reproductive system, human
Introduction
 organ system by which humans reproduce and bear live offspring. Provided all organs are present, normally constructed, and functioning properly, the essential features of human reproduction are (1) liberation of an ovum, or egg, at a specific time in the reproductive cycle, (2) internal fertilization of the ovum by spermatozoa, or sperm cells, (3) transport of the fertilized ovum to the uterus, or womb, (4) implantation of the blastocyst, the early embryo developed from the fertilized ovum, in the wall of the uterus, (5) formation of a placenta and maintenance of the unborn child during the entire period of gestation, (6) birth (parturition) of the child and expulsion of the placenta, and (7) suckling and care of the child, with an eventual return of the maternal organs to virtually their original state.
organ system by which humans reproduce and bear live offspring. Provided all organs are present, normally constructed, and functioning properly, the essential features of human reproduction are (1) liberation of an ovum, or egg, at a specific time in the reproductive cycle, (2) internal fertilization of the ovum by spermatozoa, or sperm cells, (3) transport of the fertilized ovum to the uterus, or womb, (4) implantation of the blastocyst, the early embryo developed from the fertilized ovum, in the wall of the uterus, (5) formation of a placenta and maintenance of the unborn child during the entire period of gestation, (6) birth (parturition) of the child and expulsion of the placenta, and (7) suckling and care of the child, with an eventual return of the maternal organs to virtually their original state.
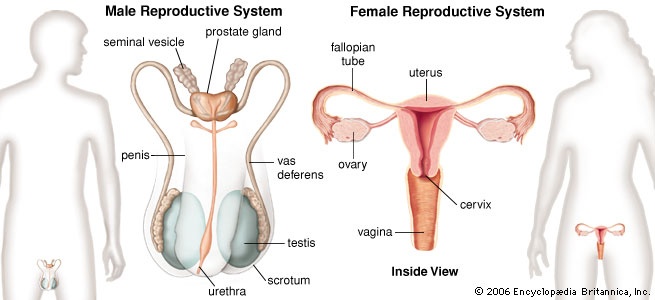
 For this biological process to be carried out, certain organs and structures are required in both the male and the female. The source of the ova (the female germ cells) is the female ovary; that of spermatozoa (the male germ cells) is the testis. In females, the two ovaries are situated in the pelvic cavity; in males, the two testes are enveloped in a sac of skin, the scrotum, lying below and outside the abdomen. Besides producing the germ cells, or gametes (gamete), the ovaries and testes are the source of hormones that cause full development of secondary sexual characteristics and also the proper functioning of the reproductive tracts. These tracts comprise the fallopian tubes (fallopian tube), the uterus, the vagina, and associated structures in females and the penis, the sperm channels (epididymis, ductus deferens, and ejaculatory ducts), and other related structures and glands in males. The function of the fallopian tube is to convey an ovum, which is fertilized in the tube, to the uterus, where gestation (development before birth) takes place. The function of the male ducts is to convey spermatozoa from the testis, to store them, and, when ejaculation occurs, to eject them with secretions from the male glands through the penis.
For this biological process to be carried out, certain organs and structures are required in both the male and the female. The source of the ova (the female germ cells) is the female ovary; that of spermatozoa (the male germ cells) is the testis. In females, the two ovaries are situated in the pelvic cavity; in males, the two testes are enveloped in a sac of skin, the scrotum, lying below and outside the abdomen. Besides producing the germ cells, or gametes (gamete), the ovaries and testes are the source of hormones that cause full development of secondary sexual characteristics and also the proper functioning of the reproductive tracts. These tracts comprise the fallopian tubes (fallopian tube), the uterus, the vagina, and associated structures in females and the penis, the sperm channels (epididymis, ductus deferens, and ejaculatory ducts), and other related structures and glands in males. The function of the fallopian tube is to convey an ovum, which is fertilized in the tube, to the uterus, where gestation (development before birth) takes place. The function of the male ducts is to convey spermatozoa from the testis, to store them, and, when ejaculation occurs, to eject them with secretions from the male glands through the penis.At copulation, or sexual intercourse, the erect penis is inserted into the vagina, and spermatozoa contained in the seminal fluid ( semen) are ejaculated into the female genital tract. Spermatozoa then pass from the vagina through the uterus to the fallopian tube to fertilize the ovum in the outer part of the tube. Females exhibit a periodicity in the activity of their ovaries and uterus, which starts at puberty and ends at the menopause. The periodicity is manifested by menstruation at intervals of about 28 days; important changes occur in the ovaries and uterus during each reproductive, or menstrual, cycle. Periodicity, and subsequently menstruation, is suppressed during pregnancy and lactation.
This articles describes the organs, both male and female, that are involved in human reproduction. The reproductive process itself is covered in other articles. For a detailed discussion of the series of changes that occur in a woman's body as her fetus develops, see pregnancy. For a description of the stages of labour and delivery, see parturition. For the development of the unborn child during gestation, see human embryology. For coverage of the many diseases and disorders that can affect the reproductive organs, see reproductive system disease.
Development of the reproductive organs
The sex of a child is determined at the time of fertilization of the ovum by the spermatozoon (sperm). The differences between a male and a female are genetically determined by the chromosomes (chromosome) that each possesses in the nuclei of the cells. Once the genetic sex has been determined, there normally follows a succession of changes that will result, finally, in the development of an adult male or female. There is, however, no external indication of the sex of an embryo during the first eight weeks of its life within the uterus. This is a neutral or indifferent stage during which the sex of an embryo can be ascertained only by examination of the chromosomes in its cells.
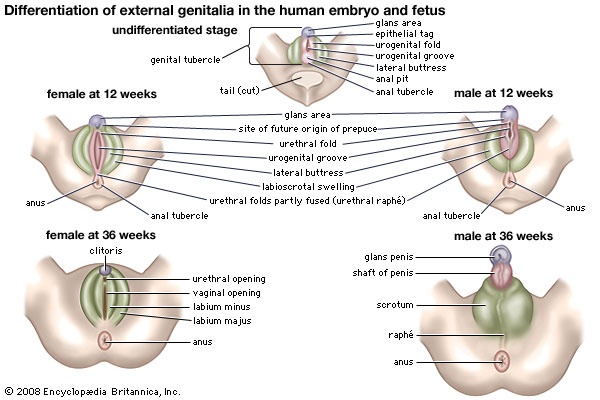 The next phase, one of differentiation, begins first in gonads (gonad) that are to become testes and a week or so later in those destined to be ovaries. Embryos of the two sexes are initially alike in possessing similar duct systems linking the undifferentiated gonads with the exterior and in having similar external genitalia, represented by three simple protuberances. The embryos each have four ducts, the subsequent fate of which is of great significance in the eventual anatomical differences between men and women. Two ducts closely related to the developing urinary system are called mesonephric, or wolffian, ducts (Wolffian duct). In males each mesonephric duct becomes differentiated into four related structures: a duct of the epididymis, a ductus deferens, an ejaculatory duct, and a seminal vesicle. In females the mesonephric ducts are largely suppressed. The other two ducts, called the paramesonephric or müllerian ducts, persist, in females, to develop into the fallopian tubes (fallopian tube), the uterus, and part of the vagina; in males they are largely suppressed. Differentiation also occurs in the primitive external genitalia, which in males become the penis and scrotum and in females the vulva (the clitoris, labia, and vestibule of the vagina).
The next phase, one of differentiation, begins first in gonads (gonad) that are to become testes and a week or so later in those destined to be ovaries. Embryos of the two sexes are initially alike in possessing similar duct systems linking the undifferentiated gonads with the exterior and in having similar external genitalia, represented by three simple protuberances. The embryos each have four ducts, the subsequent fate of which is of great significance in the eventual anatomical differences between men and women. Two ducts closely related to the developing urinary system are called mesonephric, or wolffian, ducts (Wolffian duct). In males each mesonephric duct becomes differentiated into four related structures: a duct of the epididymis, a ductus deferens, an ejaculatory duct, and a seminal vesicle. In females the mesonephric ducts are largely suppressed. The other two ducts, called the paramesonephric or müllerian ducts, persist, in females, to develop into the fallopian tubes (fallopian tube), the uterus, and part of the vagina; in males they are largely suppressed. Differentiation also occurs in the primitive external genitalia, which in males become the penis and scrotum and in females the vulva (the clitoris, labia, and vestibule of the vagina).At birth the organs appropriate to each sex have developed and are in their adult positions but are not functioning. Various abnormalities can occur during development of sex organs in embryos, leading to hermaphroditism, pseudohermaphroditism, and other chromosomally induced conditions. During childhood until puberty there is steady growth in all reproductive organs and a gradual development of activity. puberty marks the onset of increased activity in the sex glands and the steady development of secondary sexual characteristics.
In males at puberty the testes (testis) enlarge and become active, the external genitalia enlarge, and the capacity to ejaculate (ejaculation) develops. Marked changes in height and weight occur as hormonal secretion from the testes increases. The larynx, or voice box, enlarges, with resultant deepening of the voice. Certain features in the skeleton, as seen in the pelvic bones and skull, become accentuated. The hair in the armpits and the pubic hair becomes abundant and thicker. Facial hair develops, as well as hair on the chest, abdomen, and limbs. Hair at the temples recedes. Skin glands become more active, especially apocrine glands (a type of sweat gland that is found in the armpits and groin and around the anus).
In females at puberty, the external genitalia enlarge and the uterus commences its periodic activity with menstruation. The breasts develop, and there is a deposition of body fat in accordance with the usual contours of the mature female. Growth of axillary (armpit) and pubic hair is more abundant, and the hair becomes thicker.
The male reproductive system
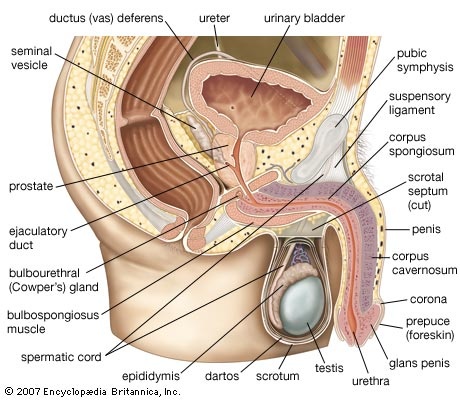
The male gonads are the testes; they are the source of spermatozoa and also of male sex hormones called androgens (androgen). The other genital organs are the epididymides; the ductus, or vasa, deferentia; the seminal vesicles; the ejaculatory ducts; and the penis; as well as certain accessory structures, such as the prostate and the bulbourethral (Cowper) glands. The principal functions of these structures are to transport the spermatozoa from the testes to the exterior, to allow their maturation on the way, and to provide certain secretions that help form the semen.External genitalia
The penis
 The penis, the male organ of copulation, is partly inside and partly outside the body. The inner part, attached to the bony margins of the pubic arch (that part of the pelvis directly in front and at the base of the trunk), is called the root of the penis. The second, or outer, portion is free, pendulous, and enveloped all over in skin; it is termed the body of the penis. The organ is composed chiefly of cavernous or erectile tissue that becomes engorged with blood to produce considerable enlargement and erection. The penis is traversed by a tube, the urethra, which serves as a passage both for urine and for semen.
The penis, the male organ of copulation, is partly inside and partly outside the body. The inner part, attached to the bony margins of the pubic arch (that part of the pelvis directly in front and at the base of the trunk), is called the root of the penis. The second, or outer, portion is free, pendulous, and enveloped all over in skin; it is termed the body of the penis. The organ is composed chiefly of cavernous or erectile tissue that becomes engorged with blood to produce considerable enlargement and erection. The penis is traversed by a tube, the urethra, which serves as a passage both for urine and for semen.The body of the penis, sometimes referred to as the shaft, is cylindrical in shape when flaccid but when erect is somewhat triangular in cross section, with the angles rounded. This condition arises because the right corpus cavernosum and the left corpus cavernosum, the masses of erectile tissue, lie close together in the dorsal part of the penis, while a single body, the corpus spongiosum, which contains the urethra, lies in a midline groove on the undersurface of the corpora cavernosa. The dorsal surface of the penis is that which faces upward and backward during erection.
The slender corpus spongiosum reaches beyond the extremities of the erectile corpora cavernosa and at its outer end is enlarged considerably to form a soft, conical, sensitive structure called the glans penis. The base of the glans has a projecting margin, the corona, and the groove where the corona overhangs the corpora cavernosa is referred to as the neck of the penis. The glans is traversed by the urethra, which ends in a vertical, slitlike, external opening. The skin over the penis is thin and loosely adherent and at the neck is folded forward over the glans for a variable distance to form the prepuce or foreskin. A median fold, the frenulum of the prepuce, passes to the undersurface of the glans to reach a point just behind the urethral opening. The prepuce can usually be readily drawn back to expose the glans.
The root of the penis comprises two crura, or projections, and the bulb of the penis. The crura and the bulb are attached respectively to the edges of the pubic arch and to the perineal membrane (the fibrous membrane that forms a floor of the trunk). Each crus is an elongated structure covered by the ischiocavernosus muscle, and each extends forward, converging toward the other, to become continuous with one of the corpora cavernosa. The oval bulb of the penis lies between the two crura and is covered by the bulbospongiosus muscle. It is continuous with the corpus spongiosum. The urethra enters it on the flattened deep aspect that lies against the perineal membrane, traverses its substances, and continues into the corpus spongiosum.
The two corpora cavernosa are close to one another, separated only by a partition in the fibrous sheath that encloses them. The erectile tissue of the corpora is divided by numerous small fibrous bands into many cavernous spaces, relatively empty when the penis is flaccid but engorged with blood during erection. The structure of the tissue of the corpus spongiosum is similar to that of the corpora cavernosa, but there is more smooth muscle and elastic tissue. A deep fascia, or sheet of connective tissue, surrounding the structures in the body of the penis is prolonged to form the suspensory ligament, which anchors the penis to the pelvic bones at the midpoint of the pubic arch.
The penis has a rich blood supply from the internal pudendal artery, a branch of the internal iliac artery, which supplies blood to the pelvic structures and organs, the buttocks, and the inside of the thighs. erection is brought about by distension of the cavernous spaces with blood, which is prevented from draining away by compression of the veins in the area.
The penis is amply supplied with sensory and autonomic (involuntary) nerves. Of the autonomic nerve fibres the sympathetic fibres cause constriction of blood vessels, and the parasympathetic fibres cause their dilation. It is usually stated that ejaculation is brought about by the sympathetic system, which at the same time inhibits the desire to urinate and also prevents the semen from entering the bladder.
The scrotum
The scrotum is a pouch of skin lying below the pubic symphysis and just in front of the upper parts of the thighs. It contains the testes and lowest parts of the spermatic cord. A scrotal septum or partition divides the pouch into two compartments and arises from a ridge, or raphe, visible on the outside of the scrotum. The raphe turns forward onto the undersurface of the penis and is continued back onto the perineum (the area between the legs and as far back as the anus). This arrangement indicates the bilateral origin of the scrotum from two genital swellings that lie one on each side of the base of the phallus, the precursor of the penis or clitoris in the embryo. The swellings are also referred to as the labioscrotal swellings, because in females they remain separate to form the labia majora and in males they unite to form the scrotum.
The skin of the scrotum is thin, pigmented, devoid of fatty tissue, and more or less folded and wrinkled. There are some scattered hairs and sebaceous glands (sebaceous gland) on its surface. Below the skin is a layer of involuntary muscle, the dartos, which can alter the appearance of the scrotum. On exposure of the scrotum to cold air or cold water, the dartos contracts and gives the scrotum a shortened, corrugated appearance; warmth causes the scrotum to become smoother, flaccid, and less closely tucked in around the testes. Beneath the dartos muscle are layers of fascia continuous with those forming the coverings of each of the two spermatic cords, which suspend the testes within the scrotum and contain each ductus deferens, the testicular blood and lymph vessels, the artery to the cremaster muscle (which draws the testes upward), the artery to each ductus deferens, the genital branch of the genitofemoral nerve, and the testicular network of nerves.
The scrotum is supplied with blood by the external pudendal branches of the femoral artery, which is the chief artery of the thigh, and by the scrotal branches of the internal pudendal artery. The veins follow the arteries. The lymphatic drainage is to the lymph nodes in the groin.
The testes (testis)
 The two testes (testis), or testicles, which usually complete their descent into the scrotum from their point of origin on the back wall of the abdomen in the seventh month after conception, are suspended in the scrotum by the spermatic cords. Each testis is 4 to 5 cm (about 1.5 to 2 inches) long and is enclosed in a fibrous sac, the tunica albuginea. This sac is lined internally by the tunica vasculosa, containing a network of blood vessels, and is covered by the tunica vaginalis, which is a continuation of the membrane that lines the abdomen and pelvis. The tunica albuginea has extensions into each testis that act as partial partitions to divide the testis into approximately 250 compartments, or lobules.
The two testes (testis), or testicles, which usually complete their descent into the scrotum from their point of origin on the back wall of the abdomen in the seventh month after conception, are suspended in the scrotum by the spermatic cords. Each testis is 4 to 5 cm (about 1.5 to 2 inches) long and is enclosed in a fibrous sac, the tunica albuginea. This sac is lined internally by the tunica vasculosa, containing a network of blood vessels, and is covered by the tunica vaginalis, which is a continuation of the membrane that lines the abdomen and pelvis. The tunica albuginea has extensions into each testis that act as partial partitions to divide the testis into approximately 250 compartments, or lobules.Each lobule contains one or more convoluted tubules, or narrow tubes, where sperm are formed. The tubules, if straightened, would extend about 70 cm (about 28 inches). The multistage process of sperm formation, which takes about 60 days, goes on in the lining of the tubules, starting with the spermatogonia, or primitive sperm cells, in the outermost layer of the lining. Spermatozoa (sperm) leaving the tubules are not capable of independent motion, but they undergo a further maturation process in the ducts of the male reproductive tract; the process may be continued when, after ejaculation, they pass through the female tract. Maturation of the sperm in the female tract is called capacitation.
Each spermatozoon is a slender elongated structure with a head, a neck, a middle piece, and a tail. The head contains the cell nucleus. When the spermatozoon is fully mature, it is propelled by the lashing movements of the tail.
The male sex hormone testosterone is produced by Leydig cells. These cells are located in the connective (interstitial) tissue that holds the tubules together within each lobule. The tissue becomes markedly active at puberty under the influence of the interstitial-cell-stimulating hormone of the anterior lobe of the pituitary gland; this hormone in women is called luteinizing hormone. Testosterone stimulates the male accessory sex glands (prostate, seminal vesicles) and also brings about the development of male secondary sex characteristics at puberty. The hormone may also be necessary to cause maturation of sperm and to heighten the sex drive of the male. The testis is also the source of some of the female sex hormone estrogen, which may exert an influence on pituitary activity.
Each testis is supplied with blood by the testicular arteries, which arise from the front of the aorta just below the origin of the renal (kidney) arteries. Each artery crosses the rear abdominal wall, enters the spermatic cord, passes through the inguinal canal, and enters the upper end of each testis at the back. The veins leaving the testis and epididymis form a network, which ascends into the spermatic cord. The lymph vessels, which also pass through the spermatic cord, drain to the lateral and preaortic lymph nodes. Nerve fibres to the testis accompany the vessels; they pass through the renal and aortic nerve plexuses, or networks.
Structures of the sperm canal
The epididymis (epididyme), ductus deferens (or vas deferens), and ejaculatory ducts (ejaculatory duct) form the sperm canal. Together they extend from the testis to the urethra, where it lies within the prostate. Sperm are conveyed from the testis along some 20 ductules, or small ducts, which pierce the fibrous capsule to enter the head of the epididymis. The ductules are straight at first but become dilated and then much convoluted to form distinct compartments within the head of the epididymis. They each open into a single duct, the highly convoluted duct of the epididymis, which constitutes the “body” and “tail” of the structure. It is held together by connective tissue but if unraveled would be nearly 6 metres (20 feet) long. The duct enlarges and becomes thicker-walled at the lower end of the tail of the epididymis, where it becomes continuous with the ductus deferens.
The ductules from the testis have a thin muscular coat and a lining that consists of alternating groups of high columnar cells with cilia (hairlike projections) and low cells lacking cilia. The cilia assist in moving sperm toward the epididymis. In the duct of the epididymis the muscle coat is thicker and the lining is thick with tufts of large nonmotile cilia. There is some evidence that the ductules and the first portion of the duct of the epididymis remove excess fluid and extraneous debris from the testicular secretions entering these tubes. The blood supply to the epididymis is by a branch from the testicular artery given off before that vessel reaches the testis.
The ductus deferens, or vas deferens, is the continuation of the duct of the epididymis. It commences at the lower part of the tail of the epididymis and ascends along the back border of the testis to its upper pole. Then, as part of the spermatic cord, it extends to the deep inguinal ring. Separating from the other elements of the spermatic cord—the blood vessels, nerves, and lymph vessels—at the ring, the ductus deferens makes its way through the pelvis toward the base of the prostate, where it is joined by the seminal vesicle to form the ejaculatory duct. A part of the ductus that is dilated and rather tortuous, near the base of the urinary bladder, is called the ampulla.
The ductus deferens has a thick coat of smooth muscle that gives it a characteristic cordlike feel. The longitudinal muscle fibres are well developed, and peristaltic contractions (contractions in waves) move the sperm toward the ampulla. The mucous membrane lining the interior is in longitudinal folds and is mostly covered with nonciliated columnar cells, although some cells have nonmotile cilia. The ampulla is thinner-walled and probably acts as a sperm store.
Accessory organs
The prostate (prostate gland) gland, seminal (seminal vesicle) vesicles, and bulbourethral glands
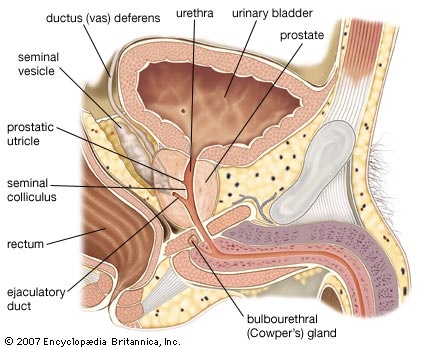 These structures provide secretions to form the bulk of the seminal fluid of an ejaculate. The prostate gland is in the lesser or true pelvis, centred behind the lower part of the pubic arch. It lies in front of the rectum. The prostate is shaped roughly like an inverted pyramid; its base is directed upward and is immediately continuous with the neck of the urinary bladder. The urethra traverses its substance. The two ejaculatory ducts enter the prostate near the upper border of its posterior surface. The prostate is of a firm consistency, surrounded by a capsule of fibrous tissue and smooth muscle. It measures about 4 cm across, 3 cm in height, and 2 cm front to back (about 1.6 by 1.2 by 0.8 inch) and consists of glandular tissue contained in a muscular framework. It is imperfectly divided into three lobes. Two lobes at the side form the main mass and are continuous behind the urethra. In front of the urethra they are connected by an isthmus of fibromuscular tissue devoid of glands. The third, or median, lobe is smaller and variable in size and may lack glandular tissue. There are three clinically significant concentric zones of prostatic glandular tissue about the urethra. A group of short glands that are closest to the urethra and discharge mucus into its channel are subject to simple enlargement. Outside these is a ring of submucosal glands (glands from which the mucosal glands develop), and farther out is a large outer zone of long branched glands, composing the bulk of the glandular tissue. prostate cancer is almost exclusively confined to the outer zone. The glands of the outer zone are lined by tall columnar cells that secrete prostatic fluid under the influence of androgens from the testis. The fluid is thin, milky, and slightly acidic.
These structures provide secretions to form the bulk of the seminal fluid of an ejaculate. The prostate gland is in the lesser or true pelvis, centred behind the lower part of the pubic arch. It lies in front of the rectum. The prostate is shaped roughly like an inverted pyramid; its base is directed upward and is immediately continuous with the neck of the urinary bladder. The urethra traverses its substance. The two ejaculatory ducts enter the prostate near the upper border of its posterior surface. The prostate is of a firm consistency, surrounded by a capsule of fibrous tissue and smooth muscle. It measures about 4 cm across, 3 cm in height, and 2 cm front to back (about 1.6 by 1.2 by 0.8 inch) and consists of glandular tissue contained in a muscular framework. It is imperfectly divided into three lobes. Two lobes at the side form the main mass and are continuous behind the urethra. In front of the urethra they are connected by an isthmus of fibromuscular tissue devoid of glands. The third, or median, lobe is smaller and variable in size and may lack glandular tissue. There are three clinically significant concentric zones of prostatic glandular tissue about the urethra. A group of short glands that are closest to the urethra and discharge mucus into its channel are subject to simple enlargement. Outside these is a ring of submucosal glands (glands from which the mucosal glands develop), and farther out is a large outer zone of long branched glands, composing the bulk of the glandular tissue. prostate cancer is almost exclusively confined to the outer zone. The glands of the outer zone are lined by tall columnar cells that secrete prostatic fluid under the influence of androgens from the testis. The fluid is thin, milky, and slightly acidic.The seminal vesicles (seminal vesicle) are two structures, about 5 cm (2 inches) in length, lying between the rectum and the base of the bladder. Their secretions form the bulk of semen. Essentially, each vesicle consists of a much-coiled tube with numerous diverticula or outpouches that extend from the main tube, the whole being held together by connective tissue. At its lower end the tube is constricted to form a straight duct or tube that joins with the corresponding ductus deferens to form the ejaculatory duct. The vesicles are close together in their lower parts, but they are separated above where they lie close to the deferent ducts. The seminal vesicles have longitudinal and circular layers of smooth muscle, and their cavities are lined with mucous membrane, which is the source of the secretions of the organs. These secretions are ejected by muscular contractions during ejaculation. The activity of the vesicles is dependent on the production of the hormone androgen by the testes. The secretion is thick, sticky, and yellowish; it contains the sugar fructose and is slightly alkaline.
The bulbourethral glands (bulbourethral gland), often called Cowper glands, are pea-shaped glands that are located beneath the prostate gland at the beginning of the internal portion of the penis. The glands, which measure only about 1 cm (0.4 inch) in diameter, have slender ducts that run forward and toward the centre to open on the floor of the spongy portion of the urethra. They are composed of a network of small tubes, or tubules, and saclike structures; between the tubules are fibres of muscle and elastic tissue that give the glands muscular support. Cells within the tubules and sacs contain droplets of mucus, a thick protein compound. The fluid excreted by these glands is clear and thick and acts as a lubricant; it is also thought to function as a flushing agent that washes out the urethra before the semen is ejaculated; it may also help to make the semen less watery and to provide a suitable living environment for the sperm.
Ejaculatory ducts
The two ejaculatory ducts lie on each side of the midline and are formed by the union of the duct of the seminal vesicle, which contributes secretions to the semen, with the end of the ductus deferens at the base of the prostate. Each duct is about 2 cm (about 0.8 inch) long and passes between a lateral and the median lobe of the prostate to reach the floor of the prostatic urethra. This part of the urethra has on its floor (or posterior wall) a longitudinal ridge called the urethral crest. On each side is a depression, the prostatic sinus, into which open the prostatic ducts. In the middle of the urethral crest is a small elevation, the colliculus seminalis, on which the opening of the prostatic utricle is found. The prostatic utricle is a short diverticulum or pouch lined by mucous membrane; it may correspond to the vagina or uterus in the female. The small openings of the ejaculatory ducts lie on each side of or just within the opening of the prostatic utricle. The ejaculatory ducts are thin-walled and lined by columnar cells.
The female reproductive system
The female gonads, or sexual glands, are the ovaries; they are the source of ova (ovum) (eggs) and of the female sex hormones estrogens (estrogen) and progestogens (progesterone). The fallopian, or uterine, tubes conduct ova to the uterus, which lies within the lesser or true pelvis. The uterus connects through the cervical canal with the vagina. The vagina opens into the vestibule about which lie the external genitalia, collectively known as the vulva.External genitalia
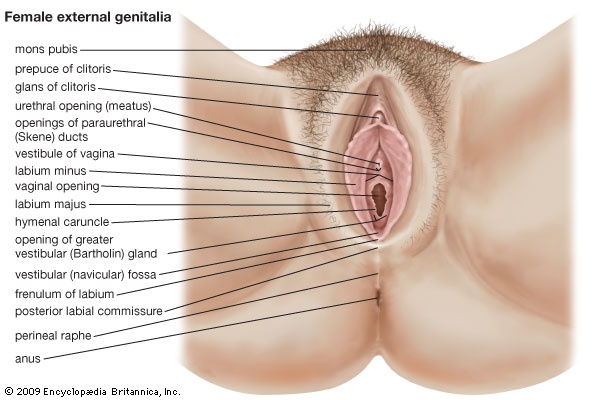 The female external genitalia include the structures placed about the entrance to the vagina and external to the hymen, the membrane across the entrance to the vagina. They are the mons pubis (also called the mons veneris), the labia majora and minora, the clitoris, the vestibule of the vagina, the bulb of the vestibule, and the greater vestibular glands.
The female external genitalia include the structures placed about the entrance to the vagina and external to the hymen, the membrane across the entrance to the vagina. They are the mons pubis (also called the mons veneris), the labia majora and minora, the clitoris, the vestibule of the vagina, the bulb of the vestibule, and the greater vestibular glands.The mons pubis is the rounded eminence, made by fatty tissue beneath the skin, lying in front of the pubic symphysis. A few fine hairs may be present in childhood; later, at puberty, they become coarser and more numerous. The upper limit of the hairy region is horizontal across the lower abdomen.
The labia majora are two marked folds of skin that extend from the mons pubis downward and backward to merge with the skin of the perineum. They form the lateral boundaries of the vulval or pudendal cleft, which receives the openings of the vagina and the urethra. The outer surface of each labium is pigmented and hairy; the inner surface is smooth but possesses sebaceous glands (sebaceous gland). The labia majora contain fat and loose connective tissue and sweat glands. They correspond to the scrotum in the male and contain tissue resembling the dartos muscle. The round ligament (see below The uterus (reproductive system, human)) ends in the tissue of the labium. The labia minora are two small folds of skin, lacking fatty tissue, that extend backward on each side of the opening into the vagina. They lie inside the labia majora and are some 4 cm (about 1.5 inches) in length. In front, an upper portion of each labium minus passes over the clitoris—the structure in the female corresponding to the penis (excluding the urethra) in the male—to form a fold, the prepuce of the clitoris, and a lower portion passes beneath the clitoris to form its frenulum. The two labia minora are joined at the back across the midline by a fold that becomes stretched at childbirth. The labia minora lack hairs but possess sebaceous and sweat glands.
The clitoris is a small erectile structure composed of two corpora cavernosa separated by a partition. Partially concealed beneath the forward ends of the labia minora, it possesses a sensitive tip of spongy erectile tissue, the glans clitoridis. The external opening of the urethra is some 2.5 cm (about 1 inch) behind the clitoris and immediately in front of the vaginal opening.
The vestibule of the vagina is the cleft between the labia minora into which the urethra and vagina open. The hymen vaginae lies at the opening of the vagina: it is a thin fold of mucous membrane that varies in shape. After rupture of the hymen, the small rounded elevations that remain are known as the carunculae hymenales. The bulb of the vestibule, corresponding to the bulb of the penis, is two elongated masses of erectile tissue that lie one on each side of the vaginal opening. At their posterior ends lie the greater vestibular glands, small mucous glands that open by a duct in the groove between the hymen and each labium minus. They correspond to the bulbourethral glands of the male.
The blood supply and nerve supply of the female external genital organs are similar to those supplying corresponding structures in the male.
Internal structures
The vagina
The vagina (the word means “sheath”) is the canal that extends from the cervix (uterine cervix) (outer end) of the uterus within the lesser pelvis down to the vestibule between the labia minora. The orifice of the vagina is guarded by the hymen. The vagina lies behind the bladder and urethra and in front of the rectum and anal canal (anus). Its walls are collapsed; the anterior wall is some 7.5 cm (3 inches) in length, whereas the posterior wall is about 1.5 cm (0.6 inch) longer. The vagina is directed obliquely upward and backward. The axis of the vagina forms an angle of over 90° with that of the uterus. This angle varies considerably depending on conditions in the bladder, in the rectum, and during pregnancy. The cervix of the uterus projects for a short distance into the vagina and is normally pressed against its posterior wall. There are, therefore, recesses in the vagina at the back, on each side, and at the front of the cervix. These are known as the posterior fornix (behind the cervix and the largest), the lateral fornices (at the sides), and the anterior fornix (at the front of the cervix). The position of the uterus in relation to the vagina is described further in the section on the uterus.The upper part of the posterior wall of the vagina is covered by peritoneum or membrane that is folded back onto the rectum to form the recto-uterine pouch. The lower part of the posterior vaginal wall is separated from the anal canal by a mass of tissue known as the perineal body.
The vagina has a mucous membrane and an outer smooth muscle coat closely attached to it. The mucous membrane has a longitudinal ridge in the midline of both the anterior and posterior walls. The ridges are known as the columns of the vagina; many rugae, or folds, extend from them to each side. The furrows between the rugae are more marked on the posterior wall and become especially pronounced before the birth of a child. The membrane undergoes little change during the menstrual cycle (menstruation) (except in its content of glycogen, a complex starchlike carbohydrate); this is in contradistinction to the situation in many mammals in which marked exfoliation (shedding of the surface cells) can occur. No glands are present in the vaginal lining, and mucus present has been secreted by the glands in the cervical canal of the uterus. The smooth muscle coat consists of an outer longitudinal layer and a less developed inner circular layer. The lower part of the vagina is surrounded by the bulbospongiosus muscle, a striped muscle attached to the perineal body.
The blood supply to the vagina is derived from several adjacent vessels, there being a vaginal artery from the internal iliac artery and also vaginal branches from the uterine, middle rectal, and internal pudendal arteries, all branches of the internal iliac artery. The nerve supply to the lower part of the vagina is from the pudendal nerve and from the inferior hypogastric and uterovaginal plexuses.
The uterus
Uterine structure
The uterus, or womb, is shaped like an inverted pear. It is a hollow, muscular organ with thick walls, and it has a glandular lining called the endometrium. In an adult the uterus is 7.5 cm (3 inches) long, 5 cm (2 inches) in width, and 2.5 cm (1 inch) thick, but it enlarges to four to five times this size in pregnancy. The narrower, lower end is called the cervix (uterine cervix); this projects into the vagina. The cervix is made of fibrous connective tissue and is of a firmer consistency than the body of the uterus. The two fallopian tubes (fallopian tube) enter the uterus at opposite sides, near its top. The part of the uterus above the entrances of the tubes is called the fundus; the part below is termed the body. The body narrows toward the cervix, and a slight external constriction marks the juncture between the body and the cervix.The uterus does not lie in line with the vagina but is usually turned forward (anteverted) to form approximately a right angle with it. The position of the uterus is affected by the amount of distension in the urinary bladder and in the rectum. Enlargement of the uterus in pregnancy causes it to rise up into the abdominal cavity, so that there is closer alignment with the vagina. The nonpregnant uterus also curves gently forward; it is said to be anteflexed. The uterus is supported and held in position by the other pelvic organs, by the muscular floor or diaphragm of the pelvis, by certain fibrous ligaments, and by folds of peritoneum. Among the supporting ligaments are two double-layered broad ligaments, each of which contains a fallopian tube along its upper free border and a round ligament, corresponding to the gubernaculum testis of the male, between its layers. Two ligaments—the cardinal (Mackenrodt) ligaments—at each side of the cervix are also important in maintaining the position of the uterus.
The cavity of the uterus is remarkably small in comparison with the size of the organ. Except during pregnancy, the cavity is flattened, with front and rear walls touching, and is triangular. The triangle is inverted, with its base at the top, between the openings of the two fallopian tubes, and with its apex at the isthmus of the uterus, the opening into the cervix. The canal of the cervix is flattened from front to back and is somewhat larger in its middle part. It is traversed by two longitudinal ridges and has oblique folds stretching from each ridge in an arrangement like the branches of a tree. The cervical canal is 2.5 cm (about 1 inch) in length; its opening into the vagina is called the external os of the uterus. The external os is small, almost circular, and often depressed. After childbirth, the external os becomes bounded by lips in front and in back and is thus more slitlike. The cervical canal is lined by a mucous membrane containing numerous glands that secrete a clear, alkaline mucus. The upper part of this lining undergoes cyclical changes resembling, but not as marked as, those occurring in the body of the uterus. Numerous small cysts (nabothian cysts) are found in the cervical mucous membrane. It is from this region that cervical smears are taken in order to detect early changes indicative of cancer (cervical cancer).
The uterus is composed of three layers of tissue. On the outside is a serous coat of peritoneum (a membrane exuding a fluid like blood minus its cells and the clotting factor fibrinogen), which partially covers the organ. In front it covers only the body of the cervix; behind it covers the body and the part of the cervix that is above the vagina and is prolonged onto the posterior vaginal wall; from there it is folded back to the rectum. At the side the peritoneal layers stretch from the margin of the uterus to each side wall of the pelvis, forming the two broad ligaments of the uterus.
The middle layer of tissue (myometrium) is muscular and comprises the greater part of the bulk of the organ. It is very firm and consists of densely packed, unstriped, smooth muscle fibres. Blood vessels, lymph vessels, and nerves are also present. The muscle is more or less arranged in three layers of fibres running in different directions. The outermost fibres are arranged longitudinally. Those of the middle layer run in all directions without any orderly arrangement; this layer is the thickest. The innermost fibres are longitudinal and circular in their arrangement.
The innermost layer of tissue in the uterus is the mucous membrane, or endometrium. It lines the uterine cavity as far as the isthmus of the uterus, where it becomes continuous with the lining of the cervical canal. The endometrium contains numerous uterine glands that open into the uterine cavity and are embedded in the cellular framework or stroma of the endometrium. Numerous blood vessels and lymphatic spaces are also present. The appearances of the endometrium vary considerably at the different stages in reproductive life. It begins to reach full development at puberty and thereafter exhibits dramatic changes during each menstrual cycle (menstruation). It undergoes further changes before, during, and after pregnancy, during the menopause, and in old age. These changes are for the most part hormonally induced and controlled by the activity of the ovaries (ovary).
The endometrium in the menstrual cycle
 To understand the nature of the changes in the endometrium during each menstrual cycle it is usual to consider the endometrium to be composed of three layers. They blend imperceptibly but are functionally distinct: the inner two layers are shed at menstruation, and the outer or basal layer remains in position against the innermost layer of the myometrium. The three layers are called, respectively, the stratum compactum, the stratum spongiosum, and the stratum basale epidermidis. The stratum compactum is nearest to the uterine cavity and contains the lining cells and the necks of the uterine glands; its stroma is relatively dense. Superficial blood vessels lie beneath the lining cells. The stratum spongiosum is the large middle layer. It contains the main portions of uterine glands and accompanying blood vessels; the stromal cells are more loosely arranged and larger than in the stratum compactum. The stratum basale epidermidis lies against the uterine muscle; it contains blood vessels and the bases of the uterine glands. Its stroma remains relatively unaltered during the menstrual cycle.
To understand the nature of the changes in the endometrium during each menstrual cycle it is usual to consider the endometrium to be composed of three layers. They blend imperceptibly but are functionally distinct: the inner two layers are shed at menstruation, and the outer or basal layer remains in position against the innermost layer of the myometrium. The three layers are called, respectively, the stratum compactum, the stratum spongiosum, and the stratum basale epidermidis. The stratum compactum is nearest to the uterine cavity and contains the lining cells and the necks of the uterine glands; its stroma is relatively dense. Superficial blood vessels lie beneath the lining cells. The stratum spongiosum is the large middle layer. It contains the main portions of uterine glands and accompanying blood vessels; the stromal cells are more loosely arranged and larger than in the stratum compactum. The stratum basale epidermidis lies against the uterine muscle; it contains blood vessels and the bases of the uterine glands. Its stroma remains relatively unaltered during the menstrual cycle.The menstrual cycle extends over a period of about 28 days (normal range 21–34 days), from the first day of one menstrual flow to the first day of the next. It reflects the cycle of changes occurring in the ovary, which is itself under the control of the anterior lobe of the pituitary gland. The menstrual cycle is divided into four phases: menstrual, postmenstrual, proliferative, and secretory.
The secretory phase reaches its climax about a week after ovulation. Ovulation occurs in midcycle, about 14 days before the onset of the next menstrual flow. The endometrium has been prepared and has been stimulated to a state of active secretion for the reception of a fertilized ovum. The stage has been set for the attachment of the blastocyst, derived from a fertilized ovum, to the endometrium and for its subsequent embedding. This process is called implantation; its success depends on the satisfactory preparation of the endometrium in both the proliferative and secretory phases. When implantation occurs, a hormone from certain cells of the blastocyst causes prolongation of the corpus luteum and its continued activity. This causes suppression of menstruation and results in the maintenance of the endometrium and its further stimulation by progesterone, with consequent increased thickening. The endometrium of early pregnancy is known as the decidua.
In a cycle in which fertilization of the ovum has not taken place, the secretory phase terminates in menstruation.
The endometrium needs to be in a certain state of preparedness before implantation can occur. When this stage has been passed, menstruation occurs. Repair then reestablishes an endometrium capable of being stimulated again to the critical stage when implantation can occur.
Blood supply and innervation
The uterus is supplied with blood by the two uterine arteries, which are branches of the internal iliac arteries, and by ovarian arteries, which connect with the ends of the uterine arteries and send branches to supply the uterus. The nerves to the uterus include the sympathetic nerve fibres, which produce contraction of uterine muscle and constriction of vessels, and parasympathetic (sacral) fibres, which inhibit muscle activity and cause dilation of blood vessels.
The fallopian tubes
 The fallopian (fallopian tube), or uterine, tubes carry ova from the ovaries to the cavity of the uterus. Each opens into the abdominal cavity near an ovary at one end and into the uterus at the other. Three sections of the tubes are distinguished: the funnel-shaped outer end, or infundibulum; the expanded and thin-walled intermediate portion, or ampulla; and the cordlike portion, the isthmus, that opens into the uterus. The infundibulum is fringed with irregular projections called fimbriae. One fimbria, somewhat larger than the others, is usually attached to the ovary. The opening into the abdomen is at the bottom of the infundibulum and is small. fertilization of the ovum usually occurs in the ampulla of the tube. Normally the fertilized egg is transported to the uterus, but occasionally it may adhere to the tube and start developing as an ectopic pregnancy, or tubal pregnancy. The tube is unable to support this pregnancy, and the conceptus may be extruded through the abdominal opening or may cause rupture of the tube, with ensuing hemorrhage.
The fallopian (fallopian tube), or uterine, tubes carry ova from the ovaries to the cavity of the uterus. Each opens into the abdominal cavity near an ovary at one end and into the uterus at the other. Three sections of the tubes are distinguished: the funnel-shaped outer end, or infundibulum; the expanded and thin-walled intermediate portion, or ampulla; and the cordlike portion, the isthmus, that opens into the uterus. The infundibulum is fringed with irregular projections called fimbriae. One fimbria, somewhat larger than the others, is usually attached to the ovary. The opening into the abdomen is at the bottom of the infundibulum and is small. fertilization of the ovum usually occurs in the ampulla of the tube. Normally the fertilized egg is transported to the uterus, but occasionally it may adhere to the tube and start developing as an ectopic pregnancy, or tubal pregnancy. The tube is unable to support this pregnancy, and the conceptus may be extruded through the abdominal opening or may cause rupture of the tube, with ensuing hemorrhage.The fallopian tube is covered by peritoneum except on its border next to the broad ligament. There are inner circular and outer longitudinal layers of smooth muscle fibres continuous with those of the uterus. The inner lining has numerous longitudinal folds that are covered with ciliated columnar and secretory cells. Muscular contraction, movement of the hairlike cilia, and the passage of the watery secretions all probably assist in the transport of sperm to the ampulla and of a fertilized ovum toward the uterus.
The ovaries (ovary)
Ovarian structure
The female gonads, or primary sex organs, corresponding to the testes (testis) in a male, are the two ovaries (ovary). Each is suspended by a mesentery, or fold of membrane, from the back layer of the broad ligament of the uterus. In a woman who has not been pregnant, the almond-shaped ovary lies in a vertical position against a depression, the ovarian fossa, on the side wall of the lesser pelvis. This relationship is altered during and after pregnancy. Each ovary is somewhat over 2.5 cm (1 inch) in length, 1.25 cm (0.5 inch) across, and slightly less in thickness, but the size varies much with age and with state of activity.The mesentery of the ovary helps to keep it in position, and within this membrane lie the ovarian artery and vein, lymphatic vessels, and nerve fibres. The fallopian tube arches over the ovary and curves downward on its inner or medial surface.
Except at its hilum, the point where blood vessels and the nerve enter the ovary and where the mesentery is attached, the surface of the ovary is smooth and is covered by cubical cells. Beneath the surface, the substance of the ovary is divided into an outer portion, the cortex, and an inner portion, or medulla. The outermost part of the cortex, immediately beneath the outer covering, forms a thin connective tissue zone, the tunica albuginea. The rest of the cortex consists of stromal or framework cells, contained in a fine network of fibres, and also the follicles and corpora lutea.
The ovarian follicles, sometimes called graafian follicles, are rounded enclosures for the developing ova in the cortex near the surface of the ovary. At birth and in childhood they are present as numerous primary or undeveloped ovarian follicles. Each contains a primitive ovum, or oocyte, and each is covered by a single layer of flattened cells. As many as 700,000 primary follicles are contained in the two ovaries of a young female. Most of these degenerate before or after puberty.
ovulation
 During the onset of puberty and thereafter until menopause (except during pregnancy), there is a cyclic development of one or more follicles each month into a mature follicle. The covering layer of the primary follicle thickens and can be differentiated into an inner membrana granulosa and an outer vascularized theca interna. The cells of these layers (mostly the theca interna) produce estrogenic steroid hormones that exert their effects on the endometrium of the uterus and on other tissues. The maintenance and growth of the follicle to maturity is brought about by a follicle-stimulating hormone (FSH) from the anterior lobe of the pituitary gland. Another hormone, called luteinizing hormone (LH), from the anterior lobe, assists FSH to cause the maturing, now fluid-filled follicle to secrete estrogens (estrogen). LH also causes a ripe follicle (1.0–1.5 cm 【0.4–0.6 inch】 in diameter) to rupture, causing the liberation of the oocyte into the peritoneal cavity and thence into the fallopian tube. This liberation of the oocyte is called ovulation; it occurs at about the midpoint of the reproductive cycle, on the 13th or 14th day of a 28-day cycle as measured from the first day of the menstrual flow.
During the onset of puberty and thereafter until menopause (except during pregnancy), there is a cyclic development of one or more follicles each month into a mature follicle. The covering layer of the primary follicle thickens and can be differentiated into an inner membrana granulosa and an outer vascularized theca interna. The cells of these layers (mostly the theca interna) produce estrogenic steroid hormones that exert their effects on the endometrium of the uterus and on other tissues. The maintenance and growth of the follicle to maturity is brought about by a follicle-stimulating hormone (FSH) from the anterior lobe of the pituitary gland. Another hormone, called luteinizing hormone (LH), from the anterior lobe, assists FSH to cause the maturing, now fluid-filled follicle to secrete estrogens (estrogen). LH also causes a ripe follicle (1.0–1.5 cm 【0.4–0.6 inch】 in diameter) to rupture, causing the liberation of the oocyte into the peritoneal cavity and thence into the fallopian tube. This liberation of the oocyte is called ovulation; it occurs at about the midpoint of the reproductive cycle, on the 13th or 14th day of a 28-day cycle as measured from the first day of the menstrual flow.After ovulation the ruptured follicle collapses because of loss of its follicular fluid and rapidly becomes transformed into a soft, well-vascularized glandular structure known as the corpus luteum(“yellow body”). The corpus luteum develops rapidly, becomes vascularized after about four days, and is fully established by nine days. The gland produces the steroid hormone progesterone and some estrogens. Its activity is both stimulated and maintained by luteinizing hormone. Progesterone stimulates glandular proliferation and secretion in an endometrium primed by estrogens.
While the ovarian follicle matures, the primary oocyte divides into a secondary oocyte and a small rudimentary ovum called the first polar body. This occurs at about the time when the follicle develops its cavity; the oocyte also gains a translucent acellular covering, or envelope, the zona pellucida. The secondary oocyte is liberated at ovulation; it is 120–140 micrometres in diameter and is surrounded by the zona pellucida and a few layers of cells known as the corona radiata. The final maturation of the oocyte, with the formation of the rudimentary ovum called the second polar body, occurs at the time of fertilization.
If fertilization does not occur, then the life of the corpus luteum is limited to about 14 days. Degeneration of the gland starts toward the end of this period, and menstruation occurs. The corpus luteum shrinks, fibrous tissue is formed, and it is converted into a scarlike structure called a corpus albicans, which persists for a few months.
Should fertilization occur and be followed by implantation of the blastocyst, hormones (particularly human chorionic gonadotropin) are produced by cells of the blastocyst to prolong the life of the corpus luteum. It persists in an active state for at least the first two months of pregnancy, until the placental tissue has taken over its hormone-producing function. The corpus luteum of pregnancy then also retrogresses, becoming a fibrous scar by the time of parturition.
Blood supply and innervation
The ovarian arteries arise from the front of the aorta in a manner similar to the testicular arteries, but at the brim of the lesser pelvis they turn down into the pelvic cavity. Passing in the suspensory ligament of the ovary, each artery reaches the broad ligament below the fallopian tube and then passes into the mesovarium to divide into branches distributed to the ovary. One branch continues in the broad ligament to anastomose with the uterine artery. The ovarian veins emerge from each ovary as a network that eventually becomes a single vein; the terminations are similar to those of the testicular veins. The nerves are derived from the ovarian nerve network on the ovarian artery.
Additional Reading
The characteristics of human reproduction and reproductive organs are reviewed in Robert H. Williams (ed.), Textbook of Endocrinology, 6th ed. (1981), covering in detail the endocrine aspects of the male and female reproductive systems; Richard E. Jones, Human Reproductive Biology (1991); Ralph M. Wynn and William P. Jollie (eds.), Biology of the Uterus, 2nd ed. (1989), a useful summary of modern research on many aspects of uterine anatomy and physiology; Ethel Sloane, Biology of Women, 3rd ed. (1993); and Claude Gompel and Steven G. Silverberg, Pathology in Gynecology and Obstetrics, 4th ed. (1994), which contains a chapter on each organ of the female genital tract and one on the breast. Reproductive technologies are explained in Lori B. Andrews, New Conceptions (1984); Arthur L. Wisot and David R. Meldrum, New Options for Fertility (1990); and Sherman J. Silber, How to Get Pregnant with the New Technology (1991). Ed.
- Cetinje
- Cetshwayo
- cetyl alcohol
- Ceuta
- Ceva, Giovanni
- Ceva, Tommaso
- Cevdet Paşa, Ahmed
- Ceyhan River
- Ceylon ironwood
- Ceṭṭi
- C.F. Beyers
- CG-4
- CGI
- C.G. Seligman
- C.H.
- Chaadayev, Pyotr Yakovlevich
- Chaban-Delmas, Jacques
- sieur de Crébillon, Claude-Prosper Jolyot
- sieur de Crébillon, Prosper Jolyot
- sieur de Fontenelle, Bernard Le Bovier
- Sieur De La Mauvissière Castelnau, Michel de
- Sieur de Roberval, Jean-François de La Rocque
- sieur de Saint-Amant, Marc-Antoine Girard
- sieur d'Esnambuc Bélain, Pierre
- Sieur DuLhut, Daniel Greysolon