细胞周期调节(卷名:现代医学)
cell cycle regulation
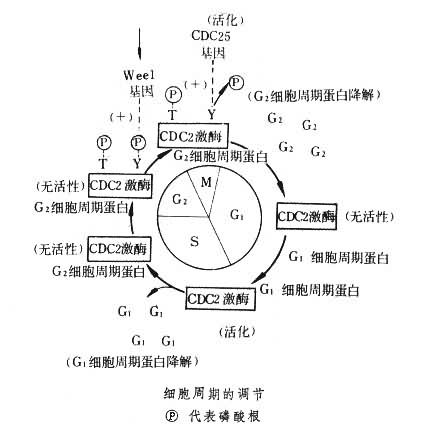
细胞周期是单个真核细胞生长和分裂成子细胞的过程,可分为G1、S、G2及M各期。这是由许多细胞周期调节因子参与调控的生物化学过程。其中起主要作用的是由细胞分裂周期基因(CDC2)的产物及细胞周期蛋白构成的 M期促发因子(MPF)。MPF的磷酸化(失活)和脱磷酸化(活化)是控制细胞周期自G2进入M期的关键。
细胞周期调节的模式 细胞周期大致可分为四期:G1期,细胞开始生长;S期,DNA合成,染色体复制;G2期,第二生长期;M期,细胞有丝分裂。早在1900年前后,科学家已经认识到细胞周期的各种基本活动,但只是80年代末才揭示了调节细胞周期的主要生物化学过程,并获得自酵母至人类普遍适用的模式。
过去一直认为细胞周期受核的控制,细胞浆的活动是被动跟随。例如,蛙卵在进入有丝分裂时会突然收缩,导致细胞分裂,这显然是核分裂所诱发的。有人发现,即使将核去掉,细胞也会周期性地收缩。这表明胞浆中存在有调节细胞周期的因子,其活动不完全受控于细胞核的活动,这种因子称为M期促发因子(MPF)。后又有人用实验证明,当细胞因蛋白质合成被抑制而停留在分裂间期时,注入含活性的 MPF提取液,可促进细胞进入有丝分裂(M期),这表明MPF是M期的正常诱导物。
另有人则从遗传学角度进行研究,发现在啤酒酵母(Sacharomyces cerevisiae) 的某些突变株中,其细胞周期可被“截止”在几个特定点上。这些突变株中,每个突变基因的产物与细胞周期通过各特定点有关,这些基因统称为细胞分裂周期基因(CDC基因),他们并进一步阐明这些CDC基因激活的顺序。在已发现的多种CDC基因中,以CDC2最为重要。它的表达产物是分子量为34kd的蛋白质,称P34CDC2,对细胞进入有丝分裂是关键性的。1988年首次分离到MPF,并获悉MPF由两种蛋白质分子所组成,其中之一是CDC2基因产物。往蛙卵的细胞质提取液中,加入精子的核,则在试管中核的有丝分裂(DNA的合成)可被诱导进行;但若去除CDC2蛋白,则DNA的合成将中止,核也就不能进行有丝分裂。这说明CDC2在细胞周期中至关重要。它在有丝分裂过程中被激活,而在分裂期后则失活,于是造成细胞周期的周而复始,它的突变将阻止细胞进行有丝分裂。CDC2蛋白的浓度在细胞周期过程中始终是稳定的;而MPF的另一组分在每次分裂间期时被合成,而在有丝分裂时则被降解,其含量发生周期性的波动;这个组分是CDC2活性的调节单位,称为细胞周期蛋白(亦译循环素)。
细胞周期蛋白有多种,由蛙卵中分离所得者为分子量45kd的蛋白质,酵母细胞周期蛋白的分子量为56kd。以RNA酶破坏蛙卵细胞质提取液中的所有RNA,若另单加海蛤周期蛋白的mRNA,又可使精子的核进行有丝分裂。选择性地阻止细胞周期蛋白的合成,而并不干扰其他蛋白质的合成,结果在上述“蛙卵提取液-精子核”的体系中,使核停留在分裂间期。以上两方面的实验异曲同工地证明细胞周期蛋白在细胞周期中的重要性。
由此可见,MPF是由具有催化功能的P34CDC2及具有调节功能的细胞周期蛋白组成的复合体。细胞周期蛋白在有丝分裂时被降解,在分裂间期时被重新合成,以调解P34CDC2的活性。现知P34CDC2是丝/苏氨酸蛋白激酶当它被激活时,可磷酸化某些转录因子,使一些与 DNA合成有关或与细胞分裂有关的酶和蛋白质获得表达,或直接激活某些与细胞周期有关的蛋白质,以推动细胞周期的运行。
细胞周期中有两个突发的转折点:G1→S及G2→M、只有在此两个转折点时,P34CDC2才突显活性。在G1期时,P34CDC2不显活性,随着G1细胞周期蛋白(包括CLN1、CLN2、CLN3) 的逐渐积累合成,两者结合成复合体 MPF,此时P34CDC2方显示苏/丝氨酸蛋白激酶活性,激活某些转录因子,使与 DNA合成有关的基因获得表达,细胞乃进入S期,积极复制DNA。
随之,G1细胞周期蛋白降解, P34CDC2失活,细胞进入G2期。在G2期时,蛋白质积极合成,细胞体积逐渐增大,为分为两个子细胞作好准备。同时G2细胞周期蛋白(包括CLB1、CLB2、CLB3、CLB4)又渐积累合成(G1细胞周期蛋白与G2细胞周期蛋白是不同的),并又与P34CDC2结合成 MPF前体。正由于G2细胞周期蛋白不同于G1细胞周期蛋白,所以经它调节控制的P34CDC2的底物特异性也各异。
值得注意的是有丝分裂的发生是一个突发的过程,而G2细胞周期蛋白的合成是一个逐渐累积合成的过程,在G2细胞周期蛋白没有积累到足够量时,P34CDC2不宜表现活性。要解决这个问题,办法是通过另一基因产物对P34CDC2进行磷酸化修饰,经磷酸化的P34CDC2暂时不表现蛋白激酶活性。现在已获知这一基因为wee1基因,其产物是分子量为 107kd的蛋白质,具有蛋白激酶活性,能特异性地对P34CDC2上的酪氨酸15进行磷酸化。同时还有另外的蛋白激酶将苏氨酸167和(或)苏氨酸14磷酸化。一旦当有促分裂的因素(如分裂素)刺激存在,它即激发一种酪氨酷蛋白磷酸酶,将P34CDC2上的酪15磷酸水解下来,P34CDC2的蛋白激酶活性乃被激活。编码这一酪氨酸蛋白磷酸酶的基因称 CDC25,这是启动有丝分裂的关键因素。随着 P34CDC2的活化,许多与有丝分裂有关的蛋白质被磷酸化而发挥生理效应,如层连蛋白磷酸化,即导致核膜解体;组蛋白H1磷酸化,即导致染色质聚合;微管蛋白磷酸化,即导致细胞骨架的重新组合,等等,这便推进有丝分裂过程。与此同时,G2细胞周期蛋白也被磷酸化,特异性地降解G2细胞周期蛋白的泛蛋白连接酶体系也被磷酸化而激活,促进G2细胞周期蛋白的迅速降解。G2细胞周期蛋白的降解,使P34CDC2失去调节蛋白(即G2细胞周期蛋白)而又失活,细胞 B又趋于静止期。等到新的 G1细胞周期蛋白合成,细胞方能又由G1期进入S期,开始一轮新的细胞周期。细胞周期就是这样周而复始地进行。
强调 MPF为主的细胞周期调节因子对细胞周期的控制和推进作用,并不应忽视核对细胞周期的深刻影响。通常当 DNA有损或复制不完全时,RAD9基因发挥作用,可抑制G2细胞周期蛋白(CLB1,CLB2)并使P34CDC2高度磷酸化而失活,以阻止有丝分裂。但在此时,若加入CDC25产物(即能使P34CDC2的酪氨酸15脱磷酸的酪氨酸蛋白磷酸酶),则即使DNA复制不完全,细胞也可进入M期,由此足见细胞周期调节因子的重要性。
上述细胞周期的调节过程(见图)仅是一个简单的模式,随着研究的深入,发现的调节因子愈来愈多,其影响也更错综复杂,但最基本的调节机构仍是如上所描述的普遍模式。现已发现的细胞周期蛋白有 A、B、C、D、E等多种;CDC2在酵母中只有一种,但在高等真核细胞(蛙卵、果蝇细胞等)中则有两种:CDC2B产物,为P32CDC2B,调节G1→S;CDC2A产物,为P34CDC2B,调节G2→M。调节 P34CDC2磷酸化及脱磷酸化的酶也有多种,如为酪氨酸蛋白激酶进行编码的基因有weel及niml;为酪氨酸蛋白磷酸酶编码的基因有bws1,bim G及dis 2等。此外,还有其他因子间接地参与调节,从而使细胞周期的调节十分精密,不至因为某一环节的失常而出现细胞的异常分裂。
细胞周期调节与生命科学 细胞周期的研究是细胞生物学中最根本的问题。例如与生长增殖有关激素、生长因子、癌基因及抗癌基因的作用机理,必然直接间接地与细胞周期有关;胚胎的有序发育,成熟机体的健康,组织的再生及衰老也无不与细胞周期密切相关。某些细胞周期蛋白基因(主要是G1细胞周期蛋白)本身就是原癌基因,如甲状旁腺瘤的癌基因PRAD1已证明就是细胞周期蛋白Di。有的癌基因产物(如c-src及c-abl)是酪氨酸蛋白激酶,可被P34CDC2-G2细胞周期蛋白所激活。人肝细胞癌的肝炎病毒 B基因片段,可整合到细胞周期蛋白A基因所在部位,使细胞周期蛋白A过量表达而致癌变。又通常情况下转录因子 E2F与抗癌基因RB或P53结合成复合物,减弱了E2F的转录活性;而腺病毒转化蛋白E1A则能竞争性地与抗癌基因RB或 P53结合,释出游离E2F,大大增强了E2F的转录作用,使与细胞增殖有关的早期基因(如c-fos,c-myc等)过量表达而致癌。鉴于细胞周期的推动主要决定于P34CDC2蛋白激酶的激活和失活,即磷酸化和脱磷酸化,这又由weel及CDC25两基因所控制,因此,有人认为此两基因表达比例的失常,也许是细胞衰老式癌变的原因之一。总之,细胞周期调节的研究对于细胞生物学、分子生物学和医学具有十分重要的理论和实际意义,是现代生物学中的一个根本问题。
参考书目
A.W.Murray & M.W.Kirschner: What Controls the Cell Cycle, Scientific American, No.3,56~63,1991.