bacteria
Introduction
singular bacterium
 any of a group of microscopic single-celled organisms that live in enormous numbers in almost every environment on the surface of Earth, from deep-sea vents to the digestive tracts of humans.
any of a group of microscopic single-celled organisms that live in enormous numbers in almost every environment on the surface of Earth, from deep-sea vents to the digestive tracts of humans.Bacteria lack a membrane-bound nucleus and other internal structures and are therefore ranked among the unicellular life-forms called prokaryotes (prokaryote). Prokaryotes are the dominant living creatures on Earth, having been present for perhaps three-quarters of Earth history and having adapted to almost all available ecological habitats. As a group, they display exceedingly diverse metabolic capabilities and can use almost any organic compound, and some inorganic compounds (inorganic compound), as a food source. Some bacteria can cause diseases in humans, animals, or plants, but most are harmless and are beneficial ecological agents whose metabolic activities sustain higher life-forms. Other bacteria are symbionts (symbiosis) of plants and invertebrates, where they carry out important functions for the host, such as nitrogen fixation and cellulose degradation. Without prokaryotes, soil would not be fertile, and dead organic material would decay much more slowly. Some bacteria are widely used in the preparation of foods, chemicals, and antibiotics (antibiotic). Studies of the relationships between different groups of bacteria continue to yield new insights into the origin of life on Earth and the mechanisms of evolution.
The bacterial (prokaryote) cell
Bacteria as prokaryotes

 All living organisms on Earth are made up of one of two basic types of cells: eukaryotic (eukaryote) cells, in which the genetic material is enclosed within a nuclear membrane, or prokaryotic (prokaryote) cells, in which the genetic material is not separated from the rest of the cell. Traditionally, all prokaryotic cells were called bacteria and were classified in the prokaryotic kingdom Monera (moneran). However, their classification as Monera, equivalent in taxonomy to the other kingdoms—Plantae, Animalia, Fungi, and Protista—understated the remarkable genetic and metabolic diversity exhibited by prokaryotic cells relative to eukaryotic cells. In the late 1970s American microbiologist Carl Woese pioneered a major change in classification by placing all organisms into three domains—Eukarya, Bacteria (originally called Eubacteria), and archaea (originally called Archaebacteria)—to reflect the three ancient lines of evolution. The prokaryotic organisms that were formerly known as bacteria were then divided into two of these domains, Bacteria and Archaea. Bacteria and Archaea are superficially similar; for example, they do not have intracellular organelles, and they have circular DNA. However, they are fundamentally distinct, and their separation is based on the genetic evidence for their ancient and separate evolutionary lineages, as well as fundamental differences in their chemistry and physiology. Members of these two prokaryotic domains are as different from one another as they are from eukaryotic cells.
All living organisms on Earth are made up of one of two basic types of cells: eukaryotic (eukaryote) cells, in which the genetic material is enclosed within a nuclear membrane, or prokaryotic (prokaryote) cells, in which the genetic material is not separated from the rest of the cell. Traditionally, all prokaryotic cells were called bacteria and were classified in the prokaryotic kingdom Monera (moneran). However, their classification as Monera, equivalent in taxonomy to the other kingdoms—Plantae, Animalia, Fungi, and Protista—understated the remarkable genetic and metabolic diversity exhibited by prokaryotic cells relative to eukaryotic cells. In the late 1970s American microbiologist Carl Woese pioneered a major change in classification by placing all organisms into three domains—Eukarya, Bacteria (originally called Eubacteria), and archaea (originally called Archaebacteria)—to reflect the three ancient lines of evolution. The prokaryotic organisms that were formerly known as bacteria were then divided into two of these domains, Bacteria and Archaea. Bacteria and Archaea are superficially similar; for example, they do not have intracellular organelles, and they have circular DNA. However, they are fundamentally distinct, and their separation is based on the genetic evidence for their ancient and separate evolutionary lineages, as well as fundamental differences in their chemistry and physiology. Members of these two prokaryotic domains are as different from one another as they are from eukaryotic cells.Prokaryotic cells (i.e., Bacteria and Archaea) are fundamentally different from the eukaryotic (eukaryote) cells that constitute other forms of life. Prokaryotic cells are defined by a much simpler design than is found in eukaryotic cells. The most apparent simplification is the lack of intracellular organelles (organelle), which are features characteristic of eukaryotic cells. Organelles are discrete membrane-enclosed structures that are contained in the cytoplasm and include the nucleus, where genetic information is retained, copied, and expressed; the mitochondria and chloroplasts (chloroplast), where chemical or light energy is converted into metabolic energy; the lysosome, where ingested proteins are digested and other nutrients are made available; and the endoplasmic reticulum and the Golgi complex, where the proteins (protein) that are synthesized by and released from the cell are assembled, modified, and exported. All of the activities performed by organelles also take place in bacteria, but they are not carried out by specialized structures. In addition, prokaryotic cells are usually much smaller than eukaryotic cells. The small size, simple design, and broad metabolic capabilities of bacteria allow them to grow and divide very rapidly and to inhabit and flourish in almost any environment.
Prokaryotic and eukaryotic cells differ in many other ways, including lipid composition, structure of key metabolic enzymes (enzyme), responses to antibiotics and toxins (toxin), and the mechanism of expression of genetic information. Eukaryotic organisms contain multiple linear chromosomes with genes that are much larger than they need to be to encode the synthesis of proteins. Substantial portions of the ribonucleic acid ( RNA) copy of the genetic information (deoxyribonucleic acid, or DNA) are discarded, and the remaining messenger RNA (mRNA) is substantially modified before it is translated into protein. In contrast, bacteria have one circular chromosome that contains all of their genetic information, and their mRNAs are exact copies of their gene and are not modified.
Diversity of structure of bacteria
Although bacterial cells are much smaller and simpler in structure than eukaryotic cells, the bacteria are an exceedingly diverse group of organisms that differ in size, shape, habitat, and metabolism. Much of the knowledge about bacteria has come from studies of disease-causing bacteria, which are more readily isolated in pure culture and more easily investigated than are many of the free-living species of bacteria. It must be noted that many free-living bacteria are quite different from the bacteria that are adapted to live as animal parasites (parasitism). Thus, there are no absolute rules about bacterial composition or structure, and there are many exceptions to any general statement.
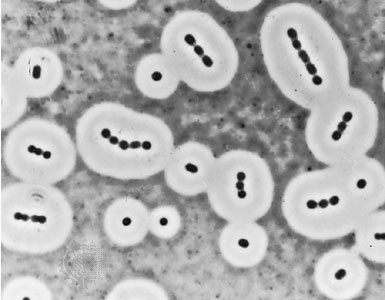
 Individual bacteria can assume one of three basic shapes: spherical ( coccus), rodlike ( bacillus), or curved ( vibrio, Spirillum, or spirochete). Considerable variation is seen in the actual shapes of bacteria, and cells can be stretched or compressed in one dimension. Bacteria that do not separate from one another after cell division form characteristic clusters that are helpful in their identification. For example, some cocci are found mainly in pairs, including Streptococcus pneumoniae, a pneumococcus that causes bacterial lobar pneumonia, and Neisseria gonorrhoeae, a gonococcus that causes the sexually transmitted disease gonorrhea. Most streptococci (Streptococcus) resemble a long strand of beads, whereas the staphylococci (staphylococcus) form random clumps (the name “staphylococci” is derived from the Greek word staphyle, meaning “cluster of grapes”). In addition, some coccal bacteria occur as square or cubical packets. The rod-shaped bacilli usually occur singly, but some strains form long chains, such as rods of the corynebacteria, normal inhabitants of the mouth that are frequently attached to one another at random angles. Some bacilli have pointed ends, whereas others have squared ends, and some rods are bent into a comma shape. These bent rods are often called vibrios and include Vibrio cholerae, which causes cholera. Other shapes of bacteria include the spirilla, which are bent and rebent, and the spirochetes, which form a helix similar to a corkscrew, in which the cell body is wrapped around a central fibre called the axial filament.
Individual bacteria can assume one of three basic shapes: spherical ( coccus), rodlike ( bacillus), or curved ( vibrio, Spirillum, or spirochete). Considerable variation is seen in the actual shapes of bacteria, and cells can be stretched or compressed in one dimension. Bacteria that do not separate from one another after cell division form characteristic clusters that are helpful in their identification. For example, some cocci are found mainly in pairs, including Streptococcus pneumoniae, a pneumococcus that causes bacterial lobar pneumonia, and Neisseria gonorrhoeae, a gonococcus that causes the sexually transmitted disease gonorrhea. Most streptococci (Streptococcus) resemble a long strand of beads, whereas the staphylococci (staphylococcus) form random clumps (the name “staphylococci” is derived from the Greek word staphyle, meaning “cluster of grapes”). In addition, some coccal bacteria occur as square or cubical packets. The rod-shaped bacilli usually occur singly, but some strains form long chains, such as rods of the corynebacteria, normal inhabitants of the mouth that are frequently attached to one another at random angles. Some bacilli have pointed ends, whereas others have squared ends, and some rods are bent into a comma shape. These bent rods are often called vibrios and include Vibrio cholerae, which causes cholera. Other shapes of bacteria include the spirilla, which are bent and rebent, and the spirochetes, which form a helix similar to a corkscrew, in which the cell body is wrapped around a central fibre called the axial filament.Bacteria are the smallest living creatures. An average-size bacterium, such as the rod-shaped Escherichia coli, a normal inhabitant of the intestinal tract (intestine) of humans and animals, is about 2 micrometres (μm; millionths of a metre) long and 0.5 μm in diameter, and the spherical cells of Staphylococcus aureus (staphylococcus) are up to 1 μm in diameter. A few bacterial types are even smaller, such as Mycoplasma pneumoniae (mycoplasma), which is one of the smallest bacteria, ranging from about 0.1 to 0.25 μm in diameter; the rod-shaped Bordetella pertussis, which is the causative agent of whooping cough, ranging from 0.2 to 0.5 μm in diameter and 0.5 to 1 μm in length; and the corkscrew-shaped Treponema pallidum, which is the causative agent of syphilis, averaging only 0.15 μm in diameter but 10 to 13 μm in length. Some bacteria are relatively large, such as Azotobacter, which has diameters of 2 to 5 μm or more; the cyanobacterium Synechococcus, which averages 6 μm by 12 μm; and Achromatium, which has a minimum width of 5 μm and a maximum length of 100 μm, depending on the species. Giant bacteria can be visible with the unaided eye, such as Titanospirillum namibiensis, which averages 750 μm in diameter, and the rod-shaped Epulopsicium fishelsoni, which averages 80 μm in diameter by 600 μm in length.
Bacteria are unicellular microorganisms and thus are generally not organized into tissues. Each bacterium grows and divides independently of any other bacterium, although aggregates of bacteria, sometimes containing members of different species, are frequently found. Many bacteria can form aggregated structures called biofilms (biofilm). Organisms in biofilms often display substantially different properties from the same organism in the individual state or the planktonic state. Bacteria that have aggregated into biofilms can communicate information about population size and metabolic state. This type of communication is called quorum sensing and operates by the production of small molecules called autoinducers or pheromones (pheromone). The concentration of quorum-sensing molecules—most commonly peptides (peptide) or acylated homoserine lactones (AHLs; special signaling chemicals)—is related to the number of bacteria of the same or different species that are in the biofilm and helps coordinate the behaviour of the biofilm.
Morphological features of bacteria
The Gram stain
 Bacteria are so small that their presence was only first recognized in 1677, when the Dutch naturalist Antonie van Leeuwenhoek (Leeuwenhoek, Antonie van) saw microscopic organisms in a variety of substances with the aid of primitive microscopes (microscope) (more similar in design to modern magnifying glasses than modern microscopes), some of which were capable of more than 200-fold magnification. Now bacteria are usually examined under light microscopes capable of more than 1,000-fold magnification; however, details of their internal structure can be observed only with the aid of much more powerful transmission electron microscopes (microscope). Unless special phase-contrast microscopes are used, bacteria have to be stained with a coloured dye so that they will stand out from their background.
Bacteria are so small that their presence was only first recognized in 1677, when the Dutch naturalist Antonie van Leeuwenhoek (Leeuwenhoek, Antonie van) saw microscopic organisms in a variety of substances with the aid of primitive microscopes (microscope) (more similar in design to modern magnifying glasses than modern microscopes), some of which were capable of more than 200-fold magnification. Now bacteria are usually examined under light microscopes capable of more than 1,000-fold magnification; however, details of their internal structure can be observed only with the aid of much more powerful transmission electron microscopes (microscope). Unless special phase-contrast microscopes are used, bacteria have to be stained with a coloured dye so that they will stand out from their background.One of the most useful staining reactions for bacteria is called the Gram stain, developed in 1884 by the Danish physician Hans Christian Gram. Bacteria in suspension are fixed to a glass slide by brief heating and then exposed to two dyes that combine to form a large blue dye complex within each cell. When the slide is flushed with an alcohol solution, gram-positive bacteria retain the blue colour and gram-negative bacteria lose the blue colour. The slide is then stained with a weaker pink dye that causes the gram-negative bacteria to become pink, whereas the gram-positive bacteria remain blue. The Gram stain reacts to differences in the structure of the bacterial cell surface, differences that are apparent when the cells are viewed under an electron microscope.
The cell envelope
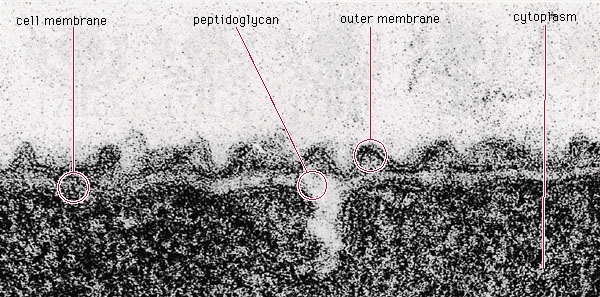 The bacterial cell surface (or envelope) can vary considerably in its structure, and it plays a central role in the properties and capabilities of the cell. The one feature present in all cells is the cytoplasmic membrane, which separates the inside of the cell from its external environment, regulates the flow of nutrients, maintains the proper intracellular milieu, and prevents the loss of the cell's contents. The cytoplasmic membrane carries out many necessary cellular functions, including energy generation, protein secretion, chromosome segregation, and efficient active transport of nutrients. It is a typical unit membrane composed of proteins and lipids, basically similar to the membrane that surrounds all eukaryotic cells. It appears in electron micrographs as a triple-layered structure of lipids and proteins that completely surround the cytoplasm.
The bacterial cell surface (or envelope) can vary considerably in its structure, and it plays a central role in the properties and capabilities of the cell. The one feature present in all cells is the cytoplasmic membrane, which separates the inside of the cell from its external environment, regulates the flow of nutrients, maintains the proper intracellular milieu, and prevents the loss of the cell's contents. The cytoplasmic membrane carries out many necessary cellular functions, including energy generation, protein secretion, chromosome segregation, and efficient active transport of nutrients. It is a typical unit membrane composed of proteins and lipids, basically similar to the membrane that surrounds all eukaryotic cells. It appears in electron micrographs as a triple-layered structure of lipids and proteins that completely surround the cytoplasm.Lying outside of this membrane is a rigid wall that determines the shape of the bacterial cell. The wall is made of a huge molecule called peptidoglycan (or murein). In gram-positive bacteria the peptidoglycan forms a thick meshlike layer that retains the blue dye of the Gram stain by trapping it in the cell. In contrast, in gram-negative bacteria the peptidoglycan layer is very thin (only one or two molecules deep), and the blue dye is easily washed out of the cell.
Peptidoglycan occurs only in the Bacteria (except for those without a cell wall, such as Mycoplasma). Peptidoglycan is a long-chain polymer of two repeating sugars (N-acetylglucosamine and N-acetyl muramic acid), in which adjacent sugar chains are linked to one another by peptide bridges that confer rigid stability. The nature of the peptide bridges differs considerably between species of bacteria but in general consists of four amino acids (amino acid): L- alanine linked to D- glutamic acid, linked to either diaminopimelic acid in gram-negative bacteria or L- lysine, L-ornithine, or diaminopimelic acid in gram-positive bacteria, which is finally linked to D-alanine. In gram-negative bacteria the peptide bridges connect the D-alanine on one chain to the diaminopimelic acid on another chain. In gram-positive bacteria there can be an additional peptide chain that extends the reach of the cross-link; for example, there is an additional bridge of five glycines (glycine) in Staphylococcus aureus.
Peptidoglycan synthesis is the target of many useful antimicrobial agents (antimicrobial agent), including the β-lactam antibiotics (e.g., penicillin) that block the cross-linking of the peptide bridges. Some of the proteins that animals synthesize as natural antibacterial defense factors attack the cell walls of bacteria. For example, an enzyme called lysozyme splits the sugar chains that are the backbone of peptidoglycan molecules. The action of any of these agents weakens the cell wall and disrupts the bacterium.
In gram-positive bacteria the cell wall is composed mainly of a thick peptidoglycan meshwork interwoven with other polymers called teichoic acids (from the Greek word teichos, meaning “wall”) and some proteins or lipids. In contrast, gram-negative bacteria have a complex cell wall that is composed of multiple layers in which an outer membrane layer lies on top of a thin peptidoglycan layer. This outer membrane is composed of phospholipids (phospholipid), which are complex lipids that contain molecules of phosphate, and lipopolysaccharides, which are complex lipids that are anchored in the outer membrane of cells by their lipid end and have a long chain of sugars extending away from the cell into the medium. Lipopolysaccharides, often called endotoxins (endotoxin), are toxic to animals and humans; their presence in the bloodstream can cause fever, shock, and even death. For most gram-negative bacteria, the outer membrane forms a barrier to the passage of many chemicals that would be harmful to the bacterium, such as dyes and detergents (detergent) that normally dissolve cellular membranes. Impermeability to oil-soluble compounds is not seen in other biological membranes and results from the presence of lipopolysaccharides in the membrane and from the unusual character of the outer membrane proteins. As evidence of the ability of the outer membrane to confer resistance to harsh environmental conditions, some gram-negative bacteria grow well in oil slicks, jet fuel tanks, acid mine drainage, and even bottles of disinfectants (disinfectant).
The archaea have markedly different surface structures from the Bacteria. They do not have peptidoglycan; instead, their membrane lipids are made up of branched isoprenoids (isoprenoid) linked to glycerol by ether bonds. Some archaea have a wall material that is similar to peptidoglycan, except that the specific sugar linked to the amino acid bridges is not muramic acid but talosaminuronic acid. Many other archaeal species use proteins as the basic constituent of their walls, and some lack a rigid wall.
Capsules and slime layers
 Many bacterial cells secrete some extracellular material in the form of a capsule or a slime layer. A slime layer is loosely associated with the bacterium and can be easily washed off, whereas a capsule is attached tightly to the bacterium and has definite boundaries. Capsules can be seen under a light microscope by placing the cells in a suspension of India ink. The capsules exclude the ink and appear as clear halos surrounding the bacterial cells. Capsules are usually polymers of simple sugars (polysaccharides), although the capsule of Bacillus anthracis is made of polyglutamic acid. Most capsules are hydrophilic (“water-loving”) and may help the bacterium avoid desiccation (dehydration) by preventing water loss. Capsules can protect a bacterial cell from ingestion and destruction by white blood cells (leukocyte) ( phagocytosis). While the exact mechanism for escaping phagocytosis is unclear, it may occur because capsules make bacterial surface components more slippery, helping the bacterium to escape engulfment by phagocytic cells. The presence of a capsule in Streptococcus pneumoniae (pneumococcus) is the most important factor in its ability to cause pneumonia. Mutant strains of S. pneumoniae that have lost the ability to form a capsule are readily taken up by white blood cells and do not cause disease. The association of virulence and capsule formation is also found in many other species of bacteria.
Many bacterial cells secrete some extracellular material in the form of a capsule or a slime layer. A slime layer is loosely associated with the bacterium and can be easily washed off, whereas a capsule is attached tightly to the bacterium and has definite boundaries. Capsules can be seen under a light microscope by placing the cells in a suspension of India ink. The capsules exclude the ink and appear as clear halos surrounding the bacterial cells. Capsules are usually polymers of simple sugars (polysaccharides), although the capsule of Bacillus anthracis is made of polyglutamic acid. Most capsules are hydrophilic (“water-loving”) and may help the bacterium avoid desiccation (dehydration) by preventing water loss. Capsules can protect a bacterial cell from ingestion and destruction by white blood cells (leukocyte) ( phagocytosis). While the exact mechanism for escaping phagocytosis is unclear, it may occur because capsules make bacterial surface components more slippery, helping the bacterium to escape engulfment by phagocytic cells. The presence of a capsule in Streptococcus pneumoniae (pneumococcus) is the most important factor in its ability to cause pneumonia. Mutant strains of S. pneumoniae that have lost the ability to form a capsule are readily taken up by white blood cells and do not cause disease. The association of virulence and capsule formation is also found in many other species of bacteria. A capsular layer of extracellular polysaccharide material can enclose many bacteria into a biofilm and serves many functions. Streptococcus mutans, which causes dental caries (caries), splits the sucrose in food and uses one of the sugars to build its capsule, which sticks tightly to the tooth. The bacteria that are trapped in the capsule use the other sugar to fuel their metabolism and produce a strong acid ( lactic acid) that attacks the tooth enamel. When Pseudomonas aeruginosa colonizes the lungs of persons with cystic fibrosis, it produces a thick capsular polymer of alginic acid that contributes to the difficulty of eradicating the bacterium. Bacteria of the genus Zoogloea secrete fibres of cellulose that enmesh the bacteria into a floc that floats on the surface of liquid and keeps the bacteria exposed to air, a requirement for the metabolism of this genus. A few rod-shaped bacteria, such as Sphaerotilus, secrete long chemically complex tubular sheaths that enclose substantial numbers of the bacteria. The sheaths of these and many other environmental bacteria can become encrusted with iron or manganese oxides.
A capsular layer of extracellular polysaccharide material can enclose many bacteria into a biofilm and serves many functions. Streptococcus mutans, which causes dental caries (caries), splits the sucrose in food and uses one of the sugars to build its capsule, which sticks tightly to the tooth. The bacteria that are trapped in the capsule use the other sugar to fuel their metabolism and produce a strong acid ( lactic acid) that attacks the tooth enamel. When Pseudomonas aeruginosa colonizes the lungs of persons with cystic fibrosis, it produces a thick capsular polymer of alginic acid that contributes to the difficulty of eradicating the bacterium. Bacteria of the genus Zoogloea secrete fibres of cellulose that enmesh the bacteria into a floc that floats on the surface of liquid and keeps the bacteria exposed to air, a requirement for the metabolism of this genus. A few rod-shaped bacteria, such as Sphaerotilus, secrete long chemically complex tubular sheaths that enclose substantial numbers of the bacteria. The sheaths of these and many other environmental bacteria can become encrusted with iron or manganese oxides.Flagella, fimbriae, and pili
Many bacteria are motile, able to swim through a liquid medium or glide or swarm across a solid surface. Swimming and swarming bacteria possess flagella (flagellum), which are the extracellular appendages needed for motility. Flagella are long, helical filaments made of a single type of protein and located either at the ends of rod-shaped cells, as in Vibrio cholerae or Pseudomonas aeruginosa, or all over the cell surface, as in Escherichia coli. Flagella can be found on both gram-positive and gram-negative rods but are rare on cocci and are trapped in the axial filament in the spirochetes. The flagellum is attached at its base to a basal body in the cell membrane. The protomotive force generated at the membrane is used to turn the flagellar filament, in the manner of a turbine driven by the flow of hydrogen ions through the basal body into the cell. When the flagella are rotating in a counterclockwise direction, the bacterial cell swims in a straight line; clockwise rotation results in swimming in the opposite direction or, if there is more than one flagellum per cell, in random tumbling. Chemotaxis allows a bacterium to adjust its swimming behaviour so that it can sense and migrate toward increasing levels of an attractant chemical or away from a repellent one.Not only are bacteria able to swim or glide toward more favourable environments, but they also have appendages that allow them to adhere to surfaces and keep from being washed away by flowing fluids. Some bacteria, such as E. coli and Neisseria gonorrhoeae, produce straight, rigid, spikelike projections called fimbriae (Latin for “threads” or “fibres”) or pili (Latin for “hairs”), which extend from the surface of the bacterium and attach to specific sugars on other cells—for these strains, intestinal or urinary-tract epithelial cells (epithelium), respectively. Fimbriae are present only in gram-negative bacteria. Certain pili (called sex pili) are used to allow one bacterium to recognize and adhere to another in a process of sexual mating called conjugation (see below Bacterial reproduction (bacteria)). Many aquatic bacteria produce an acidic mucopolysaccharide holdfast, which allows them to adhere tightly to rocks or other surfaces.
The cytoplasm (cytoplasm)
Although bacteria differ substantially in their surface structures, their interior contents are quite similar and display relatively few structural features.
Genetic content
The genetic information of all cells resides in the sequence of nitrogenous bases in the extremely long molecules of DNA. Unlike the DNA in eukaryotic cells, which resides in the nucleus, DNA in bacterial cells is not sequestered in a membrane-bound organelle but appears as a long coil distributed through the cytoplasm. In many bacteria the DNA is present as a single, circular chromosome, although some bacteria may contain two chromosomes, and in some cases the DNA is linear rather than circular. A variable number of smaller, usually circular DNA molecules, called plasmids (plasmid), can carry auxiliary information.
The sequence of bases in the DNA has been determined for hundreds of bacteria. The amount of DNA in bacterial chromosomes ranges from 580,000 base pairs in Mycoplasma gallinarum to 4,700,000 base pairs in E. coli to 9,140,000 base pairs in Myxococcus xanthus. The length of the E. coli chromosome is about 1.2 mm, which is striking in view of the fact that the length of the cell is about 0.001 mm.
As in all organisms, bacterial DNA contains the four nitrogenous bases adenine (A), cytosine (C), guanine (G), and thymine (T). The rules of base pairing for double-stranded DNA molecules require that the number of adenine and thymine bases be equal and that the number of cytosine and guanine bases also be equal. The relationship between the number of pairs of G and C bases and the number of pairs of A and T bases is an important indicator of evolutionary and adaptive genetic changes within an organism. The proportion, or molar ratio, of G + C can be measured as G + C divided by the sum of all the bases (A + T + G + C) multiplied by 100 percent. The extent to which G + C ratios vary between organisms may be considerable. In plants and animals, the proportion of G + C is about 50 percent. A far wider range in the proportion of G + C is seen in prokaryotes, extending from about 25 percent in most Mycoplasma to about 50 percent in E. coli to nearly 75 percent in Micrococcus, actinomycetes (actinomycete), and fruiting myxobacteria. The G + C content within a species in a single genus, however, is very similar.
Cytoplasmic structures
The cytoplasm of bacteria contains high concentrations of enzymes, metabolites, and salts. In addition, the proteins of the cell are made on ribosomes (ribosome) that are scattered throughout the cytoplasm. Bacterial ribosomes are different from ribosomes in eukaryotic cells in that they are smaller, have fewer constituents (consist of three types of ribosomal RNA 【rRNA】 and 55 proteins, as opposed to four types of rRNA and 78 proteins in eukaryotes), and are inhibited by different antibiotics than those that act on eukaryotic ribosomes.
There are numerous inclusion bodies, or granules, in the bacterial cytoplasm. These bodies are never enclosed by a membrane and serve as storage vessels. glycogen, which is a polymer of glucose, is stored as a reserve of carbohydrate and energy. Volutin, or metachromatic granules, contains polymerized phosphate and represents a storage form for inorganic phosphate and energy. Many bacteria possess lipid droplets that contain polymeric esters of poly-β-hydroxybutyric acid or related compounds. This is in contrast to eukaryotes, which use lipid droplets to store triglycerides. In bacteria, storage granules are produced under favourable growth conditions and are consumed after the nutrients have been depleted from the medium. Many aquatic bacteria produce gas vacuoles, which are protein-bound structures that contain air and allow the bacteria to adjust their buoyancy. Bacteria can also have internal membranous structures that form as outgrowths of the cytoplasmic membrane.
Biotypes of bacteria
The fact that pathogenic bacteria are constantly battling their host's immune system might account for the bewildering number of different strains, or types, of bacteria that belong to the same species but are distinguishable by serological tests (serological test). Microbiologists often identify bacteria by the presence of specific molecules on their cell surfaces, which are detected with specific antibodies (antibody). Antibodies are serum proteins that bind very tightly to foreign molecules (antigens (antigen)) in an immune reaction aimed at removing or destroying the antigens. Antibodies have remarkable specificity, and the substitution of even one amino acid in a protein might prevent that protein from being recognized by an antibody.
In the case of E. coli and Salmonella enterica, there are thousands of different strains (called serovars, for serological variants), which differ from one another mainly or solely in the antigenic identity of their lipopolysaccharide, flagella (flagellum), or capsule. Different serovars of these enteric bacteria are often found to be associated with the ability to inhabit different host animals or to cause different diseases. Formation of these numerous serovars reflects the ability of bacteria to respond effectively to the intense defensive actions of the immune system.
Bacterial reproduction
Reproductive processes
binary fission
Most prokaryotes reproduce by a process of binary fission (binary fission), in which the cell grows in volume until it divides in half to yield two identical daughter cells. Each daughter cell can continue to grow at the same rate as its parent. For this process to occur, the cell must grow over its entire surface until the time of cell division, when a new hemispherical pole forms at the division septum in the middle of the cell. In gram-positive bacteria the septum grows inward from the plasma membrane along the midpoint of the cell; in gram-negative bacteria the walls are more flexible, and the division septum forms as the side walls pinch inward, dividing the cell in two. In order for the cell to divide in half, the peptidoglycan structure must be different in the hemispherical cap than in the straight portion of the cell wall, and different wall-cross-linking enzymes must be active at the septum than elsewhere.
budding
 A group of environmental bacteria reproduces by budding (budding bacterium). In this process a small bud forms at one end of the mother cell or on filaments called prosthecae. As growth proceeds, the size of the mother cell remains about constant, but the bud enlarges. When the bud is about the same size as the mother cell, it separates. This type of reproduction is analogous to that in budding fungi, such as brewer's yeast (Saccharomyces cerevisiae (Saccharomyces)). One difference between fission and budding is that, in the latter, the mother cell often has different properties from the offspring. In some Pasteuria strains, the daughter buds have a flagellum and are motile, whereas the mother cells lack flagella but have long pili and holdfast appendages at the end opposite the bud. The related Planctomyces, found in plankton, have long fibrillar stalks at the end opposite the bud. In Hyphomicrobium a hyphal filament (prostheca) grows out of one end of the cell, and the bud grows out of the tip of the prostheca, separated by a relatively long distance from the mother cell.
A group of environmental bacteria reproduces by budding (budding bacterium). In this process a small bud forms at one end of the mother cell or on filaments called prosthecae. As growth proceeds, the size of the mother cell remains about constant, but the bud enlarges. When the bud is about the same size as the mother cell, it separates. This type of reproduction is analogous to that in budding fungi, such as brewer's yeast (Saccharomyces cerevisiae (Saccharomyces)). One difference between fission and budding is that, in the latter, the mother cell often has different properties from the offspring. In some Pasteuria strains, the daughter buds have a flagellum and are motile, whereas the mother cells lack flagella but have long pili and holdfast appendages at the end opposite the bud. The related Planctomyces, found in plankton, have long fibrillar stalks at the end opposite the bud. In Hyphomicrobium a hyphal filament (prostheca) grows out of one end of the cell, and the bud grows out of the tip of the prostheca, separated by a relatively long distance from the mother cell.Sporulation
Many environmental bacteria are able to produce stable dormant, or resting, forms as a branch of their life cycle to enhance their survival under adverse conditions. These processes are not an obligate stage of the cell's life cycle but rather an interruption. Such dormant forms are called endospores, cysts, or heterocysts (primarily seen in cyanobacteria), depending on the method of spore formation, which differs between groups of bacteria.
The ability to form endospores is found among bacteria in a number of genera, predominantly gram-positive groups, including the aerobic rod Bacillus, the microaerophilic rod Sporolactobacillus, the anaerobic rods Clostridium and Desulfotomaculum, the coccus Sporosarcina, and the filamentous Thermoactinomyces. The formation of a spore occurs in response to nutritional deprivation. Consequently, endospores do not possess metabolic activity until nutrients become available, at which time they are able to differentiate from spores into vegetative cells. Only one spore is formed inside each bacterial cell during sporulation. The formation of a spore begins with invagination of the cytoplasmic membrane around a copy of the bacterial chromosome, thus separating the contents of the smaller cell from the mother cell. The membrane of the mother cell engulfs the smaller cell within its cytoplasm, effectively providing two concentric unit membranes to protect the developing spore. A thin spore membrane and a thick cortex of a peptidoglycan are laid down between the two unit membranes. A rigid spore coat forms outside the cortex, enclosing the entire spore structure. The spore coat has keratin-like properties that are able to resist the lethal effects of heat, desiccation (dehydration), freezing, chemicals, and radiation. The ability of endospores to resist these noxious agents may ensue from the extremely low water content inside the spore. Methane-oxidizing bacteria in the genus Methylosinus also produce desiccation-resistant spores, called exospores.
Cysts are thick-walled structures produced by dormant members of Azotobacter, Bdellovibrio (bdellocysts), and Myxococcus (myxospores). They are resistant to desiccation and other harmful conditions but to a lesser degree than are endospores. In encystment by the nitrogen-fixing Azotobacter, cell division is followed by the formation of a thick, multilayered wall and coat that surround the resting cell. The filamentous actinomycetes (actinomycete) produce reproductive spores of two categories: conidiospores (conidium), which are chains of multiple spores formed on aerial or substrate mycelia (mycelium), or sporangiospores, which are formed in specialized sacs called sporangia.
Exchange of genetic information
Bacteria do not have an obligate sexual reproductive stage in their life cycle, but they can be very active in the exchange of genetic information. The genetic information carried in the DNA can be transferred from one cell to another; however, this is not a true exchange, because only one partner receives the new information. In addition, the amount of DNA that is transferred is usually only a small piece of the chromosome. There are several mechanisms by which this takes place. In transformation, bacteria take up free fragments of DNA that are floating in the medium. To take up the DNA efficiently, bacterial cells must be in a competent state, which is defined by the capability of bacteria to bind free fragments of DNA and is formed naturally only in a limited number of bacteria, such as Haemophilus, Neisseria, Streptococcus, and Bacillus. Many other bacteria, including E. coli, can be rendered competent artificially under laboratory conditions, such as by exposure to solutions of calcium chloride (CaCl2). transformation is a major tool in recombinant DNA technology, because fragments of DNA from one organism can be taken up by a second organism, thus allowing the second organism to acquire new characteristics.
transduction is the transfer of DNA from one bacterium to another by means of a bacteria-infecting virus called a bacteriophage. Transduction is an efficient means of transferring DNA between bacteria because DNA enclosed in the bacteriophage is protected from physical decay and from attack by enzymes in the environment and is injected directly into cells by the bacteriophage. However, widespread gene transfer by means of transduction is of limited significance because the packaging of bacterial DNA into a virus is inefficient and the bacteriophages are usually highly restricted in the range of bacterial species that they can infect. Thus, interspecies transfer of DNA by transduction is rare.
conjugation is the transfer of DNA by direct cell-to-cell contact that is mediated by plasmids (plasmid) (nonchromosomal DNA molecules). Conjugative plasmids encode an extremely efficient mechanism that mediates their own transfer from a donor cell to a recipient cell. The process takes place in one direction since only the donor cells contain the conjugative plasmid. In gram-negative bacteria, donor cells produce a specific plasmid-coded pilus, called the sex pilus, which attaches the donor cell to the recipient cell. Once connected, the two cells are brought into direct contact, and a conjugal bridge forms through which the DNA is transferred from the donor to the recipient. Many conjugative plasmids can be transferred between, and reproduce in, a large number of different gram-negative bacterial species. Some chemolithotrophic bacteria carry plasmids that are very large, ranging in size from 400,000 to 700,000 base pairs. The bacterial chromosome can also be transferred during conjugation, although this happens less frequently than plasmid transfer. Conjugation allows the inheritance of large portions of genes and may be responsible for the existence of bacteria with traits of several different species. Conjugation also has been observed in the gram-positive genus Enterococcus, but the mechanism of cell recognition and DNA transfer is different from that which occurs in gram-negative bacteria.
Growth of bacterial populations
Growth of bacterial cultures is defined as an increase in the number of bacteria in a population rather than in the size of individual cells. The growth of a bacterial population occurs in a geometric or exponential manner: with each division cycle (generation), one cell gives rise to 2 cells, then 4 cells, then 8 cells, then 16, then 32, and so forth. The time required for the formation of a generation, the generation time (G), can be calculated from the following formula:
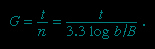
In the formula, B is the number of bacteria present at the start of the observation, b is the number present after the time period t, and n is the number of generations. The relationship shows that the mean generation time is constant and that the rate at which the number of bacteria increases is proportional to the number of bacteria at any given time. This relationship is valid only during the period when the population is increasing in an exponential manner, called the log phase of growth. For this reason, graphs that show the growth of bacterial cultures are plotted as the logarithm of the number of cells.
The generation time, which varies among bacteria, is controlled by many environmental conditions and by the nature of the bacterial species. For example, Clostridium perfringens, one of the fastest-growing bacteria, has an optimum generation time of about 10 minutes; Escherichia coli can double every 20 minutes; and the slow-growing Mycobacterium tuberculosis has a generation time in the range of 12 to 16 hours. The composition of the growth medium is a major factor controlling the growth rate. The growth rate increases up to a maximum when the medium provides a better energy source and more of the biosynthetic intermediates that the cell would otherwise have to make for itself.
 When bacteria are placed in a medium that provides all of the nutrients that are necessary for their growth, the population exhibits four phases of growth that are representative of a typical bacterial growth curve. Upon inoculation into the new medium, bacteria do not immediately reproduce, and the population size remains constant. During this period, called the lag phase, the cells are metabolically active and increase only in cell size. They are also synthesizing the enzymes and factors needed for cell division and population growth under their new environmental conditions. The population then enters the log phase, in which cell numbers increase in a logarithmic fashion, and each cell generation occurs in the same time interval as the preceding ones, resulting in a balanced increase in the constituents of each cell. The log phase continues until nutrients are depleted or toxic products accumulate, at which time the cell growth rate slows, and some cells may begin to die. Under optimum conditions, the maximum population for some bacterial species at the end of the log phase can reach a density of 10 to 30 billion cells per millilitre.
When bacteria are placed in a medium that provides all of the nutrients that are necessary for their growth, the population exhibits four phases of growth that are representative of a typical bacterial growth curve. Upon inoculation into the new medium, bacteria do not immediately reproduce, and the population size remains constant. During this period, called the lag phase, the cells are metabolically active and increase only in cell size. They are also synthesizing the enzymes and factors needed for cell division and population growth under their new environmental conditions. The population then enters the log phase, in which cell numbers increase in a logarithmic fashion, and each cell generation occurs in the same time interval as the preceding ones, resulting in a balanced increase in the constituents of each cell. The log phase continues until nutrients are depleted or toxic products accumulate, at which time the cell growth rate slows, and some cells may begin to die. Under optimum conditions, the maximum population for some bacterial species at the end of the log phase can reach a density of 10 to 30 billion cells per millilitre.The log phase of bacterial growth is followed by the stationary phase, in which the size of a population of bacteria remains constant, even though some cells continue to divide and others begin to die. The stationary phase is followed by the death phase, in which the death of cells in the population exceeds the formation of new cells. The length of time before the onset of the death phase depends on the species and the medium. Bacteria do not necessarily die even when starved of nutrients, and they can remain viable for long periods of time.
Ecology of bacteria
Distribution in nature
Prokaryotes are ubiquitous on the Earth's surface. They are found in every accessible environment, from polar ice to bubbling hot springs (hot spring), from mountaintops to the ocean floor, and from plant and animal bodies to forest soils. Some bacteria can grow in soil or water at temperatures near freezing (0 °C 【32 °F】), whereas others thrive in water at temperatures near boiling (100 °C 【212 °F】). Each bacterium is adapted to live in a particular environmental niche, be it oceanic surfaces, mud sediments, soil, or the surfaces of another organism. The level of bacteria in the air is low but significant, especially when dust has been suspended. In uncontaminated natural bodies of water, bacterial counts can be in the thousands per millilitre; in fertile soil, bacterial counts can be in the millions per gram; and in feces, bacterial counts can exceed billions per gram.
Prokaryotes are important members of their habitats. Although they are small in size, their sheer numbers mean that their metabolism plays an enormous role—sometimes beneficial, sometimes harmful—in the conversion of elements in their external environment. Probably every naturally occurring substance, and many synthetic ones, can be degraded (metabolized) by some species of bacteria (often members of the aerobic Pseudomonas groups). The largest stomach of the cow (cattle), the rumen, is a fermentation chamber in which bacteria digest the cellulose in grasses and feeds, converting them to fatty acids (fatty acid) and amino acids (amino acid), which are the fundamental nutrients used by the cow and the basis for the cow's production of milk. Organic wastes in sewage or compost piles are converted by bacteria either into suitable nutrients for plant metabolism or into gaseous methane (CH4) and carbon dioxide. The remains of all organic materials, including plants and animals, are eventually converted to soil and gases through the activities of bacteria and other microorganisms and are thereby made available for further growth.
Many bacteria live in streams and other sources of water (water purification), and their presence at low population densities in a sample of water does not necessarily indicate that the water is unfit for consumption. However, water that contains bacteria such as E. coli, which are normal inhabitants of the intestinal tract (intestine) of humans and animals, indicates that sewage or fecal material has recently polluted that water source. Such coliform bacteria may be pathogens (disease-causing organisms) themselves, and their presence signals that other, less easily detected bacterial and viral pathogens may also be present. Procedures used in water purification plants— settling, filtration, and chlorination—are designed to remove these and any other microorganisms and infectious agents (infectious disease) that may be present in water that is intended for human consumption. Also, sewage treatment is necessary to prevent the release of pathogenic bacteria and viruses from wastewater into water supplies. Sewage treatment plants also initiate the decay of organic materials (proteins, fats, and carbohydrates) in the wastewater. The breakdown of organic material by microorganisms in the water consumes oxygen ( biochemical oxygen demand), causing a decrease in the oxygen level, which can be very harmful to aquatic life in streams and lakes that receive the wastewater. One objective of sewage treatment is to oxidize as much organic material as possible before its discharge into the water system, thereby reducing the biochemical oxygen demand of the wastewater. Sewage digestion tanks and aeration devices specifically exploit the metabolic capacity of bacteria for this purpose. (For more information about the treatment of wastewater, see environmental works: Water-pollution control (environmental works).)
 Soil bacteria are extremely active in effecting biochemical changes by transforming the various substances, humus and minerals, that characterize soil. Elements that are central to life, such as carbon, nitrogen, and sulfur, are converted by bacteria from inorganic gaseous compounds into forms that can be used by plants and animals. Bacteria also convert the end products of plant and animal metabolism into forms that can be used by bacteria and other microorganisms. The nitrogen cycle can illustrate the role of bacteria in effecting various chemical changes. Nitrogen exists in nature in several oxidation states, as nitrate, nitrite, dinitrogen gas, several nitrogen oxides, ammonia, and organic amines (amine) (ammonia compounds containing one or more substituted hydrocarbons (hydrocarbon)). nitrogen fixation is the conversion of dinitrogen gas from the atmosphere into a form that can be used by living organisms. Some nitrogen-fixing bacteria, such as Azotobacter, Clostridium pasteurianum (Clostridium), and Klebsiella pneumoniae (Klebsiella), are free-living, whereas species of Rhizobium live in an intimate association with leguminous plants (legume). Rhizobium organisms in the soil recognize and invade the root hairs of their specific plant host, enter the plant tissues, and form a root nodule. This process causes the bacteria to lose many of their free-living characteristics. They become dependent upon the carbon supplied by the plant, and, in exchange for carbon, they convert nitrogen gas to ammonia, which is used by the plant for its protein synthesis and growth. In addition, many bacteria can convert nitrate to amines for purposes of synthesizing cellular materials or to ammonia when nitrate is used as electron acceptor. denitrifying bacteria convert nitrate to dinitrogen gas. The conversion of ammonia or organic amines to nitrate is accomplished by the combined activities of the aerobic organisms Nitrosomonas and Nitrobacter, which use ammonia as an electron donor.
Soil bacteria are extremely active in effecting biochemical changes by transforming the various substances, humus and minerals, that characterize soil. Elements that are central to life, such as carbon, nitrogen, and sulfur, are converted by bacteria from inorganic gaseous compounds into forms that can be used by plants and animals. Bacteria also convert the end products of plant and animal metabolism into forms that can be used by bacteria and other microorganisms. The nitrogen cycle can illustrate the role of bacteria in effecting various chemical changes. Nitrogen exists in nature in several oxidation states, as nitrate, nitrite, dinitrogen gas, several nitrogen oxides, ammonia, and organic amines (amine) (ammonia compounds containing one or more substituted hydrocarbons (hydrocarbon)). nitrogen fixation is the conversion of dinitrogen gas from the atmosphere into a form that can be used by living organisms. Some nitrogen-fixing bacteria, such as Azotobacter, Clostridium pasteurianum (Clostridium), and Klebsiella pneumoniae (Klebsiella), are free-living, whereas species of Rhizobium live in an intimate association with leguminous plants (legume). Rhizobium organisms in the soil recognize and invade the root hairs of their specific plant host, enter the plant tissues, and form a root nodule. This process causes the bacteria to lose many of their free-living characteristics. They become dependent upon the carbon supplied by the plant, and, in exchange for carbon, they convert nitrogen gas to ammonia, which is used by the plant for its protein synthesis and growth. In addition, many bacteria can convert nitrate to amines for purposes of synthesizing cellular materials or to ammonia when nitrate is used as electron acceptor. denitrifying bacteria convert nitrate to dinitrogen gas. The conversion of ammonia or organic amines to nitrate is accomplished by the combined activities of the aerobic organisms Nitrosomonas and Nitrobacter, which use ammonia as an electron donor.In the carbon cycle, carbon dioxide is converted into cellular materials by plants and autotrophic prokaryotes, and organic carbon is returned to the atmosphere by heterotrophic life-forms. The major breakdown product of microbial decomposition is carbon dioxide, which is formed by respiring aerobic organisms.
methane, another gaseous end product of carbon metabolism, is a relatively minor component of the global carbon cycle but of importance in local situations and as a renewable energy source for human use. Methane production is carried out by the highly specialized and obligately anaerobic methanogenic prokaryotes, all of which are archaea. Methanogens use carbon dioxide as their terminal electron acceptor and receive electrons from hydrogen gas (H2). A few other substances can be converted to methane by these organisms, including methanol, formic acid, acetic acid, and methylamines. Despite the extremely narrow range of substances that can be used by methanogens, methane production is very common during the anaerobic decomposition of many organic materials, including cellulose, starch, proteins, amino acids, fats, alcohols (alcohol), and most other substrates. Methane formation from these materials requires that other anaerobic bacteria degrade these substances either to acetate or to carbon dioxide and hydrogen gas, which are then used by the methanogens. The methanogens support the growth of the other anaerobic bacteria in the mixture by removing hydrogen gas formed during their metabolic activities for methane production. Consumption of the hydrogen gas stimulates the metabolism of other bacteria.
Despite the fact that methanogens have such a restricted metabolic capability and are quite sensitive to oxygen, they are widespread on Earth. Large amounts of methane are produced in anaerobic environments, such as swamps (swamp) and marshes (marsh), but significant amounts also are produced in soil and by ruminant animals. At least 80 percent of the methane in the atmosphere has been produced by the action of methanogens, the remainder being released from coal deposits or natural gas wells.
The importance of bacteria to humans
Bacteria in food
milk from a healthy cow initially contains very few bacteria, which primarily come from the skin of the cow and the procedures for handling the milk. Milk is an excellent growth medium for numerous bacteria, and the bacteria can increase rapidly in numbers unless the milk is properly processed. Bacterial growth can spoil the milk or even pose a serious health hazard if pathogenic bacteria are present. Diseases that can be transmitted from an infected cow include tuberculosis (Mycobacterium tuberculosis), undulant fever (brucellosis) (Brucella abortus), and Q fever (Coxiella burnetii). In addition, typhoid fever (Salmonella typhi) can be transmitted through milk from an infected milk handler. pasteurization procedures increase the temperature of the milk to 63 °C (145 °F) for 30 minutes or to 71 °C (160 °F) for 15 seconds, which kills any of the pathogenic bacteria that might be present, although these procedures do not kill all microorganisms.
Certain bacteria convert milk into useful dairy products, such as buttermilk, yogurt, and cheese. Commercially cultured buttermilk is prepared from skim milk inoculated with a starter culture of Streptococcus lactis or S. cremoris, together with Leuconostoc citrovorum or L. dextranicum. The combined action of Streptococcus and Leuconostoc consumes the milk sugar, produces lactic acid, and precipitates milk protein (casein). yogurt and other fermented milk products are produced in a similar manner using different cultures of bacteria. Many cheeses (cheese) are likewise made through the action of bacteria. Growth in milk of an acid-producing bacterium such as S. lactis causes the casein to precipitate as curd. Following the removal of moisture and the addition of salt, the curd is allowed to ripen through the action of other microorganisms. Lactobacilli (Lactobacillus), streptococci, and propionibacteria are important for the ripening of Swiss cheese (Emmentaler) and the production of its characteristic taste and large gas bubbles. In addition, Brevibacterium linens is responsible for the flavour of Limburger cheese, and molds (mold) (Penicillium species) are used in the manufacture of Roquefort and Camembert cheeses (Camembert cheese). Other types of bacteria have long been used in the preparation and preservation of various foods produced through bacterial fermentation, including pickled products, sauerkraut, and olives (olive).
Substances contributing to the virulence of pathogenic bacteria Substances contributing to the virulence of pathogenic bacteriaThe toxins of many pathogenic bacteria that are transmitted in foods can cause food poisoning when ingested. These include a toxin produced by Staphylococcus aureus, which causes a rapid, severe, but limited gastrointestinal distress, or the toxin of Clostridium botulinum, which is often lethal. Production of botulism toxin can occur in canned nonacidic foods that have been incompletely cooked before sealing. C. botulinum forms heat-resistant spores that can germinate into vegetative bacterial cells that thrive in the anaerobic environment, which is conducive to the production of their extremely potent toxin. Other food-borne infections are actually transmitted from an infected food handler, including typhoid fever, salmonellosis ( Salmonella species), and shigellosis (Shigella dysenteriae).
Bacteria in industry
Some compounds produced by bacteria on an industrial scale Some compounds produced by bacteria on an industrial scaleAnaerobic sugar fermentation reactions by various bacteria produce different end products. The production of ethanol by yeasts (yeast) has been exploited by the brewing industry for thousands of years and is used for fuel production. Specific bacteria carry out the oxidation of alcohol to acetic acid in the production of vinegar. Other fermentation processes make even more valuable products. Organic compounds, such as acetone, isopropanol, and butyric acid, are produced in fermentation by various Clostridium species and can been prepared on an industrial scale. Other bacterial products and reactions have been discovered in organisms from extreme environments. There is considerable interest in the enzymes isolated from thermophilic bacteria, in which reactions may be carried out at higher rates owing to the higher temperatures at which they can occur.
Deposits of insoluble ferric iron (iron in the +3 oxidation state) are common in many environments. Bacterial reduction of ferric iron is common in waterlogged soils, bogs, and anaerobic portions of lakes. When the soluble ferrous iron (iron in the +2 oxidation state) thus formed reaches aerobic regions under neutral pH, the ferrous iron spontaneously oxidizes to insoluble, brown ferric deposits. In an acidic environment, iron does not readily oxidize from the ferrous to the ferric state. However, this reaction is tremendously accelerated by the acidophilic lithotrophic bacterium Thiobacillus ferrooxidans. When pyritic (pyrite) (ferrous sulfide) deposits are exposed to the air by mining operations, there is slow spontaneous oxidation of pyrite to ferrous ions and sulfuric acid. When the production of sulfuric acid causes conditions to reach a certain level of acidity, T. ferrooxidans thrives and oxidizes ferrous iron to the ferric form, which in turn oxidizes more pyrite in a continuously increasing fashion, with the formation of substantial amounts of sulfuric acid. The acidity of the environment may increase to a level near a pH of 2, which is a better environment for the solubilization of many other metal ions, particularly aluminum. Some of the ferrous iron generated by the bacteria is carried away by groundwater into surrounding streams, making them acidic and loaded with iron, which precipitates and forms deposits of iron some distance downstream from the mine. Acid mine drainage would not develop in the absence of bacterial activity, and the only practical way to prevent its occurrence is to seal or cover the acid-bearing material to prevent exposure to air. (For more information of oxidation and reduction reactions, see oxidation-reduction reaction (oxidation–reduction reaction)).
Although bacterial oxidation of sulfide materials results in the undesirable formation of acid mine drainage, the same reaction has been put to use for microbial leaching of copper, uranium, and other valuable metals from low-grade sulfide-containing ores (ore). These metals are released from the ore after their conversion to more soluble forms by the direct oxidation of the metal by the bacterium and by the indirect oxidation of the metals in the ore by the ferric iron that was formed by bacterial action.
Microbial decomposition of petroleum products by hydrocarbon-oxidizing bacteria and fungi is of considerable ecological importance. The microbial decomposition of petroleum is an aerobic process, which is prevented if the oil settles to the layer of anaerobic sediment at the bottom (natural oil deposits in anaerobic environments are millions of years old). Hydrocarbon-oxidizing bacteria attach to floating oil droplets on the water surface, where their action eventually decomposes the oil to carbon dioxide. It is becoming a common practice to spray such bacteria and their growth factors onto oil spills (oil spill) to enhance the rate of degradation of the nonvolatile aliphatic (aliphatic compound) and aromatic (aromatic compound) hydrocarbons.
Bacteria in medicine
 Types of bacteria that cause disease in humans Types of bacteria that cause disease in humansBacterial diseases have played a dominant role in human history. Widespread epidemics (epidemic) of cholera and plague reduced populations of humans in some areas of the world by more than one-third. Bacterial pneumonia was probably the major cause of death in the aged. Perhaps more armies were defeated by typhus, dysentery, and other bacterial infections than by force of arms. With modern advances in plumbing and sanitation, the development of bacterial vaccines (vaccine), and the discovery of antibacterial antibiotics (antibiotic), the incidence of bacterial disease has been reduced. Bacteria have not disappeared as infectious agents, however, since they continue to evolve, creating increasingly virulent strains and acquiring resistance to many antibiotics.
Types of bacteria that cause disease in humans Types of bacteria that cause disease in humansBacterial diseases have played a dominant role in human history. Widespread epidemics (epidemic) of cholera and plague reduced populations of humans in some areas of the world by more than one-third. Bacterial pneumonia was probably the major cause of death in the aged. Perhaps more armies were defeated by typhus, dysentery, and other bacterial infections than by force of arms. With modern advances in plumbing and sanitation, the development of bacterial vaccines (vaccine), and the discovery of antibacterial antibiotics (antibiotic), the incidence of bacterial disease has been reduced. Bacteria have not disappeared as infectious agents, however, since they continue to evolve, creating increasingly virulent strains and acquiring resistance to many antibiotics.Although most bacteria are beneficial or even necessary for life on Earth, a few are known for their detrimental impact on humans. None of the archaea are currently considered to be pathogens, but animals, including humans, are constantly bombarded and inhabited by large numbers and varieties of Bacteria. Most bacteria that contact an animal are rapidly eliminated by the host's defenses. The oral cavities, intestinal tract, and skin are colonized by enormous numbers of specific types of bacteria that are adapted to life in those habitats. These organisms are harmless under normal conditions and become dangerous only if they somehow pass across the barriers of the body and cause infection. Some bacteria are adept at invasion of a host and are called pathogens, or disease producers. Some pathogens act at specific parts of the body, such as meningococcal (meningococcus) bacteria (Neisseria meningitidis), which invade and irritate the meninges, the membranes surrounding the brain and spinal cord; the diphtheria bacterium (Corynebacterium diphtheriae), which initially infects the throat; and the cholera bacterium (Vibrio cholerae), which reproduces in the intestinal tract, where the toxin that it produces causes the voluminous diarrhea characteristic of this cholera. Other bacteria that can infect humans include staphylococcal bacteria (primarily Staphylococcus aureus), which can infect the skin to cause boils (boil) (furuncles), the bloodstream to cause septicemia (blood poisoning), the heart valves to cause endocarditis, or the bones to cause osteomyelitis.

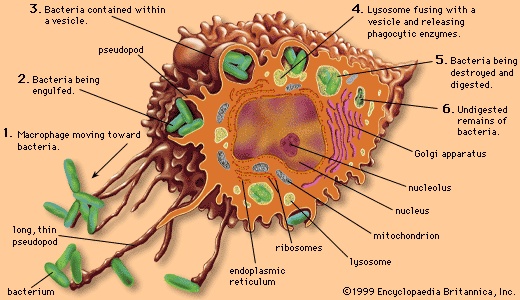 Pathogenic bacteria that invade an animal's bloodstream can use any of a number of mechanisms to evade the host's immune system, including the formation of long lipopolysaccharide chains to provide resistance to a group of serum immune proteins, called complement, that normally retard the bacterium. The pathogenic restructuring of bacterial surface proteins prevents antibodies produced by the animal from recognizing the pathogen and in some cases gives the pathogen the ability to survive and grow in phagocytic white blood cells (leukocyte). Many pathogenic bacteria produce toxins that assist them in invading the host. Among these toxins are proteases (proteolytic enzyme), enzymes that break down tissue proteins, and lipases (lipase), enzymes that break down lipid (fat) and damage cells by disrupting their membranes. Other toxins disrupt cell membranes by forming a pore or channel in them. Some toxins are enzymes that modify specific proteins involved in protein synthesis or in control of host cell metabolism; examples include the diphtheria, cholera, and pertussis (whooping cough) toxins.
Pathogenic bacteria that invade an animal's bloodstream can use any of a number of mechanisms to evade the host's immune system, including the formation of long lipopolysaccharide chains to provide resistance to a group of serum immune proteins, called complement, that normally retard the bacterium. The pathogenic restructuring of bacterial surface proteins prevents antibodies produced by the animal from recognizing the pathogen and in some cases gives the pathogen the ability to survive and grow in phagocytic white blood cells (leukocyte). Many pathogenic bacteria produce toxins that assist them in invading the host. Among these toxins are proteases (proteolytic enzyme), enzymes that break down tissue proteins, and lipases (lipase), enzymes that break down lipid (fat) and damage cells by disrupting their membranes. Other toxins disrupt cell membranes by forming a pore or channel in them. Some toxins are enzymes that modify specific proteins involved in protein synthesis or in control of host cell metabolism; examples include the diphtheria, cholera, and pertussis (whooping cough) toxins.Some pathogenic bacteria form areas in the host's body where they are closed off and protected from the immune system, as occurs in the boils in the skin formed by staphylococci and the cavities in the lungs formed by Mycobacterium tuberculosis. Bacteroides fragilis is the most numerous inhabitant of the human intestinal tract and causes no difficulties for the host as long as it remains there. If this bacterium gets into the body by means of an injury, the bacterial capsule stimulates the body to wall off the bacteria into an abscess, which reduces the spread of the bacteria. In many instances, the symptoms of bacterial infections are actually the result of an excessive response by the immune system rather than of the production of toxic factors by the bacterium.
Other means of combating pathogenic bacterial infections include the use of biotherapeutic agents, or probiotics. These are harmless bacteria that interfere with the colonization by pathogenic bacteria. Another approach employs bacteriophages, viruses that kill bacteria, for the treatment of infections by specific bacterial pathogens. In addition, recombinant DNA technologies (recombinant DNA technology), developed during the 1980s, have made it possible to synthesize nearly any protein in bacteria, with E. coli serving as the usual host organism in this process. Recombinant DNA technology is used for the inexpensive, large-scale production of extremely scarce and valuable animal or human proteins, such as hormones (hormone), blood-clotting factors, and even antibodies (antibody).
Evolution of bacteria
Bacteria have existed from very early in the history of life on Earth. Bacteria fossils (fossil) discovered in rocks date from at least the Devonian Period (416 to 359.2 million years ago), and there are convincing arguments that bacteria have been present since early Precambrian time, about 3.5 billion years ago. Bacteria were widespread on Earth at least since the middle of the Proterozoic Eon, about 1.5 billion years ago, when oxygen appeared in the atmosphere as a result of the action of the cyanobacteria (blue-green algae). Bacteria have thus had plenty of time to adapt to their environments and to have given rise to numerous descendant forms.
The nature of the original predecessor involved in the origin of life is subject to considerable speculation. It has been suggested that the original cell might have used RNA as its genetic material, since investigations have shown that RNA molecules can have numerous catalytic functions. The Bacteria and archaea diverged from their common precursor very early in this time period. The two types of prokaryotes tend to inhabit different types of environments and give rise to new species at different rates. Many Archaea prefer high-temperature niches. One major branch of the archaeal tree consists only of thermophilic species, and many of the methanogens in another major branch can grow at high temperatures. In contrast, no major eubacterial (eubacterium) branch consists solely of thermophiles. Both Bacteria and Archaea contain members that are able to grow at very high temperatures, as well as other species that are able to grow at low temperatures. Another prominent difference is that bacteria have widely adapted to aerobic conditions, whereas many archaea are obligate anaerobes. No archaea are obligately photosynthetic. Perhaps the archaea are a more primitive type of organism with an impaired genetic response to changing environmental conditions. A limited ability to adapt to new situations could restrict the archaea to harsh environments, where there is less competition from other life-forms.
Organisms must evolve or adapt to changing environments, and it is clear that mutations (mutation), which are changes in the sequence of nucleotides (nucleotide) in an organism's DNA, occur constantly in all organisms. The changes in DNA sequence might result in changes in the amino acid sequence of the protein that is encoded by that stretch of DNA. As a result, the altered protein might be either better-suited or less well-suited for function under the prevailing conditions. Although many nucleotide changes that can occur in DNA have no effect on the fitness of the cell, if the nucleotide change enhances the growth of that cell even by a small degree, then the mutant form would be able to increase its relative numbers in the population. If the nucleotide change retards the growth of the cell, however, then the mutant form would be outgrown by the other cells and lost.
The ability to transfer genetic information between organisms is a major factor in adaptation to changes in environment. The exchange of DNA is an essential part of the life cycle of higher eukaryotic organisms (eukaryote) and can occur in all eukaryotes. Genetic exchange occurs throughout the bacterial world as well, and, although the amount of DNA that is transferred is small, this transfer can occur between distantly related organisms. Genes carried on plasmids (plasmid) can find their way onto the bacterial chromosome and become a stable part of the bacterium's inheritance. Organisms usually possess mobile genetic elements called transposons that can rearrange the order and presence of any genes on the chromosome. Transposons may play a role in helping to accelerate the pace of evolution.
Many examples of the rapid evolution of bacteria are available. Before the 1940s, antibiotics (antibiotic) were not used in medical practice. When antibiotics did eventually come into use, the majority of pathogenic bacteria were sensitive to them. Since then, however, the bacterial resistance to one or more antibiotics has increased to the point that previously effective antibiotics are no longer useful against certain types of bacteria. Most examples of antibiotic resistance in pathogenic bacteria are not the result of a mutation that alters the protein that the antibiotic attacks, although this mechanism can occur. Instead, antibiotic resistance often involves the production by the bacterium of enzymes that alter the antibiotic and render it inactive. The major factor in the spread of antibiotic resistance is transmissible plasmids, which carry the genes for the drug-inactivating enzymes from one bacterial species to another. Although the original source of the gene for these enzymes is not known, mobile genetic elements (transposons) may have played a role in their appearance and may also allow their transfer to other bacterial types.
Biosynthesis, nutrition, and growth of bacteria
Factors affecting bacterial growth
Nutritional requirements
Bacteria differ dramatically with respect to the conditions that are necessary for their optimal growth. In terms of nutritional needs, all cells require sources of carbon, nitrogen, sulfur, phosphorus, numerous inorganic salts (e.g., potassium, magnesium, sodium, calcium, and iron), and a large number of other elements called micronutrients (e.g., zinc, copper, manganese, selenium, tungsten, and molybdenum). Carbon is the element required in the greatest amount by bacteria since hydrogen and oxygen can be obtained from water, which is a prerequisite for bacterial growth. Also required is a source of energy to fuel the metabolism of the bacterium. One means of organizing bacteria is based on these fundamental nutritional needs: the carbon source and the energy source.
There are two sources a cell can use for carbon: inorganic compounds (inorganic compound) and organic compounds (organic compound). Organisms that use the inorganic compound carbon dioxide (CO2) as their source of carbon are called autotrophs. Bacteria that require an organic source of carbon, such as sugars, proteins (protein), fats (fat), or amino acids (amino acid), are called heterotrophs (or organotrophs). Many heterotrophs, such as Escherichia coli or Pseudomonas aeruginosa, synthesize all of their cellular constituents from simple sugars such as glucose because they possess the necessary biosynthetic pathways. Other heterotrophs have lost some of these biosynthetic pathways; in order to grow, they require that their environments contain particular amino acids, nitrogenous bases, or vitamins that are chemically intact.
In addition to carbon, bacteria need energy, which is almost always obtained by the transfer of an electron from an electron donor to an electron acceptor. There are three basic sources of energy: light, inorganic compounds, and organic compounds. Phototrophic bacteria use photosynthesis to generate cellular energy in the form of adenosine triphosphate (ATP) from light energy. Chemotrophs obtain their energy from chemicals (organic and inorganic compounds); chemolithotrophs obtain their energy from reactions with inorganic salts; and chemoheterotrophs obtain their carbon and energy from organic compounds (the energy source may also serve as the carbon source in these organisms).
In most cases, cellular energy is generated by means of electron-transfer reactions, in which electrons move from an organic or inorganic donor molecule to an acceptor molecule via a pathway that conserves the energy released during the transfer of electrons by trapping it in a form that the cell can use for its chemical or physical work. The primary form of energy that is captured from the transfer of electrons is ATP. The metabolic processes that break down organic molecules to generate energy are called catabolic reactions (catabolism). In contrast, the metabolic processes that synthesize molecules are called anabolic reactions (anabolism).
Many bacteria can use a large number of compounds as carbon and energy sources, whereas other bacteria are highly restricted in their metabolic capabilities. While carbohydrates (carbohydrate) are a common energy source for eukaryotes, these molecules are metabolized by only a limited number of species of bacteria, since most bacteria do not possess the necessary enzymes to metabolize these often complex molecules. Many species of bacteria instead depend on other energy sources, such as amino acids, fats, or other compounds. Other compounds of significance to bacteria include phosphate, sulfate, and nitrogen. Low levels of phosphate in many environments, particularly in water, can be a limiting factor for the growth of bacteria, since many bacteria cannot synthesize phosphate. Most bacteria can convert sulfate or sulfide to the organic form needed for protein synthesis. The capability of a living organism to incorporate nitrogen from ammonia is widespread in nature, and bacteria differ in their ability to convert other forms of nitrogen, such as nitrate in the soil or dinitrogen gas (N2) in the atmosphere, into cell material.
A particularly important nutrient of bacteria is iron, an abundant element in the Earth's crust (crust). Iron is a component of heme proteins, such as hemoglobin in red blood cells and cytochromes (cytochrome) in electron transfer chains, as well as many other iron-containing proteins involved in electron-transfer reactions. Iron is needed for the growth of almost all organisms. In aerobic environments at neutral pH values, ferrous iron (iron in the +2 state) is oxidized to ferric iron (iron in the +3 state), which is virtually insoluble in water and unable to enter cells. Many bacteria synthesize and secrete chemicals called siderophores that bind very tightly to iron and make it soluble in water. The bacteria then take up these iron-siderophore complexes and remove the iron for their synthetic tasks. The ability to acquire iron in this way is particularly important to pathogenic (disease-causing) bacteria, which must compete with their host for iron. In anaerobic environments, iron can exist in the more soluble ferrous state and is readily available to bacteria.
Some bacteria are obligate parasites and grow only within a living host cell. rickettsia and Chlamydia, for example, grow in eukaryotic cells, and Bdellovibrio grow in bacterial cells. Treponema pallidum is difficult, if not impossible, to grow in culture, probably because it requires low oxygen tension and low oxidation-reduction levels, which are provided by the presence of animal cells, rather than any specific nutrient. Because some bacteria may thrive only as animal or plant parasites or only in a rich source of nutrients such as milk, they likely do not thrive as free bacteria in nature. Many bacteria from natural environments exist in a consortium with other bacteria and are difficult to isolate and culture separately from the other members of that partnership.
Physical requirements
The physical requirements that are optimal for bacterial growth vary dramatically for different bacterial types. As a group, bacteria display the widest variation of all organisms in their ability to inhabit different environments. Some of the most prominent factors are described in the following sections.
oxygen
One of the most prominent differences between bacteria is their requirement for, and response to, atmospheric oxygen (O2). Whereas essentially all eukaryotic organisms require oxygen to thrive, many species of bacteria can grow under anaerobic (aerobe) conditions. Bacteria that require oxygen to grow are called obligate aerobic bacteria. In most cases, these bacteria require oxygen to grow because their methods of energy production and respiration depend on the transfer of electrons to oxygen, which is the final electron acceptor in the electron transport reaction. Obligate aerobes include Bacillus subtilis, Pseudomonas aeruginosa, Mycobacterium tuberculosis, and Thiobacillus ferrooxidans.
Bacteria that grow only in the absence of oxygen, such as Clostridium, Bacteroides, and the methane-producing archaea (methanogens), are called obligate anaerobes because their energy-generating metabolic processes are not coupled with the consumption of oxygen. In fact, the presence of oxygen actually poisons some of their key enzymes. Some bacteria (S. pneumoniae) are microaerophilic or aerotolerant anaerobes because they grow better in low concentrations of oxygen. In these bacteria, oxygen often stimulates minor metabolic processes that enhance the major routes of energy production. Facultative anaerobes can change their metabolic processes depending on the presence of oxygen, using the more efficient process of respiration (cellular respiration) in the presence of oxygen and the less efficient process of fermentation in the absence of oxygen. Examples of facultative anaerobes include E. coli and S. aureus.
The response of bacteria to oxygen is not determined simply by their metabolic needs. Oxygen is a very reactive molecule and forms several toxic by-products, such as superoxide (O2−.), hydrogen peroxide (H2O2), and the hydroxyl radical (OH·). Aerobic organisms produce enzymes that detoxify these oxygen products. The most common of detoxifying enzymes are catalase, which breaks down hydrogen peroxide, and superoxide dismutase, which breaks down superoxide. The combined action of these enzymes to remove hydrogen peroxide and superoxide is important because these by-products together with iron form the extremely reactive hydroxyl radical, which is capable of killing the cell. Anaerobic bacteria generally do not produce catalase, and their levels of superoxide dismutase vary in rough proportion with the cell's sensitivity to oxygen. Many anaerobes are hypersensitive to oxygen, being killed upon short exposure, whereas other anaerobes, including most Clostridium species, are more tolerant to the presence of oxygen.
temperature
Bacteria have adapted to a wide range of temperatures. Bacteria that grow at temperatures of less than about 15 °C (59 °F) are psychrophiles. The ability of bacteria to grow at low temperatures is not unexpected, since the average subsurface temperature of soil in the temperate zone is about 12 °C (54 °F) and 90 percent of the oceans measure 5 °C (41 °F) or colder. Obligate psychrophiles, which have been isolated from Arctic and Antarctic ocean waters and sediments, have optimum growth temperatures of about 10 °C (50 °F) and do not survive if exposed to 20 °C (68 °F). The majority of psychrophilic bacteria are in the gram-negative genera Pseudomonas, Flavobacterium, Achromobacter, and Alcaligenes. Mesophilic bacteria are those in which optimum growth occurs between 20 and 45 °C (68 and 113 °F), although they usually can survive and grow in temperatures between 10 and 50 °C (50 and 122 °F). Animal pathogens are mesophiles.
Thermophilic prokaryotes can grow at temperatures higher than 60 °C (140 °F). These temperatures are encountered in rotting compost piles, hot springs (hot spring), and oceanic geothermal vents. In the runoff of a hot spring, thermophiles such as the bacterium Thermus aquaticus (optimum temperature for growth, 70 °C 【158 °F】; maximum temperature, 79 °C 【174 °F】) are found near the source where the temperature has fallen to about 70 °C. Thick mats of the cyanobacterium Synechococcus and the phototrophic gliding bacterium Chloroflexus develop in somewhat cooler portions of the runoff. The archaeon Sulfolobus acidocaldarius has a high tolerance for acidic conditions, which allows growth in a pH range of about 1.0 to 6.0 and a temperature optimum of 70 to 75 °C (158 to 167 °F). Numerous bacteria and archaea are adapted to the temperature range of 50 to 70 °C (122 to 158 °F), including some members of the genera Bacillus, Thermoactinomyces, Methanobacterium, Methylococcus, and Sulfolobus. Most striking was the discovery in the mid-1980s of bacteria and archaea in nutrient-rich, extremely hot hydrothermal vents on the deep seafloor. The archaea in the genus Pyrodictium thrive in the temperature range of 80 to 110 °C (176 to 230 °F), temperatures at which the water remains liquid only because of the extremely high pressures.
pH
Most bacteria grow in the range of neutral pH values (between 5 and 8), although some species have adapted to life at more acidic (acid) or alkaline (alkali) extremes. An example of an acidophilic bacterium is T. ferrooxidans. When coal seams are exposed to air through mining operations, the pyritic (pyrite) ferrous sulfide deposits are attacked by T. ferrooxidans to generate sulfuric acid, which lowers the pH to 2.0 or even 0.7. However, acid tolerance of T. ferrooxidans applies only to sulfuric acid, since these bacteria die when exposed to equivalent concentrations of other acids such as hydrochloric acid. Many bacteria cannot tolerate acidic environments, especially under anaerobic conditions, and, as a result, plant polymers degrade slowly in acidic (pH between 3.7 and 5.5) bogs, pine forests, and lakes. In contrast to acidophilic bacteria, alkalophilic bacteria are able to grow in alkaline concentrations as great as pH 10 to 11. Alkalophiles have been isolated from soils, and most are species of the gram-positive genus Bacillus.
Salt and water
 Water is a fundamental requirement for life. Some bacteria prefer salty environments and are thus called halophiles. Extreme halophiles, such as Halobacterium, show optimum growth in conditions of 20 to 30 percent salt and will lyse (break open) if this salt level is reduced. Such bacteria are found in the Dead Sea, in brine ponds, and occasionally on salted fishes and hides. Moderately halophilic bacteria grow in conditions of 5 to 20 percent salt and are found in salt brines and muds.
Water is a fundamental requirement for life. Some bacteria prefer salty environments and are thus called halophiles. Extreme halophiles, such as Halobacterium, show optimum growth in conditions of 20 to 30 percent salt and will lyse (break open) if this salt level is reduced. Such bacteria are found in the Dead Sea, in brine ponds, and occasionally on salted fishes and hides. Moderately halophilic bacteria grow in conditions of 5 to 20 percent salt and are found in salt brines and muds.Bacterial metabolism
Heterotrophic metabolism
As stated above, heterotrophic (or organotrophic) bacteria require organic molecules to provide their carbon and energy. The energy-yielding catabolic (catabolism) reactions can be of many different types, although they all involve electron-transfer reactions in which the movement of an electron from one molecule to another is coupled with an energy-trapping reaction that yields ATP (adenosine triphosphate). Some heterotrophic bacteria can metabolize sugars or complex carbohydrates (carbohydrate) to produce energy. These bacteria must produce a number of specific proteins, including enzymes that degrade the polysaccharides (polysaccharide) into their constituent sugar units, a transport system to accumulate the sugar inside the cell, and enzymes to convert the sugar into one of the central intermediates of metabolism, such as glucose-6-phosphate. There are several central pathways for carbohydrate utilization, including the Embden-Meyerhof pathway of glycolysis and the pentose phosphate pathway, both of which are also present in eukaryotic cells (eukaryote). Some bacteria possess the Entner-Doudoroff pathway, which converts glucose primarily to pyruvate, as well as other pathways that accomplish the conversion of glucose into smaller compounds with fewer enzyme-catalyzed steps.
Sugar metabolism produces energy for the cell via two different processes, fermentation and respiration (cellular respiration). Fermentation is an anaerobic process that takes place in the absence of any external electron acceptor. The organic compound, such as a sugar or amino acid, is broken down into smaller organic molecules, which accept the electrons that had been released during the breakdown of the energy source. These catabolic reactions include a few steps that result in the direct formation of ATP. When glucose is broken down to lactic acid, as occurs in some Streptococcus and Lactobacillus species, as well as in muscle cells in higher eukaryotes, each molecule of glucose yields only two molecules of ATP, and considerable quantities of glucose must be degraded to provide sufficient energy for bacterial growth. Because organic molecules are only partially oxidized during fermentation, the growth of fermentative bacteria results in the production of large quantities of organic end products and a relatively small output of energy per glucose molecule consumed. Few bacteria produce only lactic acid, which is fairly toxic for bacteria and limits the growth of a colony. A variety of additional fermentation pathways are used by specific bacteria to break down glucose; the characteristic end products of these pathways assist in the identification of the bacteria. These end products are often less toxic than lactic acid or are formed with the harnessing of additional metabolic energy. For example, the products of mixed-acid fermentation in E. coli include lactic acid, succinic acid, acetic acid, formic acid, ethyl alcohol, carbon dioxide, and hydrogen gas (hydrogen). Enterobacter aerogenes produces most of the same set of fermentation products, as well as large amounts of 2,3-butylene glycol, which is nonacidic and permits more bacterial growth.
Considerably more energy is available to the cell from respiration, a process in which the electrons from molecules of sugar are transferred not to another organic molecule but to an inorganic molecule. The most familiar respiratory process (aerobic respiration) uses oxygen as the final electron acceptor. The sugar is completely broken down to carbon dioxide and water, yielding a maximum of 38 molecules of ATP per molecule of glucose. Electrons are transferred to oxygen using the electron transport chain, a system of enzymes and cofactors located in the cell membrane (membrane) and arranged so that the passage of electrons down the chain is coupled with the movement of protons (hydrogen ions (hydrogen ion)) across the membrane and out of the cell. Electron transport induces the movement of positively charged hydrogen ions to the outside of the cell and negatively charged ions to its interior. This ion gradient results in the acidification of the external medium and an energized plasma membrane with an electrical charge of 150 to 200 millivolts. The generation of ion gradients, including this protonmotive force (gradient of protons), is a common aspect of energy generation and storage in all living organisms. The gradient of protons is used directly by the cell for many processes, including the active transport of nutrients and the rotation of flagella (flagellum). The protons also can move from the exterior of the cell into the cytoplasm by passing through a membrane enzyme called the F1F0-proton-translocating ATPase, which couples this proton movement to ATP synthesis in a process identical to that which occurs in the mitochondria of eukaryotic cells (see cell: The mitochondrion and the chloroplast (cell); see also metabolism: The combustion of food materials (metabolism)).
Bacteria that are able to use respiration produce far more energy per sugar molecule than do fermentative cells, because the complete oxidation (oxidation–reduction reaction) (breakdown) of the energy source allows complete extraction of all of the energy available as shown by the substantially greater yield of ATP for respiring organisms than for fermenting bacteria. Respiring organisms achieve a greater yield of cell material using a given amount of nutrient; they also generate fewer toxic end products. The solubility of oxygen in water is limited, however, and the growth and survival of populations of aerobic bacteria are directly proportional to the available supply of oxygen. Continuous supplies of oxygen are available only to bacteria that come into contact with air, as occurs when bacteria are able to float on a surface that exposes them to air or when the medium in which the bacteria live is stirred vigorously.
Respiration can also occur under anaerobic conditions by processes called anaerobic respiration, in which the final electron acceptor is an inorganic molecule, such as nitrate (NO3−), nitrite (NO2−), sulfate (SO42−), or carbon dioxide (CO2). The energy yields available to the cell using these acceptors are lower than in respiration with oxygen—much lower in the case of sulfate and carbon dioxide—but they are still substantially higher than the energy yields available from fermentation. The ability of some bacteria to use inorganic molecules in anaerobic respiration can have environmental significance. E. coli can use oxygen, nitrate, or nitrite as an electron acceptor, and Pseudomonas stutzeri is of major global importance for its activity in denitrification (denitrifying bacteria), the conversion of nitrate to nitrite and dinitrogen gas (N2). Desulfovibrio and Desulfuromonas reduce sulfate and elemental sulfur (S), respectively, yielding sulfide (S2−), and the bacterium Acetobacterium woodii and methanogenic archaea, such as Methanobacterium thermautotrophicum, reduce carbon dioxide to acetate and methane, respectively. The Archaea typically use hydrogen as an electron donor with carbon dioxide as an electron acceptor to yield methane or with sulfate as an electron acceptor to yield sulfide.
Autotrophic metabolism
 Autotrophic bacteria synthesize all their cell constituents using carbon dioxide as the carbon source. The most common pathways for synthesizing organic compounds from carbon dioxide are the reductive pentose phosphate (Calvin) cycle, the reductive tricarboxylic acid cycle (tricarboxylic acid cycle), and the acetyl-CoA pathway (see photosynthesis: The process of photosynthesis: carbon fixation and reduction (photosynthesis)). The Calvin cycle, elucidated by American biochemist Melvin Calvin (Calvin, Melvin), is the most widely distributed of these pathways, operating in plants, algae, photosynthetic bacteria, and most aerobic lithoautotrophic bacteria. The key step in the Calvin cycle is the reaction of ribulose 1,5-bisphosphate with carbon dioxide, yielding two molecules of 3-phosphoglycerate, a precursor to glucose. This cycle is extremely expensive for the cell in terms of energy, such that the synthesis of one molecule of glyceraldehyde-3-phosphate requires the consumption of nine molecules of ATP and the oxidation of six molecules of the electron donor, the reduced form of nicotinamide adenine dinucleotide phosphate (NADPH). Autotrophic behaviour depends on the ability of the cell to carry out photosynthetic or aerobic respiratory metabolism, which are the only processes able to deliver sufficient energy to maintain carbon fixation.
Autotrophic bacteria synthesize all their cell constituents using carbon dioxide as the carbon source. The most common pathways for synthesizing organic compounds from carbon dioxide are the reductive pentose phosphate (Calvin) cycle, the reductive tricarboxylic acid cycle (tricarboxylic acid cycle), and the acetyl-CoA pathway (see photosynthesis: The process of photosynthesis: carbon fixation and reduction (photosynthesis)). The Calvin cycle, elucidated by American biochemist Melvin Calvin (Calvin, Melvin), is the most widely distributed of these pathways, operating in plants, algae, photosynthetic bacteria, and most aerobic lithoautotrophic bacteria. The key step in the Calvin cycle is the reaction of ribulose 1,5-bisphosphate with carbon dioxide, yielding two molecules of 3-phosphoglycerate, a precursor to glucose. This cycle is extremely expensive for the cell in terms of energy, such that the synthesis of one molecule of glyceraldehyde-3-phosphate requires the consumption of nine molecules of ATP and the oxidation of six molecules of the electron donor, the reduced form of nicotinamide adenine dinucleotide phosphate (NADPH). Autotrophic behaviour depends on the ability of the cell to carry out photosynthetic or aerobic respiratory metabolism, which are the only processes able to deliver sufficient energy to maintain carbon fixation.The aerobic nonphotosynthetic lithoautotrophs are those bacteria that not only use carbon dioxide as their sole carbon source but also generate energy from inorganic compounds (electron donors) with oxygen as an electron acceptor. These bacteria are taxonomically diverse and are usually defined by the electron donor that they use. For example, Nitrosomonas europaea oxidizes ammonia (NH4+) to nitrite, and Nitrobacter winogradsky oxidizes nitrite to nitrate. Thiobacillus oxidizes thiosulfate and elemental sulfur to sulfate, and T. ferrooxidans oxidizes ferrous ions to the ferric form. This diverse oxidizing ability allows T. ferrooxidans to tolerate high concentrations of many different ions, including iron, copper, cobalt, nickel, and zinc. All of these types of bacteria appear to be obligate lithotrophs and are unable to use organic compounds to a significant degree. Carbon monoxide (CO) is oxidized to carbon dioxide by Pseudomonas carboxydovorans, and hydrogen gas (H2) is oxidized by Alcaligenes eutrophus and, to a lesser degree, by many other bacteria.
Metabolic energy is made available from the oxidation of these electron donors in basically the same way as that used by respiring heterotrophs, which transfer electrons from an organic molecule to oxygen. As electrons are passed along the electron transport chain to oxygen, a proton gradient is generated across the cell membrane. This gradient is used to generate molecules of ATP. Other reactions present in lithoautotrophs are those used for the removal of electrons from the inorganic donor and for carbon dioxide fixation.
Phototrophic metabolism
Life on Earth is dependent on the conversion of solar energy to cellular energy by the process of photosynthesis. The general process of photosynthesis makes use of pigments called chlorophylls (chlorophyll) that absorb light energy from the Sun and release an electron with a higher energy level. This electron is passed through an electron transport chain, with the generation of energy by formation of a proton gradient and concomitant ATP (adenosine triphosphate) synthesis. The electron ultimately returns to the chlorophyll. This cyclic reaction path can fulfill the energy needs of the cell. For the cell to grow, however, the Calvin cycle of carbon dioxide fixation must be activated, and electrons must be transferred to the cofactor NADP to form NADPH, which is needed in large amounts for the operation of the cycle. Thus, phototrophic cell growth requires that a source of electrons be available to replace the electrons that are consumed during biosynthetic reactions.
Photosynthetic organisms are divided into two broad groups according to the nature of the source of these electrons. One group includes the higher plants, eukaryotic algae, and the cyanobacteria ( blue-green algae); these organisms contain the pigment chlorophyll a and use water as their electron source in reactions that generate oxygen. It is thought that predecessors of the cyanobacteria carried out the global production of oxygen on the originally anoxic (absence of oxygen) Earth some 1.5 billion years ago, which made possible the development of higher forms of life. Oxygen-evolving photosynthesis requires the action of two separate light-absorbing systems to raise the energy of the electrons from water to a level high enough for their transfer to NADP. Thus, two distinct photoreaction centres are present in these organisms, one for the oxygen-generating reaction and the other for the cyclic process for energy generation. In the cyanobacteria, both photoreaction centres contain chlorophyll a. Their photosynthetic apparatus also contains other light-absorbing pigments that serve as antennae to capture light energy and transfer it to the reaction centres. Cyanobacterial antennae include additional molecules of chlorophyll a, which transfer energy to the cyclic reaction centre, and phycobilisomes, which are protein pigments that absorb light of short, high-energy wavelengths (wavelength) and transmit this energy to the oxygen-evolving reaction centre. In almost all cyanobacteria, the photosynthetic apparatus is contained in an extensive intracellular system of flattened membranous sacs, called thylakoids, the outer surfaces of which are studded with regular arrays of phycobilisome granules. This arrangement, in which pigment aggregates exist on the thylakoid surfaces, is called a photosystem.
Other photosynthetic bacteria contain only a single type of reaction centre with a different pigment, called bacteriochlorophyll, which absorbs light of long, low-energy wavelengths. These organisms require an electron donor other than water and do not release oxygen. The green bacteria (Chlorobiaceae) and purple sulfur bacteria (Chromatiaceae) use elemental sulfur, sulfide, thiosulfate, or hydrogen gas as electron donor, whereas the purple nonsulfur bacteria use electrons from hydrogen or organic substrates. These bacteria require anaerobic conditions for photosynthetic activity. The photosystem in green bacteria is related to photosystem I of higher plants, whereas that in purple bacteria is related to photosystem II, which provides some indication of an evolutionary trail from bacteria to plants (see photosynthesis: The process of photosynthesis: the light reactions (photosynthesis)).
Biosynthetic pathways of bacteria
Many prokaryotes are able to convert any given carbon source into biosynthetic building blocks (anabolism)—e.g., amino acids (amino acid), purines (purine), pyrimidines (pyrimidine), lipids (lipid), sugars, and enzyme cofactors. The amount and activity of each enzyme in these biosynthetic pathways are carefully regulated so that the cell produces only as much of any compound as is needed at any time.
During the process of evolution, some bacteria have lost genes (gene) that encode certain biosynthetic reactions and are hence likely to require nutritional supplements. For example, Mycoplasma, whose DNA content is about one-quarter the size of that of E. coli, has many nutritional requirements and has even lost the ability to make a cell wall.
Classification (taxonomy) of bacteria
Classification by genetic divergence
Genetic approaches to the classification of bacteria are aimed at identifying a degree of relatedness between organisms to obtain a more fundamental measure of the time elapsed since two organisms diverged from a common ancestor.
The specific region of DNA that has proved to be the most informative for evolutionary relatedness is 16S rRNA, the gene that encodes the RNA component of the smaller subunit of the bacterial ribosome (16S refers to the rate of sedimentation, in Svedberg units, of the RNA molecule in a centrifugal field). The 16S rRNA gene is present in all bacteria, and a related form occurs in all cells. The 16S rRNA gene of E. coli is 1,542 nucleotides long, and some of its regions are double-stranded, while other regions are single-stranded. Single-stranded regions often form loops because there is a lack of complementary bases on the opposing strand. Since 16S rRNA makes very specific contacts with many different ribosomal proteins and with other parts of itself, the pace at which spontaneous random mutation can change the sequence of the bases in the rRNA is slow. Any change in sequence at one site must be compensated for by another change elsewhere within the rRNA or in a ribosomal protein, lest the ribosome fail to assemble properly or to function in protein synthesis and the cell die.
Analysis of the 16S rRNA sequences from many organisms has revealed that some portions of the molecule undergo rapid genetic changes, thereby distinguishing between different species within the same genus. Other positions change very slowly, allowing much broader taxonomic levels to be distinguished. The comparison of 16S rRNA sequences between organisms is quantitative and is based on a defined set of assumptions. The assumption that the rate at which base changes occur and are established within a species is constant is unlikely to be true. Changes in the Earth's environment are expected to alter the ecological niches or selective pressures that affect the rate of mutation and the rate at which various species are able to evolve.
The radical differences between archaea and Bacteria, which are evident in the composition of their lipids and cell walls and in the utilization of different metabolic pathways, enzymes, and enzyme cofactors, are also reflected in the rRNA sequences. The rRNAs of Bacteria and Archaea are as different from each other as they are from eukaryotic rRNA. This suggests that the bacterial and archaeal lines diverged from a common precursor somewhat before eukaryotic cells developed. This proposal also implies that the eukaryotic line is quite ancient and probably did not arise from any currently known bacteria. It had been previously believed that eukaryotic cells arose when some bacterial cells engulfed another type of bacterium. These bacteria might have formed a symbiotic relationship (symbiosis) in which the engulfed cell continued to survive but gradually lost its independence and took on the properties of an organelle. Although the original eukaryotic cell may or may not be derived from bacteria, it remains likely, if not certain, that eukaryotic organelles (e.g., mitochondria and chloroplasts (cell)) are descendants of bacteria that were acquired by eukaryotic cells in an example of symbiotic parasitism.
Early hypotheses about the origins of life suggested that the first cells obtained their energy from the breakdown of nutrients in a rich organic liquid environment proposed to have formed in the early oceans by the action of light and intense solar radiation on the early, anaerobic atmosphere. The process of photosynthesis might have evolved much later in response to the gradual depletion of those rich nutrient sources. On the other hand, rRNA sequence analysis places photosynthetic capability in almost all of the major bacterial divisions and shows that photosynthetic genera are closely related to nonphotosynthetic genera. Since photosynthesis is such a highly conserved, mechanistically complex process, it is unlikely that the ability to carry out photosynthesis could have evolved at different times in so many different organisms. Even more widely distributed among prokaryotes is lithotrophy (from the Greek word lithos, meaning “stone”), the ability to obtain energy by the transfer of electrons (electron) from hydrogen gas to inorganic acceptors. It has been proposed that the earliest forms of life on Earth used lithotrophic metabolism and that photosynthesis was a later addition to the early bacterial progenitors. The nonlithotrophic and nonphotosynthetic forms found today arose from the earliest forms of Bacteria, although they have lost their capacities for lithotrophy and photosynthesis.
The proposal that lithotrophy was widely distributed among bacterial organisms before photosynthesis developed suggests that the Archaea came from a different line of descent than Bacteria. The only photosynthetic archaeon, Halobacterium, has a completely different type of photosynthesis that does not use chlorophyll in large protein complexes to activate an electron, as in plants and bacteria. Rather, it uses a single protein, bacteriorhodopsin, in which light energy is absorbed by retinal, a form of vitamin A, to activate a proton ( hydrogen ion).
The analysis of rRNA sequences from bacteria that are closely related to one another has revealed several surprising relationships between these organisms. For example, mycoplasma, which appear to be different from other bacteria—in that they are very small, lack a cell wall, have a very small genome, and have sterols in their cell membranes—actually are related to some gram-positive clostridia on the basis of their nucleic acid sequences. This circumstance underscores the hazard of relying on phenotypic traits (phenotype) (observable characteristics such as the absence of a cell wall) for the assignment of evolutionary or genetic relationships. In fact, there are many groupings of bacteria that are not supported by RNA sequence analysis.
Classification by morphology, biochemistry, and other features
Although classification based on genetic divergence highlights the evolutionary relationships of bacteria, classification based on the morphological and biochemical features of bacteria remains the most practical way to identify these organisms. A definitive identification scheme for bacteria was first presented in 1984 in Bergey's Manual of Systematic Bacteriology. In this scheme, bacteria are classified on the basis of many characteristics. Cell shape, nature of multicell aggregates, motility, formation of spores (spore), and reaction to the Gram stain are important. These morphological features, including the shape and colour of bacterial colonies, are not always constant and can be influenced by environmental conditions. Important in the identification of a genus and species of bacteria are biochemical tests, including the determination of the kinds of nutrients a cell can use, the products of its metabolism, the response to specific chemicals, and the presence of particular characteristic enzymes. Other criteria used for the identification of some types of bacteria might be their antigenic composition, habitat, disease production, and requirement for specific nutrients. Some tests are based on the ultrastructure of the bacteria revealed under the electron microscope (microscope) by negative staining and preparation of thin sections.
Annotated classification
The following classification is based on the version of Bergey's Manual of Systematic Bacteriology that was published in the 1980s. This system uses the nature of the cell wall as the primary determinant for classification and differs substantially from earlier versions. The orders listed in this classification are not inclusive and are intended to be illustrative of only some of the different types of bacteria that are present.
Division Gracilicutes
Gram-negative cell wall. Non-endospore-forming. Includes photosynthetic and nonphotosynthetic types; can exhibit swimming or gliding motility; includes rods, cocci, and curved forms.
Class Scotobacteria
Nonphotosynthetic gram-negative bacteria.
Order Spirochaetales
Spiral cells that swim by flexion; found in water and in the bodies of vertebrates; genera include Borrelia, Treponema, and Leptospira, all parasites of humans and other animals.
Order Pseudomonadales
Rigid-walled cells of variable shape, in some species forming chains; photosynthetic pigment present in certain species; cells usually motile by means of a single flagellum. Species in soil and in fresh water and salt water. Examples of genera: vibrio ( cholera bacteria), Pseudomonas, Nitrosomonas, Thiobacillus.
Order Rickettsiales
Obligate intracellular parasites; generally short rods. Multiply by binary transverse fission; often cause disease in humans and are transmitted by arthropods (arthropod).
Class Anoxyphotobacteria
Gram-negative bacteria that carry out the type of photosynthesis that does not release oxygen. The major groupings within this class and some constituent genera are the purple sulfur bacteria, which use sulfide or elemental sulfur as electron donors (Chromatium); purple nonsulfur bacteria, which often use organic compounds as electron donors (Rhodobacter); green sulfur bacteria (Chlorobium); and filamentous green bacteria (Chloroflexus).
Class Oxyphotobacteria
Gram-negative bacteria that carry out oxygen-evolving photosynthesis. Includes the cyanobacteria (blue-green algae) and the order Prochlorales; gliding or nonmotile forms. Most cyanobacteria are photoautotrophs and can fix dinitrogen gas. Often form long cell filaments.
Division Firmicutes
Nonphotosynthetic gram-positive bacteria.
Class Firmibacteria
Nonbranching gram-positive bacteria. Includes rods and cocci forms. Some genera form endospores.
Class Thallobacteria
Gram-positive bacteria with branched or irregular walls. Some form spores on hyphae.
Order Actinomycetales (actinomycete)
Rigid-walled cells that may grow out in a branching system, resembling mold colonies. Includes Mycobacterium tuberculosis (Mycobacterium) ( tuberculosis bacterium), Streptomyces.
Division Tenericutes
Irregular pleiomorphic cell shapes due to the absence of a rigid cell wall. Lack peptidoglycans.
Class Mollicutes
Flexible-walled cells in the order Mycoplasmatales (mycoplasma); nonmotile, highly variable in shape at different life stages. Includes mycoplasma and forms once known as pleuropneumonia-like organisms (PPLO).
Division Mendosicutes
Cell wall, when present, lacks peptidoglycan. Rods or cocci.
Class Archaebacteria (archaea)
Possess cell walls and lipids with unusual compositions that differ from all other bacteria. Lipids usually are isoprenol derivatives linked to glycerol backbone through ether linkage. Ribosomes are different in protein composition and sensitivity to antibiotics than other bacterial ribosomes. Peptidoglycan, if present, does not contain muramic acid. Non-spore-forming. Most are anaerobic. None contain chlorophyll. Many are motile by flagella. Can stain gram-positive or gram-negative but have a different cell-wall appearance than do other bacteria.
Additional Reading
Comprehensive surveys are presented in John G. Holt (ed.), Bergey's Manual of Systematic Bacteriology, 4 vol. (1984–89), a reference and sourcebook accepted as standard throughout the world for classification of bacteria and related microorganisms; Thomas D. Brock, John M. Martinko, and Michael T. Madigan, Biology of Microorganisms, 11th ed. (2006), an advanced textbook covering all general characteristics of microorganisms, including morphology, physiology, biochemistry, ecological role, and classification; and Bernard D. Davis et al., Microbiology, 4th ed. (1990), a comprehensive textbook covering bacteriology, immunology, and virology as related to medical aspects of microbiology.Thomas D. Brock, The Emergence of Bacterial Genetics (1990), describes the historical development of bacterial genetics and molecular biology. Clive Edwards (ed.), Microbiology of Extreme Environments (1990), assembles a series of studies of microorganisms that live in extreme environments, with an emphasis on their applications in technology and ecology.
- litotes
- litre
- litter
- Little Alfold
- Little America
- Little Armenia
- Little Barrier Island
- Little Belt
- Little Belt Mountains
- Little Bighorn, Battle of the
- Little Brother Montgomery
- Little Brothers of Jesus and Little Sisters of Jesus
- Little Entente
- Little Falls
- Little Karoo
- Little League
- little magazine
- Little Octobrist
- Little Orphan Annie
- little owl
- Little Poland Uplands
- Little Richard
- Little Rock
- Little, Royal
- Little Saint Bernard Pass