Australopithecus
paleontology
Introduction
Latin“southern ape”
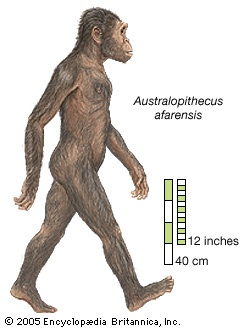
 group of extinct creatures closely related to, if not actually ancestors of, modern human beings and known from a series of fossils found at numerous sites in eastern, central, and southern Africa. The various species of Australopithecus lived during the Pliocene Epoch, which lasted from 5.3 to 1.8 million years ago (mya). As characterized by the fossil evidence, they bore a combination of human- and apelike traits. Like humans, they were bipedal (that is, they walked on two legs), but, like apes, they had small brains. Their canine teeth (tooth) were small like those of humans, but their cheek teeth were large. The genus name meaning “southern ape” refers to the first fossils found, which were discovered in South Africa. Perhaps the most famous specimen of Australopithecus is “Lucy,” a remarkably preserved fossilized skeleton from Ethiopia that has been dated to 3.2 mya.
group of extinct creatures closely related to, if not actually ancestors of, modern human beings and known from a series of fossils found at numerous sites in eastern, central, and southern Africa. The various species of Australopithecus lived during the Pliocene Epoch, which lasted from 5.3 to 1.8 million years ago (mya). As characterized by the fossil evidence, they bore a combination of human- and apelike traits. Like humans, they were bipedal (that is, they walked on two legs), but, like apes, they had small brains. Their canine teeth (tooth) were small like those of humans, but their cheek teeth were large. The genus name meaning “southern ape” refers to the first fossils found, which were discovered in South Africa. Perhaps the most famous specimen of Australopithecus is “Lucy,” a remarkably preserved fossilized skeleton from Ethiopia that has been dated to 3.2 mya. The general term australopith (or australopithecine) is used informally to refer not only to members of the genus Australopithecus but also to other humanlike primates (primate) that lived in Africa between 6 and 1.2 mya. Other australopiths include Sahelanthropus tchadensis (7–6 mya), Orrorin tugenensis (6 mya), Ardipithecus kadabba and Ardipithecus ramidus (5.8–4.4 mya), Kenyanthropus platyops (3.5–3.2 mya), and three species of Paranthropus (2.3–1.2 mya). Remains older than 6 million years are widely regarded as those of fossil apes. Undisputed evidence of the genus Homo—the genus that includes modern human beings—does not appear until about 1.8 mya, in the form of Homo ergaster, also called H. erectus (Homo erectus) (“upright man”). The remains of H. habilis (Homo habilis) (“handy man”) and H. rudolfensis are between 2.5 and 1.5 million years old, but these are difficult to differentiate from those of Australopithecus, and the identity of some of these remains is debated.
The general term australopith (or australopithecine) is used informally to refer not only to members of the genus Australopithecus but also to other humanlike primates (primate) that lived in Africa between 6 and 1.2 mya. Other australopiths include Sahelanthropus tchadensis (7–6 mya), Orrorin tugenensis (6 mya), Ardipithecus kadabba and Ardipithecus ramidus (5.8–4.4 mya), Kenyanthropus platyops (3.5–3.2 mya), and three species of Paranthropus (2.3–1.2 mya). Remains older than 6 million years are widely regarded as those of fossil apes. Undisputed evidence of the genus Homo—the genus that includes modern human beings—does not appear until about 1.8 mya, in the form of Homo ergaster, also called H. erectus (Homo erectus) (“upright man”). The remains of H. habilis (Homo habilis) (“handy man”) and H. rudolfensis are between 2.5 and 1.5 million years old, but these are difficult to differentiate from those of Australopithecus, and the identity of some of these remains is debated.The australopiths
Early species and Australopithecus anamensis
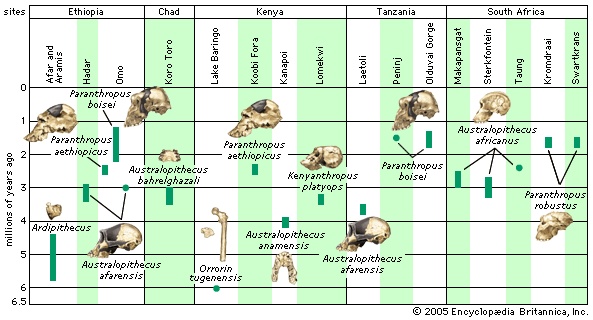 Identifying the earliest member of the human tribe (Hominini) is difficult because the predecessors of modern humans are increasingly apelike as the fossil record is followed back through time. They resemble what would be expected in the common ancestor of humans and apes in that they possess a mix of human and ape traits. For example, the earliest species, S. tchadensis, is humanlike in having small canine teeth and a face that does not project very far. However, in most other respects, including brain size, it is apelike. Whether it walked upright is not known because only a single skull, jaw fragments, and teeth have been found. Bipedalism may have been established in the six-million-year-old Orrorin tugenensis, an australopith found in the Tugen Hills near Lake Baringo (Baringo, Lake) in central Kenya. In 2001 these fossils were described as the earliest known hominin. O. tugenensis is primitive in most if not all of its body except for femurs (thighbones) that appear to share traits of bipedalism with modern humans. Like later hominins, it has teeth with thick molar enamel, but, unlike humans, it has distinctively apelike canine and premolar teeth. The case for its hominin status rests on the humanlike features of the femur. According to its discoverers, features of the thighbone implying bipedalism include its overall proportions, the internal structure of the knee, and a groove on the bone for a muscle used in upright walking (the obturator externus).
Identifying the earliest member of the human tribe (Hominini) is difficult because the predecessors of modern humans are increasingly apelike as the fossil record is followed back through time. They resemble what would be expected in the common ancestor of humans and apes in that they possess a mix of human and ape traits. For example, the earliest species, S. tchadensis, is humanlike in having small canine teeth and a face that does not project very far. However, in most other respects, including brain size, it is apelike. Whether it walked upright is not known because only a single skull, jaw fragments, and teeth have been found. Bipedalism may have been established in the six-million-year-old Orrorin tugenensis, an australopith found in the Tugen Hills near Lake Baringo (Baringo, Lake) in central Kenya. In 2001 these fossils were described as the earliest known hominin. O. tugenensis is primitive in most if not all of its body except for femurs (thighbones) that appear to share traits of bipedalism with modern humans. Like later hominins, it has teeth with thick molar enamel, but, unlike humans, it has distinctively apelike canine and premolar teeth. The case for its hominin status rests on the humanlike features of the femur. According to its discoverers, features of the thighbone implying bipedalism include its overall proportions, the internal structure of the knee, and a groove on the bone for a muscle used in upright walking (the obturator externus).Another candidate for the earliest australopith is Ardipithecus (5.8–4.4 mya), found in 1992 at Aramis in the Afar region of Ethiopia. It too is primitive compared with later hominins, though it does share a few evolutionary (evolution) novelties associated with hominins. Its cranial base is short like that of hominins, and the upper canines are shaped somewhat like those of later species. A well-preserved toe bone shows the characteristically bipedal feature of a base designed for hyperextension while walking. Interestingly, Ardipithecus fossils have been found in association with animals usually found in closed woodland habitats rather than open grasslands.
The earliest member of the genus Australopithecus is A. anamensis, discovered in 1994 by a team led by Meave Leakey (Leakey, Meave G.) at Kanapoi and Allia Bay in northern Kenya. The fossils date to 4.2–3.9 mya, and, like Ardipithecus, A. anamensis is associated with woodland animals and a few grassland species as well. It is quite primitive with a strongly protruding lower face, but at the same time it has certain dental features not seen in Ardipithecus ramidus; most conspicuous is a thickening of tooth enamel that becomes characteristic of all later hominins. In addition, the ankle and knee are specialized for upright walking. Other skeletal features are very much like those of later hominins.
 In 1998 Leakey's team also discovered Kenyanthropus platyops (3.5–3.2 mya) at Lomekwi on the western shore of Lake Turkana (Turkana) in northern Kenya. It too is associated with woodland fauna. It possesses some primitive skull features but shares with early Homo a flat and tall face. Though it overlaps in time with A. afarensis (described below), it appears to be quite distinctive in its morphology and in some respects more primitive. In other respects it resembles much later hominins, particularly H. rudolfensis, in having a relatively flat face and small molars. These traits are related to chewing and thus may be related to diet. It is therefore possible that the resemblances between H. rudolfensis and K. platyops are the result of independent adaptations to similar situations. It is equally possible that the resemblances may imply an evolutionary link between the two.
In 1998 Leakey's team also discovered Kenyanthropus platyops (3.5–3.2 mya) at Lomekwi on the western shore of Lake Turkana (Turkana) in northern Kenya. It too is associated with woodland fauna. It possesses some primitive skull features but shares with early Homo a flat and tall face. Though it overlaps in time with A. afarensis (described below), it appears to be quite distinctive in its morphology and in some respects more primitive. In other respects it resembles much later hominins, particularly H. rudolfensis, in having a relatively flat face and small molars. These traits are related to chewing and thus may be related to diet. It is therefore possible that the resemblances between H. rudolfensis and K. platyops are the result of independent adaptations to similar situations. It is equally possible that the resemblances may imply an evolutionary link between the two.Australopithecus afarensis and A. garhi
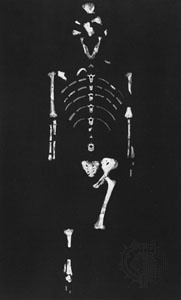
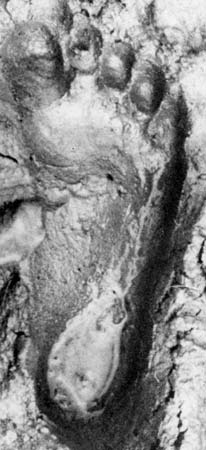 The best-known member of Australopithecus is A. afarensis, discovered in deposits in East Africa and ranging in age from 3.8 to 2.9 million years old. Part of the earliest sample derives from the northern Tanzanian site of Laetoli, where specimens range from 3.8 to 3.5 mya and include footprints preserved in volcanic ash dating to 3.6–3.5 mya. These footprints are remarkably similar to those of modern humans in key details, including a forward-pointing big toe, relatively short lateral toes, and arched feet. The main fossil sample of this species comes from Hadar, a site in the Afar region of Ethiopia. Specimens here include a 40-percent-complete skeleton of an adult female (“Lucy”) and the remains of at least nine adults and four juveniles buried together at the same time (the “First Family”). The animal fossils found in association with A. afarensis imply a habitat of woodland with patches of grassland.
The best-known member of Australopithecus is A. afarensis, discovered in deposits in East Africa and ranging in age from 3.8 to 2.9 million years old. Part of the earliest sample derives from the northern Tanzanian site of Laetoli, where specimens range from 3.8 to 3.5 mya and include footprints preserved in volcanic ash dating to 3.6–3.5 mya. These footprints are remarkably similar to those of modern humans in key details, including a forward-pointing big toe, relatively short lateral toes, and arched feet. The main fossil sample of this species comes from Hadar, a site in the Afar region of Ethiopia. Specimens here include a 40-percent-complete skeleton of an adult female (“Lucy”) and the remains of at least nine adults and four juveniles buried together at the same time (the “First Family”). The animal fossils found in association with A. afarensis imply a habitat of woodland with patches of grassland.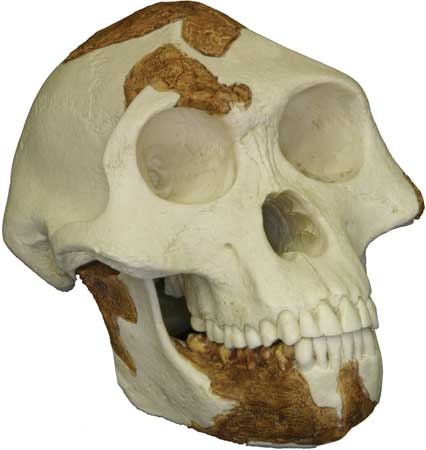 The morphology of A. afarensis is a mosaic of primitive features and evolutionary developments shared by later hominins. Its skull is primitive in having a crest and a strongly projecting (prognathic) lower face. The brain was about one-third the size of a modern human's. The dentition is also mostly primitive, with canines that shear against the lower premolars and a gap (diastema) between the upper incisors and canines. There are, however, some dental features in common with later hominins. The rest of the body also combines ape and human traits, but the lower limbs are clearly meant for walking. The most conspicuous bipedal traits include greatly shortened and broadened pelvic blades with a forward-tilted sacrum, convergent knees, horizontally oriented ankles, and a convergent big toe. Primitive features include curved toes and hands, long toes (although much shorter than those of apes), a conical rib cage, and relatively short thighs. Sexual dimorphism was strong in A. afarensis, males weighing 45 kg (99 pounds) compared with 29 kg (64 pounds) for females. Males stood about 151 cm (4 feet 11 inches), whereas females were about 105 cm (3 feet 5 inches) tall.
The morphology of A. afarensis is a mosaic of primitive features and evolutionary developments shared by later hominins. Its skull is primitive in having a crest and a strongly projecting (prognathic) lower face. The brain was about one-third the size of a modern human's. The dentition is also mostly primitive, with canines that shear against the lower premolars and a gap (diastema) between the upper incisors and canines. There are, however, some dental features in common with later hominins. The rest of the body also combines ape and human traits, but the lower limbs are clearly meant for walking. The most conspicuous bipedal traits include greatly shortened and broadened pelvic blades with a forward-tilted sacrum, convergent knees, horizontally oriented ankles, and a convergent big toe. Primitive features include curved toes and hands, long toes (although much shorter than those of apes), a conical rib cage, and relatively short thighs. Sexual dimorphism was strong in A. afarensis, males weighing 45 kg (99 pounds) compared with 29 kg (64 pounds) for females. Males stood about 151 cm (4 feet 11 inches), whereas females were about 105 cm (3 feet 5 inches) tall.In 1995 a lower jaw resembling that of A. afarensis came to light from Koro Toro, a site in the Baḥr el-Ghazāl region of northern Chad. It is 3.5–3.0 million years old and was assigned to a new species, A. bahrelghazali. In many respects it resembles East African A. afarensis, but it differs in significant details of the jaw articulation and teeth. A. bahrelghazali is the first Pliocene Epoch hominin known from central Africa and stretches the geographic range of Australopithecus 2,500 km (1,500 miles) westward.
A. garhi (2.5 mya), discovered near Hadar at Bouri in the Afar region of Ethiopia, resembles the more primitive A. afarensis more than it does A. africanus (described below). A. garhi has a projecting lower face, enormous cheek teeth, a shallow palate, a large gap (diastema) between the incisor and canine teeth, and forward-pitched incisors. Relative to the length of the upper arm, its thigh is elongated in a way approaching Homo, but its forearm is relatively long, as in apes. A. garhi is found in association with animal bones bearing cut marks that may indicate one of the earliest occurrences of tool use.
Australopithecus africanus

 In 1925 anthropologist Raymond Dart (Dart, Raymond A.) coined the genus name Australopithecus to identify a child's skull recovered from mining operations at Taung in South Africa. He called it Australopithecus africanus, meaning “southern ape of Africa.” From then until 1960 almost all that was known about australopiths came from limestone caves in South Africa. The richest source is at Sterkfontein, where Robert Broom and his team collected hundreds of specimens beginning in 1936. At first Broom simply bought fossils, but in 1946 he began excavating, aided by a crew of skillful workers. Excavation continues to this day. Sterkfontein is one of the richest sources of information about human evolution in the time period between about 3.0 and 2.5 mya. The A. africanus remains of Sterkfontein include skulls, jaws, and numerous skeletal fragments. In 1947 a partial skeleton was unearthed that revealed the humanlike specializations for bipedalism now known to be characteristic of all australopiths. Almost all of the A. africanus remains from Sterkfontein come from a deposit where there is a conspicuous absence of stone tools. An older deposit contains a beautifully preserved skeleton and skull of what might be an early variant of A. africanus. Another source of A. africanus is at Makapansgat, South Africa, where Dart and his team collected about 40 specimens during expeditions from 1947 to 1962.
In 1925 anthropologist Raymond Dart (Dart, Raymond A.) coined the genus name Australopithecus to identify a child's skull recovered from mining operations at Taung in South Africa. He called it Australopithecus africanus, meaning “southern ape of Africa.” From then until 1960 almost all that was known about australopiths came from limestone caves in South Africa. The richest source is at Sterkfontein, where Robert Broom and his team collected hundreds of specimens beginning in 1936. At first Broom simply bought fossils, but in 1946 he began excavating, aided by a crew of skillful workers. Excavation continues to this day. Sterkfontein is one of the richest sources of information about human evolution in the time period between about 3.0 and 2.5 mya. The A. africanus remains of Sterkfontein include skulls, jaws, and numerous skeletal fragments. In 1947 a partial skeleton was unearthed that revealed the humanlike specializations for bipedalism now known to be characteristic of all australopiths. Almost all of the A. africanus remains from Sterkfontein come from a deposit where there is a conspicuous absence of stone tools. An older deposit contains a beautifully preserved skeleton and skull of what might be an early variant of A. africanus. Another source of A. africanus is at Makapansgat, South Africa, where Dart and his team collected about 40 specimens during expeditions from 1947 to 1962.A. africanus is assigned only an approximate geologic age because the only dating method applicable is biostratigraphy. This indirect method compares accompanying animal fossils with those found in other African sites that have been dated more precisely using radiometric methods. The oldest dates are approximately 3.3 mya for hominin specimens (perhaps A. africanus) discovered in the late 1990s at Sterkfontein. Most of the samples of this species are between about 3.0 and perhaps 2.4 million years old.
A. africanus resembles A. afarensis in many respects but also shares unique features with early Homo that are not present in the more primitive A. afarensis. These include reduced facial projection (although there is considerable variation within A. africanus). It also possesses unique specializations not seen in A. afarensis or in early Homo that are related to powerful chewing, such as expansion of the cheek teeth, increased jaw size, and changes to the skull to accommodate the forces generated. Compared with those of A. afarensis, the lower limbs of A. africanus appear to be smaller and the upper limbs larger. Males weighed approximately 41 kg (90 pounds) and stood 138 cm (4 feet 6 inches) tall. Females weighed about 30 kg (66 pounds) and stood 115 cm (3 feet 9 inches) tall. Brain size averages 448 cc (27 cubic inches), closer to modern chimpanzees (395 cc) than to humans (1,350 cc).
Paranthropus aethiopicus
Paranthropus aethiopicus (2.7–2.3 mya) is the earliest of the so-called “robust” australopiths, a group that also includes P. robustus and P. boisei (described below). Robust refers to exaggerated features of the skull, but it does not imply robusticity in any other aspects of the body. The expansion of cheek teeth and supporting structures for grinding hard, tough food continues in later australopiths.
Further specializations for strong chewing occur in P. aethiopicus fossils from the Omo remains (Omo), discovered in the Omo River valley in southern Ethiopia, and in remains found on the western shore of Lake Turkana (Turkana) in northern Kenya. Most of the remains are in the form of isolated teeth and fragmentary jaws, but one remarkably complete skull from 2.5 mya (the “Black Skull”) was recovered from West Turkana. In features related to chewing, P. aethiopicus resembles the East African P. boisei (2.2–1.3 mya) in having enormous molars and premolars, a thick palate and jaws, and projecting cheekbones. In other respects, however, P. aethiopicus shares the primitive morphology of A. afarensis in having a projecting lower face, a large rear portion for attachment of the jaw muscle (temporalis), and flat cranial bones, among other features. Its resemblance to P. boisei may be attributable to their similar diets rather than to a closely shared descent.
Paranthropus robustus and P. boisei
 Paranthropus robustus and P. boisei are also referred to as “robust” australopiths. Some paleoanthropologists classify these two species as Australopithecus, but they appear to be closely related and distinctly different from other australopiths. In addition to a well-developed skull crest for the attachment of chewing muscles, other specializations for strong chewing include huge cheek teeth, massive jaws, and powerfully built cheekbones that project forward. These features make their skulls look very unlike those of modern humans.
Paranthropus robustus and P. boisei are also referred to as “robust” australopiths. Some paleoanthropologists classify these two species as Australopithecus, but they appear to be closely related and distinctly different from other australopiths. In addition to a well-developed skull crest for the attachment of chewing muscles, other specializations for strong chewing include huge cheek teeth, massive jaws, and powerfully built cheekbones that project forward. These features make their skulls look very unlike those of modern humans.Robert Broom recovered the first specimen of a robust australopith in 1938 from the South African cave site of Kromdraai. He gave it the name Paranthropus robustus and noted its hominin features as well as its exaggerated chewing apparatus. Between 1948 and 1952 similar fossils were unearthed from Swartkrans, South Africa, which proved to be another of the richest sources of early hominins. A third source of P. robustus is the limestone cave of Drimolen, South Africa, where a team began collecting in 1992. All three sites are located within a few kilometres of one another in a valley about 30 km (18 miles) west of Johannesburg. As with the remains of A. africanus, the only method of dating the P. robustus remains is via biostratigraphy, which indicates that P. robustus dates from about 1.8–1.5 mya. Specimens attributed to Homo also occur in the same deposits, but these are much rarer.
Broom's choice of the name Paranthropus (meaning “to the side of humans”) reflects his view that this genus was not directly ancestral to later hominins, and it has long been viewed as a distant side branch on the human evolutionary tree. Its specializations for strong chewing certainly make it appear bizarre. The choice of the name robustus referred to its heavily built jaws, teeth, and supporting structures. Its body was relatively petite, however, males weighing about 40 kg (88 pounds) and females about 32 kg (70 pounds). Its brain size is 523 cc, which is both absolutely and relatively larger than that of the earlier South African australopith, A. africanus, with its average brain of 448 cc.
 The spectacular 1959 discovery of a nearly complete skull by Mary Leakey (Leakey, Mary Douglas) at Olduvai Gorge, Tanzania, first revealed the presence of a robust australopith in East Africa. It shares with its South African cousin the combination of chewing specializations and Homo-like evolutionary novelties not present in earlier australopiths. For this reason it is included in the same genus as the South African Paranthropus, but it is different enough to warrant its own species name, P. boisei. It dates to 2.2–1.3 mya, and in that interval it is the most abundant hominin species known, with specimens numbering in the hundreds. It has the greatest development of features related to chewing (mastication), possessing truly massive cheek teeth and jaws. It lived at the same time as species of early Homo, but there is some evidence that Homo and P. boisei preferred different habitats. Despite the enormity of its chewing apparatus, it had a relatively small body, with males weighing about 49 kg (108 pounds) and females 34 kg (75 pounds). P. robustus and P. boisei fossils are found with mammals that are usually associated with dry grassland habitats.
The spectacular 1959 discovery of a nearly complete skull by Mary Leakey (Leakey, Mary Douglas) at Olduvai Gorge, Tanzania, first revealed the presence of a robust australopith in East Africa. It shares with its South African cousin the combination of chewing specializations and Homo-like evolutionary novelties not present in earlier australopiths. For this reason it is included in the same genus as the South African Paranthropus, but it is different enough to warrant its own species name, P. boisei. It dates to 2.2–1.3 mya, and in that interval it is the most abundant hominin species known, with specimens numbering in the hundreds. It has the greatest development of features related to chewing (mastication), possessing truly massive cheek teeth and jaws. It lived at the same time as species of early Homo, but there is some evidence that Homo and P. boisei preferred different habitats. Despite the enormity of its chewing apparatus, it had a relatively small body, with males weighing about 49 kg (108 pounds) and females 34 kg (75 pounds). P. robustus and P. boisei fossils are found with mammals that are usually associated with dry grassland habitats.Relationship to Homo
Quality of the fossil record
Despite the fact that hominins were a rare and insignificant part of the mammalian fauna before about 40,000 years ago, Africans (anthropologists and nonanthropologists alike) and their international colleagues have had phenomenal success in exposing a rich fossil record of australopiths. However, abundant as the fossils are, there are still limitations. For example, the evidence is restricted geographically. The first two-thirds of the fossil record comes almost entirely from sites in the East African Rift Valley and from limestone caves in South Africa. The exceptions are Sahelanthropus tchadensis and the jaw fragment from Baḥr el-Ghazāl in Chad, which call attention to the strong likelihood that other hominins lived throughout tropical and subtropical Africa but left fossils that have not yet been found.Even with comparatively rich samples of species such as Australopithecus afarensis and A. africanus, most of the specimens are very fragmentary, and even partial skeletons are rare. The A. afarensis skeleton “Lucy” stands almost alone in its completeness for the first several million years, joined only by a skeleton from Sterkfontein. The rarity of skeletons makes the reconstruction of body size and shape dependent on many assumptions, which can be subject to interpretation. Another limitation to understanding arises from homoplasy, the appearance of similarities in separate evolutionary lineages. Homoplasy was common in hominin evolution. Various evolutionary novelties appear in the record over time, but many must have evolved independently—for example, extreme expansion of the cheek teeth and all the chewing structures of “robust” australopith species and, to a lesser extent, of A. afarensis, A. africanus, and A. garhi. Extreme development of such traits links the robust australopiths—P. aethiopicus, P. boisei, and P. robustus—into a separate lineage.
A related difficulty is the limited understanding of character transformations. Are all traits truly independent evolutionary novelties, or are some of them part of complexes that change together? Jaw size and tooth size, for example, are not independent, and flexion of the base of the skull and being flat-faced are generally correlated. Taxonomic grouping (taxonomy) based on shared evolutionary novelties (cladistic analysis) brings these correlations into focus. Research on developmental biology will provide important clues about the evolutionary independence of characteristics.
The limitations outlined above are important, but they must be balanced by appreciation of successes. These successes can be organized in many ways. For example, the accumulation of evolutionary novelties can be followed over time. This approach appears obvious, but it has its subtlety, acknowledging that any known sample of fossils represents a species that was successful at the time but was not necessarily the direct ancestor of later species. Speciation probably occurred in small, isolated, peripheral populations that the fossil record has not sampled. What we collect is what was successful at the time. Therefore, we might expect to find many unique characteristics (autapomorphies) of fossil species that exclude them from direct ancestry but that also provide keys to reconstructing the common ancestor of later species.
From this point of view, the fossil record is superb. One can follow the hominin lineage step by step as the accumulation of humanlike characteristics. Oddities may be autapomorphies of a particular species, but they do not necessarily exclude the possibility that it and subsequent species shared a common ancestor. It is important to realize that this accumulation has no predetermined direction. We look back on history and see patterns, but these patterns were not established in advance, as evolution has no predetermined direction.
Changes in anatomy
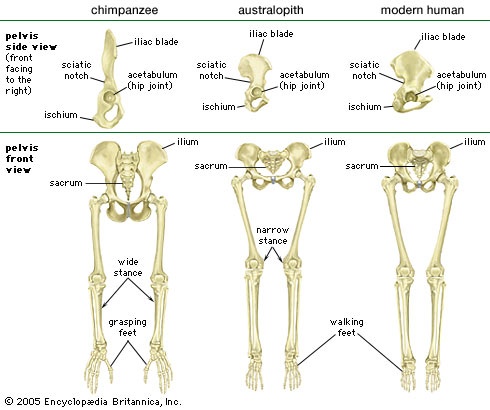 The hominin fossil record does not include a truly intermediate form between an apelike and a humanlike body. Australopithecus retains many primitive apelike traits, but, unlike any ape, it is fundamentally reorganized and highly specialized for walking upright. This state of development required a profound alteration in the genetic template to produce short pelvic blades, a forward-pointing big toe (adducted hallux), and other bipedal traits. The precise sequence of these transformations may in fact never be known from the fossil record, because different parts of the bipedal body may have changed at different rates. For example, the pelvic blades may have shortened before the big toe straightened. Such is the case with the extinct and distinctly nonhuman ape, Oreopithecus, which appears to have had reduced pelvic blades but retained a divergent big toe.
The hominin fossil record does not include a truly intermediate form between an apelike and a humanlike body. Australopithecus retains many primitive apelike traits, but, unlike any ape, it is fundamentally reorganized and highly specialized for walking upright. This state of development required a profound alteration in the genetic template to produce short pelvic blades, a forward-pointing big toe (adducted hallux), and other bipedal traits. The precise sequence of these transformations may in fact never be known from the fossil record, because different parts of the bipedal body may have changed at different rates. For example, the pelvic blades may have shortened before the big toe straightened. Such is the case with the extinct and distinctly nonhuman ape, Oreopithecus, which appears to have had reduced pelvic blades but retained a divergent big toe.Regardless of when or how traits arose, bipedalism is the diagnostic criterion for the evolutionary departure of the human tribe from apes. Bipedal behaviour, however, would have arisen before any fossil evidence of adaptations to it. Thus, the very first hominins most likely had rather apelike bodies without the adaptations for bipedalism that later became the hallmark of the human lineage. These African and possibly European species dating to the late Miocene Epoch (11.2–5.3 million years ago) came, in Charles Darwin (Darwin, Charles)'s words, “to live somewhat less on trees and more on the ground,” owing to “a change in its manner of procuring subsistence, or to a change in the conditions of its native country.”
Although Darwin and his contemporaries predicted much of what the human fossil record would eventually reveal, no one anticipated the discovery of hominins with massive jaws. African apes and modern humans have small cheek teeth relative to body size. Australopiths, on the other hand, had huge molars and premolars with concomitantly gigantic jaws, buttressed cheekbones and face bones, and large areas on the skull for the attachment of chewing muscles.
There appear to be two major structural shifts in the evolution of the human body. The first was the transition to bipedalism that is documented in A. anamensis, A. afarensis, A. africanus, and A. garhi, which span a time frame from 4.2 to 2.5 mya. The limbs and torsos among these species are difficult to assess because of the incompleteness of the fossil record. All share features with Homo, but only A. afarensis and A. africanus are complete enough to make detailed comparisons. These two species share a similar mixture of apelike, humanlike, and unique features in their wrists, hips, and knees. They apparently differ in limb joint sizes, however, with A. africanus appearing to be more apelike even though it lived later in time and had a more Homo-like skull and teeth. Both species appear to share a combination of specialized bipedal traits but are not exactly like modern humans in that they possess upper limb features associated with climbing.
The second major change in evolution appears at about 1.9 mya with the appearance of hips (hip) that are uniquely Homo. Long femurs and relatively enlarged hip joints mark a significant change in locomotion that is related, perhaps, to long-distance, efficient striding more like that seen in modern H. sapiens (Homo sapiens). The discovery of A. garhi reveals the complexity of tracing evolution of limb proportions in that it had a humanlike femur-to-upper-arm ratio yet a long, apelike forearm.
There are opposing interpretations of the primitive body traits retained in the early species of Australopithecus. One view emphasizes the bipedal specializations, whereas the other calls attention to the many primitive skull characteristics. Even so, both camps agree that all species of Australopithecus were bipedal and thus did not climb like apes. Australopiths did, however, retain features associated with tree-dwelling for at least a million years. Their different hip architecture implies some difference from modern humans in gait and climbing ability. The divergence between Australopithecus and later-appearing Homo became clearer with the discoveries of lower-body fossils associated with Homo erectus, particularly the “Strapping Youth,” also called “Turkana Boy,” found at Nariokotome, Kenya, in 1984. The striking difference between the pelvis (pelvic girdle) and femur of Australopithecus and those of Homo probably registers a major shift in adaptation between the two groups. From this perspective, Australopithecus appears to have had the hands free for carrying but was adapted only to traveling short distances. It likely had a healthy appreciation of trees for safety, feeding, and sleeping. The longer femur and more humanlike pelvis that appear by 1.9 mya in Homo mark the beginning of an important change.
Not only were there numerous species of human predecessors long ago, but many of these overlapped in time and space. Habitats favourable for hominin occupation undoubtedly appeared and disappeared throughout much of Africa over and over again with the drastic fluctuations in tropical climates that occurred during the Pliocene (Pliocene Epoch) and Pleistocene (Pleistocene Epoch) epochs. More species presumably await discovery, because there were probably many evolutionary experiments in these varied and changing habitats. Although the current sample of fossil hominins leads some to the impression that there were only a few hominin lineages, it is far more likely that the human family tree will turn out to be quite “bushy.” Species names may need to multiply to accommodate the diversity, although a balance needs to be maintained between excessive splitting groups apart and lumping them together.
Evidence regarding the relationship of Australopithecus to the origin of the genus Homo may appear to conflict, but, from the perspective of accumulated shared traits, the fossil record is less perplexing. Put simply, brains expand and cheek teeth shrink. In H. habilis (Homo habilis) (2.3–1.6 mya) the body appears to remain like that of Australopithecus—small with relatively large upper limbs and small lower limbs. If the lower limb fossils found with the skulls and teeth of a 1.9-million-year-old specimen of H. rudolfensis also belong to this species, then the more humanlike body proportions and hip architecture first appear in this species just after 2 mya. Both H. habilis and H. rudolfensis are transitional, with some primitive and some derived characteristics of later Homo species. Other skeletal remains are critical here because body size appears to be very different. H. habilis was very small (35 kg 【77 pounds】), and H. rudolfensis was large (55 kg 【121 pounds】). Scaling cheek-tooth size to body weight shows that they both had reversed the trend of ever-increasing cheek-tooth size. Relative brain size expanded, especially in H. habilis. Brain size expanded further with the appearance of H. erectus (Homo erectus) by at least 1.8 mya, but body size also increased, so that relative brain size apparently was not so dramatically expanded. The early African form of H. erectus is often referred to as H. ergaster to contrast it with the well-known Asian H. erectus. Body size and especially hind limb length reach modern proportions in this species. Other traits Australopithecus has in common with later Homo include a further reduction in facial projection as well as other features, including reduction in the size of the cheek teeth. Brains then continue to expand and cheek teeth become progressively smaller through the evolution of the genus Homo.
Additional Reading
Donald C. Johanson and Maitland A. Edey, Lucy: The Beginnings of Humankind (1981, reissued 1990), recounts the field expeditions of Lucy's discoverer (Johanson) and provides background on other human ancestors in addition to Australopithecus afarensis. Meave G. Leakey and Friedemann Schrenk, History of the Anthropoid: The Search for the Beginning (1997), produced by Films for the Humanities, is a video documentary in which Meave Leakey discusses human evolutionary theory at Tanzanian paleontological sites. William H. Kimbel, Yoel Rak, and Donald C. Johanson, The Skull of Australopithecus afarensis (2004), is a complete scientific text.
- William A Brady
- William Adams
- William Addison Dwiggins
- William-Adolphe Bouguereau
- William A. Fowler
- William Alabaster
- William Allen
- William Allen White
- William Ames
- William; and Hare Burke, William
- William; and Hare, William Burke
- William and Mary, College of
- William and Mary style
- William Archer
- William Arnold Anthony
- William Augustus Brevoort Coolidge
- William Augustus Cumberland, Duke of
- William Augustus, Duke of Cumberland
- William Avery Bishop
- William A Wheeler
- William Baffin
- William Bainbridge
- William Balfour Baikie
- God, Truce of
- Godunov, Boris