双受精(卷名:生物学)
double fertilization
被子植物的雄配子体形成的两个精子,一个与卵融合形成二倍体的合子,另一个与中央细胞的极核(通常两个)融合形成初生胚乳核的现象。双受精后由合子发育成胚,初生胚乳核发育成胚乳。(见彩图)

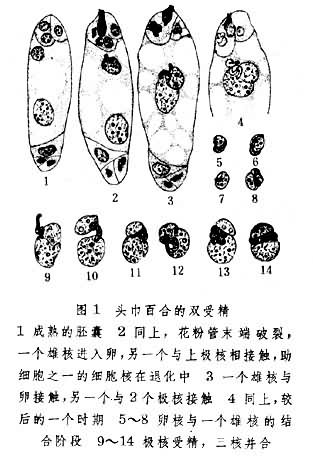
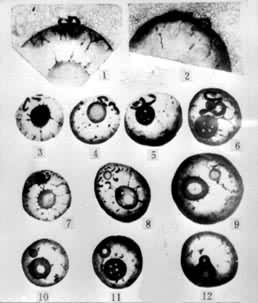
1880年,俄国植物胚胎学家И.Н.戈罗然金首先发现了裸子植物配子的融合现象。4 年后德国植物胚胎学家、细胞学家Е.А.施特拉斯布格在被子植物中也看到了这种现象。他在水晶兰中观察到胚囊中由花粉管释出两个雄性核(精子),其中一个与卵结合;当时还不了解第2个精子的命运。1898年,俄国植物胚胎学家С.Г.纳瓦申研究了头巾百合和贝母的受精作用,才发现一个精子和卵融合,而另一个和两个极核合并。这是双受精现象的最早发现。1899年,法国植物胚胎学家L.吉尼亚尔也报道了百合属和贝母属中同样的现象,并且绘出一套图(图1)。以后证实双受精是被子植物普遍存在的现象。
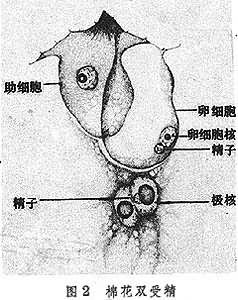
被子植物胚囊中极核同卵一样受精产生了具有父本和母本遗传性的通常是三倍体的胚乳,由这种胚乳“哺育”胚可能使后代更加巩固它双亲的特性并更富有生命力,因此有人认为双受精是被子植物繁盛的一个重要原因。
配子配合前的阶段 在这一段时期,发生的事态有:
花粉萌发和花粉管在花柱中生长 花粉由各种媒介传到雌蕊的柱头上后,立即与雌蕊发生相互作用(见受精(植物))。在亲和的情况下,花粉萌发长出花粉管并钻入柱头。花粉管进入柱头后,继续在花柱中生长。不同植物的花柱在结构上是不同的。主要有两种形式:①花柱中有宽广的中空花柱道(如百合);②花柱是实心的,中央常有特殊的引导组织(如棉花、番茄)。花粉管通过中空的花柱道时,一般是沿着通道表面及在通道细胞分泌的物质中生长。在实心花柱中,花粉管通常是在引导组织充满基质的细胞间隙中穿行。一般认为胞间隙中的基质成分是碳水化合物,主要是果胶物质。此外,花粉管还可能穿过引导组织的细胞壁生长(如棉花)或从细胞壁与质膜之间穿行(如菠菜)。
花粉管依赖其末端生长,当生长到一定长度后,原来在花粉中含有的物质全部集中到花粉管的前端。在生长过程中花粉管除利用自身的贮藏物质外,同时也可能从花柱中吸收营养。
花粉管进入胚囊和释放内容物 花粉管伸入子房后,沿子房内壁或胎座继续生长,直达胚珠,经珠孔进入胚珠,最后到达胚囊。关于花粉管沿一定道路生长的现象,一般认为是受化学物质的吸引,多数学者认为这种化学物质是由助细胞分泌的。
花粉管进入胚囊后的途径也有各种说法,如穿入一个助细胞、从卵与一助细胞之间或卵与胚囊壁之间通过等等。经过对十多种植物,如棉花、玉米、矮牵牛等的电子显微镜研究,证实花粉管是从一个助细胞进入的,而且所进入的那个助细胞常常是在花粉管进入之前解体。花粉管就在此助细胞的位置释放出内容物。
通过电子显微镜观察发现有些植物的花粉管的末端(如矮牵牛、荠菜)或末端一侧(如棉花)具有一孔,因此推测花粉管中的精子和其他内容物是通过此孔释放的。
配子配合 通常把精子与卵的融合称为配子配合,而精子与极核的融合称为三核并合。这两种融合是差不多同时发生的(图2)。关于配子配合过程的研究主要集中在下列几方面:
雄配子与雌配子是如何并合为一体的 过去有人把融合比作两个油滴的并合,或推测是由于两种配子的原生质体接触后质膜突然破裂而得以融合。20世纪60年代后应用电子显微镜研究已明确精子与雌配子是通过局部质膜的接触和溶解而融合的。1973年,美国植物胚胎学家W.A.詹森提出了被子植物双受精作用中精子转移至卵细胞和中央细胞的模式图(图3):
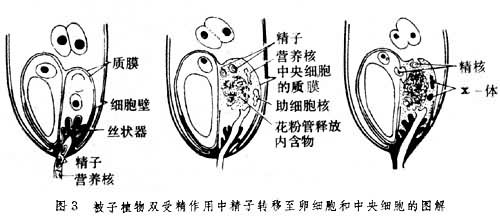 精子先释放至退化的助细胞(质膜已经消失),由于花粉管内容物释放时的力量,两个精子被分别转移至卵细胞的质膜和中央细胞的质膜处并与之接触,接触处的质膜溶解,两个精核分别转移至卵和中央细胞的细胞质中。
精子先释放至退化的助细胞(质膜已经消失),由于花粉管内容物释放时的力量,两个精子被分别转移至卵细胞的质膜和中央细胞的质膜处并与之接触,接触处的质膜溶解,两个精核分别转移至卵和中央细胞的细胞质中。精子的细胞质是否参与融合 按W.A.詹森的精子入卵模式,只是精子核转移至雌性细胞中。实际上在被子植物的受精作用中,精子的细胞质是否参加迄今尚无定论。应用电子显微镜的研究,也得出两种不同的结论:如在棉花的受精卵内看不到精子的细胞质,推测只有核参加受精;但在红萼月见草的合子中则发现有精子的质体;在白花丹受精后的卵和中央细胞中也发现有精子的线粒体和质体,表明细胞质也参与融合。
核的融合 配子融合的最后一步。1933年,苏联植物胚胎学家Е.Н.格拉西莫娃-纳瓦希娜和С.Г.纳瓦申对一种菊科植物──还阳参受精作用中精核与卵核的融合作了相当详细的描述并绘有精美的图(图4)。
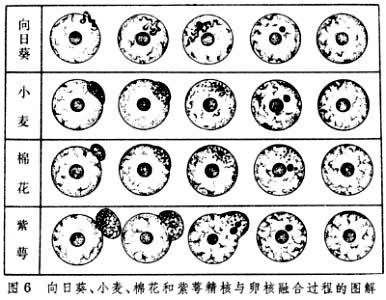
 根据1979年中国植物胚胎学家胡适宜等对向日葵、小麦、棉花和紫萼几种植物所作的详细观察证明被子植物雄核与雌核融合的基本程序是:①雄性核与雌性核接触;②雄性核和雌性核之间的核膜融合;③精子核的染色质在卵核内分散和出现雄性核仁;④雄性核的染色质与雌性核的染色质混合;⑤雄性和雌性的核仁融合为一个大的核仁(图5、图6)。
根据1979年中国植物胚胎学家胡适宜等对向日葵、小麦、棉花和紫萼几种植物所作的详细观察证明被子植物雄核与雌核融合的基本程序是:①雄性核与雌性核接触;②雄性核和雌性核之间的核膜融合;③精子核的染色质在卵核内分散和出现雄性核仁;④雄性核的染色质与雌性核的染色质混合;⑤雄性和雌性的核仁融合为一个大的核仁(图5、图6)。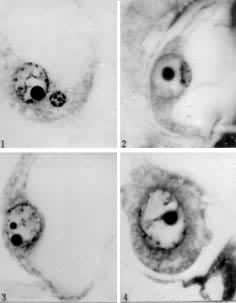
精核同极核(或次生核)的融合过程与精核同卵核的融合程序基本相同。
从花粉落到柱头上至雄性和雌性核完全融合所经历的时间,因植物不同而异。间隔期短的只有十几分钟(如橡胶草),长的可达一年(如栎属)。大多数植物为几小时至48小时之间,例如向日葵是3小时,花生是10~18小时,棉花是20~24小时,甘蓝是36~48小时,水稻是1~2小时,小麦是1~4小时,玉米是25~26小时。精卵融合与精子同极核融合通常是同时发生的,也可能精子同极核的融合更早发生,而且一般更快完成融合和进入分裂。这样胚乳的发育早于胚,可以为胚的发育提供必需的营养物质。
传粉至受精间隔期的长短还受环境因素,特别是温度的影响。温度对花粉萌发和花粉管的生长速度有直接的影响。每种植物花粉萌发和花粉管生长都有自己最适的温度,低温可使萌发和生长受抑制,从而延长花粉管到达胚囊的时间。
受精的类型 1961年Е.Н.格拉西莫娃-纳瓦希娜把被子植物受精划分为3种形式,即有丝分裂前的类型、有丝分裂后的类型和中间型。她认为精子在受精前处于有丝分裂末期,而卵是处于有丝分裂静止期。在配子融合时精子表现的行为不同是她划分受精类型的标准。在核融合过程中,精核与卵核接触后立即溶解卵的核膜并进入其中,在卵核内过渡到休眠期,融合发生在合子的有丝分裂之前,这属于有丝分裂前的类型,见于禾本科和菊科。有丝分裂后的类型的特点是精核在开始时不钻进卵核中,雄核在和卵核靠近时处于休眠状态。融合发生在合子的第一次分裂前期至中期。百合和香堇属于这种形式。中间类型是处于前两种类型之间的情况,即精核在进到卵核之前进入静止状态,在与卵核融合过程中同时开始合子的分裂。见于紫茉莉、凤仙花及紫露草等植物(图7)。

在种子植物受精作用中存在不同的核融合方式。目前,详细研究过的几种松科植物,如油松、白皮松和欧洲黑松等,表现为精子核进入卵核后,各自独立形成雄性及雌性的染色体组和自己的纺锤体,真正的融合是在受精卵有丝分裂之后;而详细研究过的几种被子植物,向日葵、小麦、棉花和紫萼,是雄性核和雌性核的染色质混合后才分裂。
卵细胞在受精前后的生理生化的变化 受精前卵细胞的超微结构的特点是:线粒体和高尔基器少,核糖体很少而且不聚集成多体。代谢和合成活动强度比较低。在受精以后,合子中的各种细胞器增加和重新分布,核常常被大量的造粉质体和线粒体包围,核糖体聚集成多聚核糖体,呈代谢活跃状态。
在受精前和受精后卵细胞核中脱氧核糖核酸 (DNA)的含量,根据孚尔根反应方法的测定是有变化的。应用这种方法已经确定,卵细胞在发育早期是强的孚尔根正反应,而达到成熟时变为弱的正反应或负反应。受精以后形成的合子中是正的孚尔根反应。对卵细胞在发育过程中所表现的孚尔根反应强弱的变化是否代表了核中 DNA含量的变化曾有争论。70年代以来,经荧光显微镜和紫外光显微分光光度计研究的结果,证明核不显示孚尔根反应不一定缺少DNA或DNA分子的核苷酸,也就是说卵细胞核在任何时期都含有DNA。
多精子进入胚囊和多精入卵 在正常的情况下,一个胚囊只接受两个精子,但是有时由于不止一个花粉管进入或是由于一个花粉管中形成不止一对精子,于是胚囊中出现有多于一对精子的现象。胚囊中额外精子的存在将有可能引起下面两种异常受精作用:①多精入卵。有两种不同的情况:一种是进入卵中多余的精子在卵中退化,另一种是附加的精子也与卵核融合,这种异常的受精形式只在极少数植物中有细胞学的证据。例如在喜马凤仙花中根据在受精的卵核内看到两个精子核的轮廓,认为是双精子受精的。②额外的精子与胚囊中的其他细胞受精。 这种现象曾在一种慈菇属植物(Sagittariagraminea)报道过。这种植物的助细胞形态上与卵细胞相似,有两个花粉管进入胚囊并放出额外的两个精子,在受精后的胚囊中有时看到 3个原胚存在,并有两个花粉管在胚囊的上部。因此推测助细胞与进入的额外精子受精而产生了多胚。在还阳参中曾观察到有些胚囊中含有2~5个卵细胞和有时一个花粉管不止两个精子进入胚囊的情况,以及在受精后的胚囊中存在两个受精卵在分裂的现象,推测有可能发生多于一个卵受精。
参考书目
胡适宜:《被子植物胚胎学》,高等教育出版社,北京,1982。
P.Mabeshwai,An introduction to the Embryology of Angiosperm, McGraw-Hill Book Co.Inc.,New York,1950.