遍。后者限为少数物种所有,过程始于未受精卵,起加速繁殖作用。
无性方式的发育 这种发育不是开始于雌性生殖细胞,而开始于一个芽状构造或球形小体。这两种情形分别称为外出芽和内出芽(见无性生殖)。由芽体长成新个体的过程与有性方式中的器官发生过程基本相同。
在腔肠动物的有刺胞类中,外出芽成为普遍现象。如多态性管水母的组成个员不但都系芽生而且在生成过程中各有一定方式和顺序,所以这类动物整个群体的形成本属胚胎发育过程,而实际上却是芽生过程。在这里胚胎发育和芽生完全是一回事。
在苔藓动物外出芽的萌发过程中,先有部分芽体外胚层进行内陷以形成一个囊状构造。此囊在形态、地位和作用上都相当于有性方式发育中的长形囊状胚。新个体即在此囊基础上生成。其过程几乎完全雷同于第一个类虫体在囊状胚基础上生成的过程。
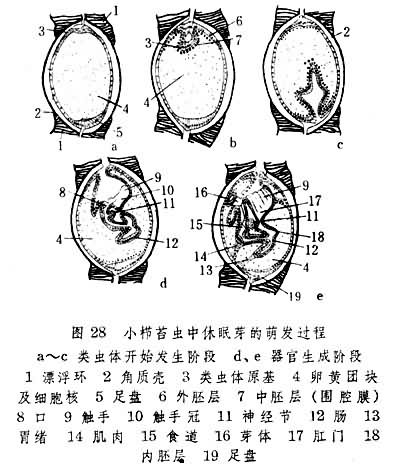
再一个例子则是苔藓动物中经内出芽法生成的休眠芽。其萌发过程(图28)既雷同外出芽过程,也近似有性方式发育中第一个类虫体生成过程。在这种情形下,该动物无性方式发育与有性方式发育间的差别大为缩小。
再生 在再生过程中,细胞加速分裂,同时由非特化转向特化从而导致形态发生,这是其与胚胎发育相似之处(见再生(动物))。
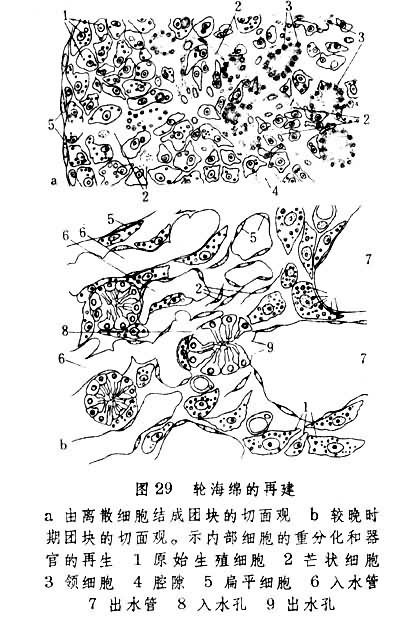
大多数无脊椎动物具有再生能力,惟程度各有不同。分散的海绵细胞经过聚合及重分化后可形成新个体。这种强能力再生现象称为形态上的再建(图29)。有些无脊椎动物能将伤残身体改建为较小的完整个体,是为变形再生。如果再生部分是新生的,则称为新建再生。也有些动物如淡水涡虫的再生兼有以上两种内容。
参考书目
E.A.施密特著,李维恩等译:《动物胚胎学》,上、下卷,高等教育出版社,北京,1956。
E.考尔舍尔特著,庄孝僡译:《比较胚胎学》,上卷,科学出版社,北京,1957。
L.H.Hymen,The Invertebrates, Vol. 1~5,McGraw-Hill Book Co.,Inc.,New York,1940~1959.
G.G.Karl,Protozoology,Springer-Verlag,Berlin,NewYork,1973.
E.W.MacBride,Text-Book of Embryology,Macmillan & Company,Ltd.,London,1914.
G.Reverberi,Experimental Embryology of Marineand Freshwater Invertebrates, North Holland Publishing Co.,Amsterdam,London,1971.
C.Dawydoff,Traité D'Embroyologie Comparée des Invertebrés,Masson Cie Editeurs,Paris,1928.