chemoreception
physiology
Introduction
 process by which organisms respond to chemical stimuli in their environments that depends primarily on the senses of taste and smell. Chemoreception relies on chemicals (chemical reaction) that act as signals to regulate cell function, without the chemical necessarily being taken into the cell for metabolic purposes. While many chemicals, such as hormones (hormone) and neurotransmitters (neurotransmitter), occur within organisms and serve to regulate specific physiological activities, chemicals in the external environment are also perceived by and elicit responses from whole organisms. All animals (animal) and microorganisms such as bacteria exhibit this latter type of chemoreception, but the two commonly recognized chemosensory systems are the senses of taste, or gustation, and smell, or olfaction.
process by which organisms respond to chemical stimuli in their environments that depends primarily on the senses of taste and smell. Chemoreception relies on chemicals (chemical reaction) that act as signals to regulate cell function, without the chemical necessarily being taken into the cell for metabolic purposes. While many chemicals, such as hormones (hormone) and neurotransmitters (neurotransmitter), occur within organisms and serve to regulate specific physiological activities, chemicals in the external environment are also perceived by and elicit responses from whole organisms. All animals (animal) and microorganisms such as bacteria exhibit this latter type of chemoreception, but the two commonly recognized chemosensory systems are the senses of taste, or gustation, and smell, or olfaction.The following article discusses the role of taste and smell and the interaction of these two sensory systems in chemoreception. For basic information about the different senses used by animals, see sensory reception (senses). For information on specific senses, see also photoreception, thermoreception, and mechanoreception.
The senses of taste and smell
taste
 In terrestrial vertebrates (vertebrate), including humans, taste receptors (receptor) are confined to the oral cavity. They are most abundant on the tongue but also occur on the palate and epiglottis and in the upper part of the esophagus. The taste receptor cells, with which incoming chemicals interact to produce electrical signals, occur in groups of 50–150. Each of these groups forms a taste bud. On the tongue, taste buds are grouped together into taste papillae. On average, the human tongue has 2,000–8,000 taste buds, implying that there are hundreds of thousands of receptor cells. However, the number of taste buds varies widely; some humans have only 500, whereas others have as many as 20,000. Healthy humans may have anywhere from three to several thousand taste buds per square centimetre on the tip of the tongue, and this variability contributes to differences in the taste sensations experienced by different people.
In terrestrial vertebrates (vertebrate), including humans, taste receptors (receptor) are confined to the oral cavity. They are most abundant on the tongue but also occur on the palate and epiglottis and in the upper part of the esophagus. The taste receptor cells, with which incoming chemicals interact to produce electrical signals, occur in groups of 50–150. Each of these groups forms a taste bud. On the tongue, taste buds are grouped together into taste papillae. On average, the human tongue has 2,000–8,000 taste buds, implying that there are hundreds of thousands of receptor cells. However, the number of taste buds varies widely; some humans have only 500, whereas others have as many as 20,000. Healthy humans may have anywhere from three to several thousand taste buds per square centimetre on the tip of the tongue, and this variability contributes to differences in the taste sensations experienced by different people.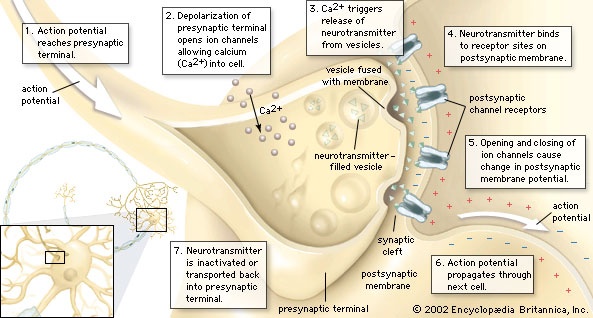 The taste buds are embedded in the epithelium of the tongue and make contact with the outside environment through a taste pore. Slender processes (microvilli) extend from the outer ends of the receptor cells through the taste pore, where the processes are covered by the mucus that lines the oral cavity. At their inner ends the taste receptor cells synapse, or connect, with afferent sensory neurons (neuron), nerve cells that conduct information to the brain. Each receptor cell synapses with several afferent sensory neurons, and each afferent neuron branches to several taste papillae, where each branch makes contact with many receptor cells. Unlike the olfactory system, in which input to the brain involves a single nerve, the afferent sensory neurons occur in three different nerves running to the brain—the facial nerve, the glossopharyngeal nerve, and the vagus nerve. Taste receptor cells of vertebrates are continually renewed throughout the life of the organism.
The taste buds are embedded in the epithelium of the tongue and make contact with the outside environment through a taste pore. Slender processes (microvilli) extend from the outer ends of the receptor cells through the taste pore, where the processes are covered by the mucus that lines the oral cavity. At their inner ends the taste receptor cells synapse, or connect, with afferent sensory neurons (neuron), nerve cells that conduct information to the brain. Each receptor cell synapses with several afferent sensory neurons, and each afferent neuron branches to several taste papillae, where each branch makes contact with many receptor cells. Unlike the olfactory system, in which input to the brain involves a single nerve, the afferent sensory neurons occur in three different nerves running to the brain—the facial nerve, the glossopharyngeal nerve, and the vagus nerve. Taste receptor cells of vertebrates are continually renewed throughout the life of the organism.The taste receptor system of terrestrial vertebrates is concerned with the detection of chemicals that are taken into the oral cavity and are present at relatively high concentrations. In humans, five different classes, or modalities, of taste are usually recognized: sweet, salt, sour, bitter, and umami. But this is an anthropocentric view of a system that has evolved to give animals information about the nutrient content and the potential dangers of the foods they eat. The major nutrient requirements of all animals are carbohydrates (carbohydrate), which act principally as a source of energy. Many lipids (lipid) can be synthesized from carbohydrates, and animals use proteins (protein) derived from carbohydrates to assemble their own body proteins. In general, animals are unable to taste proteins, but they do taste amino acids (amino acid) (from which proteins are made). Some of the amino acids taste sweet to humans, whereas others taste sour, and umami taste, which is meatlike, is a response to glutamic acid and its derivatives, such as monosodium glutamate (MSG). Sweet taste comes mainly from sugars (sugar) (carbohydrates), and bitter taste derives from potentially harmful chemicals present in food, especially plants (plant), which produce thousands of chemicals that offer the plants some protection from herbivores (herbivore). The constituents of inorganic salts (salt), such as sodium chloride, potassium chloride, and calcium chloride, are essential nutrients, but the quantities required to fulfill animal nutrient requirements are relatively small. It is possible that the salt taste reflects an animal's need to avoid ingesting too much salt, which would increase the osmotic pressure in body tissues, producing adverse effects on cell metabolism. Animals experiencing a salt deficit actively seek out and eat sodium chloride, but the sensory basis for this salt appetite is not understood. Minor essential nutrients, such as sterols and vitamins (vitamin), are not known to be tasted by animals. They are probably of such widespread occurrence that an animal's normal food contains sufficient quantities, which is true for inorganic salts. However, associative learning may also have an important role in ensuring that appropriate levels of these compounds are obtained (see below Behaviour and chemoreception: Associative learning (chemoreception)). Except for bitter-tasting substances, the chemicals that stimulate taste receptors are generally water soluble.
Humans do not make further distinctions within the five modalities. For example, different sugars may have different degrees of sweetness, but they do not have distinct tastes. Similarly, bitter-tasting substances, such as quinine or caffeine, taste bitter but do not induce separate tastes, despite great differences in their molecular structures. However, the umami receptor does give the ability to distinguish between naturally occurring amino acids and is sensitive to MSG. Natural foods contain many different chemicals; for example, the taste of an apple may stimulate all the different types of receptors to different degrees.
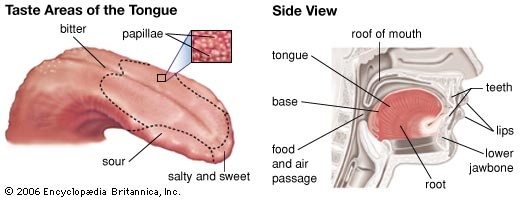 Although there is evidence that all taste buds exhibit sensitivity to all taste sensations, some areas of the tongue are sensitive to specific tastes. For example, in humans and some other mammals (mammal), taste papillae with receptor cells most sensitive to sweet taste are located at the front of the tongue, receptors preferentially tasting salt and sour are at the sides of the tongue, and receptors preferentially tasting bitter substances are at the back of the tongue. The taste receptor cells of other animals can often be characterized in similar ways to those of humans, because all animals have the same basic needs in selecting food. In addition, some organisms have other types of receptors that permit them to distinguish between classes of chemicals not directly related to diet and that enable them to make further distinctions within the modalities.
Although there is evidence that all taste buds exhibit sensitivity to all taste sensations, some areas of the tongue are sensitive to specific tastes. For example, in humans and some other mammals (mammal), taste papillae with receptor cells most sensitive to sweet taste are located at the front of the tongue, receptors preferentially tasting salt and sour are at the sides of the tongue, and receptors preferentially tasting bitter substances are at the back of the tongue. The taste receptor cells of other animals can often be characterized in similar ways to those of humans, because all animals have the same basic needs in selecting food. In addition, some organisms have other types of receptors that permit them to distinguish between classes of chemicals not directly related to diet and that enable them to make further distinctions within the modalities. smell
The olfactory system is concerned with the detection of airborne or waterborne (in aquatic animals) chemicals that may be present in very low concentrations. olfactory receptor cells are present in very large numbers (millions), forming an olfactory epithelium within the nasal cavity. Each receptor cell has a single external process that extends to the surface of the epithelium and gives rise to a number of long, slender extensions called cilia (cilium). The cilia are covered by the mucus of the nasal cavity. Unlike taste receptor cells, olfactory receptor cells have axons (axon) that connect directly to the brain. Olfactory receptor cells are continually replaced, with new cells developing from basal cells in the olfactory epithelium. In humans the receptor cells are replaced about every 60 days.
The relative size of the olfactory epithelium reflects the importance of olfaction in the lives of different animals. In some dogs (dog) the olfactory epithelium has an area of about 170 cm2, with a total of about 1 billion olfactory receptor cells; in oxen (ox) the olfactory epithelial area is only about 1–4 cm2, and the number of cells is less than 30 million. By comparison the human olfactory epithelium covers about 5–10 cm2 and has about 10–40 million olfactory receptor cells.

 Another major difference between the olfactory system and the taste system is that the axons of olfactory receptor cells extend directly into a highly organized olfactory bulb, where olfactory information is processed. Within the olfactory bulb are discrete spheres of nerve tissue called glomeruli. They are formed from the branching ends of axons of receptor cells and from the outer (dendritic) branches of interneurons, known in vertebrates as mitral cells, that pass information to other parts of the brain. Tufted cells, which are similar to but smaller than mitral cells, and periglomerular cells, another type of interneuron cell, also contribute to the formation of glomeruli. The axons of all the receptor cells that exhibit a response to a specific chemical or a range of chemicals with similar structures converge on a single glomerulus, where they connect via synapses with the interneurons. In this way, information from large numbers of receptor cells with similar properties is brought together. Thus, even if only a few receptors are stimulated because of very low concentrations of the stimulating chemical, the effects of signals from these cells are maximized. In mice (mouse) there are about 1,800 glomeruli on each side of the brain, in rabbits (rabbit) there are about 2,000, and in dogs there are as many as 5,000. Since there are millions of olfactory receptor cells, the degree of convergence of axons, and therefore of information about a particular odour, is enormous. For example, in a rabbit, axons from about 25,000 receptor cells converge on each glomerulus.
Another major difference between the olfactory system and the taste system is that the axons of olfactory receptor cells extend directly into a highly organized olfactory bulb, where olfactory information is processed. Within the olfactory bulb are discrete spheres of nerve tissue called glomeruli. They are formed from the branching ends of axons of receptor cells and from the outer (dendritic) branches of interneurons, known in vertebrates as mitral cells, that pass information to other parts of the brain. Tufted cells, which are similar to but smaller than mitral cells, and periglomerular cells, another type of interneuron cell, also contribute to the formation of glomeruli. The axons of all the receptor cells that exhibit a response to a specific chemical or a range of chemicals with similar structures converge on a single glomerulus, where they connect via synapses with the interneurons. In this way, information from large numbers of receptor cells with similar properties is brought together. Thus, even if only a few receptors are stimulated because of very low concentrations of the stimulating chemical, the effects of signals from these cells are maximized. In mice (mouse) there are about 1,800 glomeruli on each side of the brain, in rabbits (rabbit) there are about 2,000, and in dogs there are as many as 5,000. Since there are millions of olfactory receptor cells, the degree of convergence of axons, and therefore of information about a particular odour, is enormous. For example, in a rabbit, axons from about 25,000 receptor cells converge on each glomerulus.The olfactory system enables an animal (animal communication) to perceive chemicals originating outside itself that are important in the animal's behaviour and ecology. These signals do not fall into such relatively clear categories as the taste receptor system, and most organisms have the ability to distinguish between hundreds or even thousands of odours, including some odours that have very similar chemical structures. An example of the human ability to discriminate between odours is the difference in smell between caraway seed and spearmint. Yet the chemicals producing these odours, the s- and r-forms of carvone, are stereoisomers (having the same three-dimensional chemical structure, but one being a mirror image of the other). This ability to distinguish between different compounds depends on the possession of olfactory receptor cells with specific, limited ranges of sensitivity. Many of the compounds that stimulate the olfactory system of terrestrial animals are not water soluble.
 In terrestrial vertebrates the olfactory epithelium is in the nasal cavity. Because air passes through this cavity to the lungs (lung), the epithelium is continually bathed with a fresh supply of air as the animal breathes. The airflow can be enhanced so that the volume of air sampled is increased by sniffing, a technique commonly used by cats (cat, domestic), dogs, and many other animals. When bird dogs are searching for a scent on the ground, they may sniff very rapidly, perhaps creating turbulence of the air in the nasal cavity and enhancing the likelihood that odour molecules will reach the olfactory epithelium. When these dogs run into the wind with their heads held high, attempting to pick up the scent of prey, a continuous flow of inhalant air is maintained through the nostrils and thus over the olfactory epithelium.
In terrestrial vertebrates the olfactory epithelium is in the nasal cavity. Because air passes through this cavity to the lungs (lung), the epithelium is continually bathed with a fresh supply of air as the animal breathes. The airflow can be enhanced so that the volume of air sampled is increased by sniffing, a technique commonly used by cats (cat, domestic), dogs, and many other animals. When bird dogs are searching for a scent on the ground, they may sniff very rapidly, perhaps creating turbulence of the air in the nasal cavity and enhancing the likelihood that odour molecules will reach the olfactory epithelium. When these dogs run into the wind with their heads held high, attempting to pick up the scent of prey, a continuous flow of inhalant air is maintained through the nostrils and thus over the olfactory epithelium.Interaction between taste and smell
In humans and other terrestrial vertebrates, odours can reach the olfactory epithelium via the external nostrils of the nose and the internal nares, which connect the nasal cavity and the back of the oral cavity. The latter pathway becomes important when eating, and, as a result, there is considerable confusion in the use of the term taste, because odours from the food enter the nasal cavity at the same time as the taste buds are stimulated by food. The importance of odour in the common concept of taste becomes obvious when a person has a cold and can no longer “taste” food. In this case, although the taste receptor system is completely unimpaired, access to the olfactory epithelium is blocked. It is clear that the taste and smell systems are distinct in both their anatomy and their neural processing of inputs. The term flavour is an alternative to taste in the context of food, with flavour referring to the overall perception that results from both taste and smell. Use of this term avoids the confusion otherwise produced by using taste to refer specifically to the sensations produced by stimulation of taste receptors, as well as to the combined sensations of taste and smell. Although the same arguments apply to other terrestrial vertebrates, there is little knowledge of the extent to which flavour, as opposed to taste, is important in other organisms.
Cellular mechanisms in chemoreception
To produce a behavioral response in an organism, a chemical must produce a signal in the organism's nervous system. This entails processes that are initiated at the taste or smell receptor cells (cell). First, the molecule must be captured in and traverse a layer of mucus, in which the endings of the receptor cell are bathed; these are known as perireceptor events. Second, the molecule must interact with the surface of the receptor cell in a specific way to produce reactions within the cell. These reactions lead to a change in cellular electrical charge, which generates a nerve impulse. Transformation of an external stimulus into a cellular response is known as signal transduction.
The electrical signal produced by a particular nerve cell is the same regardless of the nature of the stimulus. If chemicals are to be distinguished from one another, they must stimulate separate cells. Thus, different cells are responsible for the reception of sweet, salt, sour, and bitter tastes and for distinguishing the different odours detected by the olfactory system.
Perireceptor events
Water-soluble compounds, such as sugars (sugar) and amino acids (amino acid), can move freely in the mucus covering the taste and olfactory receptor cells. However, most bitter-tasting and many volatile compounds are not water soluble and must be made soluble if they are to reach the receptors. This is achieved by binding them to soluble proteins (protein), which can move freely through the mucus. Such proteins have been isolated both from saliva and from the mucus in the nasal epithelium, although the precise role of soluble proteins in transporting chemicals to receptor cells has yet to be clearly demonstrated in mammals (mammal).
In insects (insect), taste and olfactory neurons (neuron) are contained within cuticular structures, but the sensitive nerve endings are bathed in a fluid called sensillar lymph that is analogous to the mucus of vertebrates (vertebrate). In the olfactory system this fluid contacts odour-binding receptors that presumably function in the same way as those of vertebrates but that are produced by different families of genes (gene). Three families of these receptor proteins have been identified. One family, consisting of pheromone-binding proteins, is restricted to receptors known to be sensitive to pheromones. The remaining two families contain general odorant receptors that respond to other odours (not pheromones). These proteins, to differing extents, govern which chemicals reach the membrane (cell) of the receptor cell and can be regarded as filters. Differences in their binding capacity could account for some of the differences in sensitivity of different receptor cells.
It is important that taste and odour molecules be removed from the immediate environment of the receptor cell; otherwise the cell, and thus the animal, continues to respond to something that is no longer relevant. Removal of the unwanted molecules is thought to be achieved, at least in part, by odorant-degrading enzymes (enzyme) that are also present in the mucus or other fluid surrounding the sensitive endings of the receptor cells.
Signal transduction
Information is conveyed along neurons (neuron) by electrical signals called action potentials (action potential) that are initiated by electrical changes in receptor cells. In the case of chemoreceptors, these electrical changes are induced by chemicals. The initial changes are called receptor potentials, and they are produced by the movement of positively charged ions (ion) (e.g., sodium ions) into the cell through openings in the cell membrane called ion channels. Thus, in order to stimulate a receptor cell, a chemical must cause particular ion channels to be opened. This is achieved in various ways, but it most commonly involves specific proteins called receptors (receptor) that are embedded in the cell membrane.
Within the cell membrane, receptor proteins are oriented in such a way that one end projects outside the cell and the other end projects inside the cell. This makes it possible for a chemical outside the cell, such as a molecule of an odorant or a tastant compound, to communicate with and produce changes in the cellular machinery without entering the cell. The outer and inner ends of receptor proteins involved in taste and smell are connected by a chain of amino acids. Because the chain loops seven times through the thickness of the cell membrane, it is said to have seven transmembrane domains. The sequence of amino acids forming these proteins is critically important. It is thought that stimulation occurs when a molecule with a particular shape fits into a corresponding “pocket” in the receptor molecule, rather as a key fits into a lock. A change in a single amino acid can change the form of the pocket, thus altering the chemicals that fit into the pocket. For example, one olfactory receptor protein in rats (rat) produces a greater response in the receptor cell when it interacts with an alcohol called octanol (eight carbon atoms) rather than with an alcohol known as heptanol (seven carbon atoms). Changing one amino acid from valine to isoleucine in the fifth transmembrane domain, which is thought to contribute to the shape of the pocket, alters the receptor protein in such a way that heptanol, instead of octanol, produces the greatest effect. In mice (mouse) the equivalent receptor is normally in this form, producing a greater response to heptanol than to octanol. This illustrates the importance of amino acid molecules in determining the specificity of receptor cells.
When a receptor protein binds with an appropriate chemical (known as a ligand), the protein undergoes a conformational change, which in turn leads to a sequence of chemical events within the cell involving molecules called second messengers (second messenger). Second-messenger signaling makes it possible for a single odour molecule, binding with a single receptor protein, to effect changes in the degree of opening of a large number of ion channels. This produces a large enough change in the electrical potential across the cell membrane to lead to the production of action potentials that convey information to the animal's brain.
In mammals (mammal), five families of genes (gene) encoding chemoreceptor proteins have been identified. (Genes are considered to belong to the same family if they produce proteins in which high proportions of the amino acids are arranged in similar sequences.) Two families of genes are associated with taste, one with smell, and two with the vomeronasal system (see below Chemoreception in different organisms: Terrestrial vertebrates (chemoreception)). There are about 1,000 genes in the olfactory gene family, the largest known family of genes. Since each gene produces a different odour receptor protein, this contributes to the ability of animals to smell many different compounds. Animals not only can smell many compounds but can also distinguish between them. This requires that different compounds stimulate different receptor cells. Consistent with this, evidence indicates that only one olfactory gene is active in any one olfactory receptor cell. As a consequence, each receptor cell possesses only one type of receptor protein, though it has many thousands of the particular type on the membrane of the exposed cilia of the cell. Since each cell expresses only one type of receptor protein, there must be large numbers of cells expressing each type of receptor protein to increase the likelihood that a particular odour molecule will reach a cell with the appropriate receptor protein. Once the molecule reaches the matching receptor, the cell can respond.
A quite different family of genes (gene) produces the receptor proteins associated with bitter taste, but this family is much smaller than the olfactory gene family, containing only about 80 different genes. Given the very wide range of chemical structures that produce bitter taste, it is logical that there should be a number of different receptor proteins. However, unlike with the olfactory response, animals do not distinguish different bitter compounds. This is because each of the receptor cells stimulated by these compounds produces many different kinds of receptor proteins. Thus, the same cell responds to many different compounds. This does not mean necessarily that all the genes are expressed by all the bitter-sensitive cells. It is probable that the perception of sugars, giving sweet taste, and amino acids, giving umami taste, also depend on protein receptors in the receptor cell membranes.
The mechanism by which inorganic salts are perceived is probably quite different. Because changes in electrical properties (electric field) of cell membranes depend on ionic movement, cells will be affected by ion concentrations in the medium that bathes them. It is very likely that when humans and other animals ingest common salt ( sodium chloride), sodium enters the receptor cells directly through sodium channels in the cell membrane. This has the effect of altering the internal ionic concentration and initiating an electrical signal. Responses to other salts are probably mediated in the same way, and responses to acids (acid) (sour) may be similarly effected by the movement of hydrogen ions (hydrogen ion). Acids might also produce their effects by opening ion channels that are sensitive to pH.
The gene family that governs the production of olfactory receptors (olfactory receptor) is common to all vertebrates. Yet it is well known that mammals differ in the extent to which their behaviour is affected by odours. This is a reflection of the different numbers of olfactory receptor genes that are active. In mice, which have a highly developed sense of smell, most of the approximately 1,000 olfactory genes are expressed (that is, they produce receptor proteins). But in Old World monkeys (monkey) and in the great apes (ape), gorillas (gorilla), chimpanzees (chimpanzee), and humans, as many as 70 percent of the olfactory receptor genes, though still identifiable, are nonfunctional pseudogenes. Evidence indicates that the pool of pseudogenes in humans is increasing, suggesting that, at some time in the future, the human sense of smell will be reduced even further than it is today. All the olfactory genes of dolphins (dolphin) are nonfunctional.
Animal responses to chemicals are greatly affected by chemical concentration. The more sugar present in coffee, the sweeter it tastes, and a smell may be barely perceptible or overpowering. These effects, which are very general and experimentally demonstrated in many animals, arise from the presence of large numbers of molecules at high concentration. As concentration increases, more cells are stimulated and more receptor molecules in a taste or olfactory cell are filled at one time. The result is that more action potentials (nerve impulses) are generated by more receptor cells, and the signal reaching the brain is strengthened.
It is a common occurrence that, when entering a room, a person may notice a pleasant or unpleasant smell, but within a very short time he can no longer smell it, even though the source of the smell remains. The effect is due to a waning of the response of the receptor cells and is called sensory adaptation. The cells may adapt completely within a few seconds but become responsive again following an interval without stimulation. Adaptation of taste and olfactory cells occurs in all animals but not in receptor cells of the vomeronasal organ (Jacobson organ (Jacobson's organ)).
Processing olfactory information
Although each olfactory receptor cell has only one type of receptor protein, this does not mean that each cell responds to only one chemical. Presumably the receptor site formed by the protein interacts with some specific molecular form, and any chemical that possesses this form in some part of its molecule will stimulate the cell. For example, the alcohol nonanol contains nine carbon atoms linked together linearly. It might be expected that other compounds with a similar structure would interact with the same receptor protein, and this is the case with nonanoic acid in at least some olfactory receptors of the mouse. Comparable molecules having only eight carbon atoms stimulate the same cell but require higher concentrations to activate the receptor than do molecules with nine carbon atoms, and molecules with five carbon atoms do not stimulate the receptor at all.
Each chemical interacts with more than one type of receptor protein, and, since each cell only expresses one protein, the chemical can stimulate more than one cell type. It is thought that different receptor proteins “recognize” different parts of the molecule. For example, some receptors interact with compounds exhibiting the characteristic features of an alcohol, whereas others interact with compounds having characteristic features of acids. As a result, each chemical stimulates an array of cells with different receptors, and, although each cell may be stimulated by several different compounds, the array stimulated by each compound is unique. Since each receptor cell is connected to a single glomerulus, which receives the inputs from all the receptor cells expressing a particular receptor protein, the unique set of information is conveyed into the brain, providing the basis for odour recognition. With up to 1,000 different types of receptor proteins, the number of possible combinations is enormous. This broad range of combinations provides animals with their extraordinary capacity to distinguish between thousands of odours.
Chemoreception in different organisms
Single-celled organisms
Many microorganisms are known to remain in favourable chemical environments and to disperse away from unfavourable environments. This implies that microorganisms have a chemical sense, but, because they are so small, they are unable to detect chemical gradients by simultaneous comparison of the chemical concentration at two parts of the body. Instead, microorganisms exhibit differential responses to temporal differences in concentration, implying that they have the capacity to “remember” whether the concentration previously experienced was higher or lower than the current concentration. Movement in these organisms consists of periods of movement in a straight line interrupted at intervals by a turn, or “tumble.” The organisms swim smoothly up the concentration gradient of an attractant and begin to accumulate in areas of high concentration of the attractant. Accumulation is reinforced by the organisms' own secretion of attractant chemicals. Organisms that leave the aggregation tumble, and the direction of the turn and of the new path relative to the original appear to be random. The rate of tumbling varies, with organisms tumbling most in the absence of attractants and in the presence of repellents. Organisms that tumble away from an aggregation typically swim in a straight line back to the attractant. The bacterium (bacteria) Escherichia coli accumulates in high concentrations of sugars (sugar) and some amino acids (amino acid). This is also true of the ciliate protozoan Paramecium, which accumulates in areas with high concentrations of folate or biotin—compounds that are released by bacteria, the food of these animals. However, Paramecium disperses when it encounters quinine or potassium hydroxide.
As in multicellular organisms, perception of chemicals often involves the possession of receptor proteins in the cell membrane (cell) that activate second-messenger (second messenger) systems within the cell. However, unlike with multicellular organisms, the second messengers (second messenger) of single-celled organisms cause changes in the effector mechanisms of the cell, such as the flagellum or cilium, that modify the cell's movement. This causes the organism to move appropriately, relative to the stimulus. The receptor proteins of the yeast Saccharomyces and the slime mold Dictyostelium both have seven transmembrane domains, similar to the olfactory receptors of higher organisms, although belonging to different gene families. However, in the bacterium E. coli the receptor proteins have only two transmembrane domains, perhaps reflecting the fact that bacteria, as prokaryotes (prokaryote) (lacking distinct nuclei), predate the evolution of eukaryotes (having membrane-bound nuclei).
The number of different types of receptor proteins is limited in single-celled organisms compared with multicellular organisms. This appears to be the result of limited space available on the surface of a single cell. In E. coli there are five types of receptor proteins involved in positive responses. One receptor responds to serine, an amino acid (this receptor is also sensitive to temperature and pH); a second receptor responds to aspartate and ribose, an amino acid and a sugar, respectively; a third receptor responds to galactose and maltose, both sugars; a fourth receptor responds to dipeptides; and a fifth receptor responds to oxygen and changes in reduction-oxidation potential in the cell. Metallic ions, organic acids, inorganic acids, and glycerol produce negative responses, but it is not clear whether these molecules act via receptors or via an alternative mechanism. Paramecium has membrane receptor proteins that respond to favourable compounds such as biotin and to aversive compounds such as quinine. Several hundred of each receptor type are present on the cell surface, and they may be differentially distributed; for example, Paramecium has more quinine receptors at its front end than at its back end. In E. coli a difference in concentration producing a change in the occupancy of only a single receptor site is sufficient to produce a change in behaviour.
In addition to receptor-mediated responses, environmental chemicals may act on intracellular processes by entering the cell. In bacteria, for example, sugars and some other compounds act intracellularly, and, in Paramecium, ammonium ions enter the cell as ammonia, changing the pH of the cytoplasm and affecting the membrane potential. Inside the cell these effects are integrated with effects produced via cell membrane receptors. Therefore, the overall effect in Paramecium is to change the cell membrane potential, with favourable stimuli causing slight hyperpolarization (the potential difference across the cell membrane is increased), which increases the frequency of ciliary beating and reduces the frequency with which the organism makes turns, and aversive substances producing slight depolarization (a reduction in the potential difference across the cell membrane). In flagellates (flagellate), changes in flagellar movement do not depend on general membrane effects. In species with a single flagellum, changes in direction are induced by reversals in the direction of flagellar rotation from counterclockwise to clockwise. The several flagella of E. coli normally rotate counterclockwise, and, when the flagella all have the same rotation, they form a bundle that drives the organism in a straight path. However, when one or more flagella rotate in the opposite direction, the unity of the bundle is destroyed, and the bacterium tumbles.
sperm of all animals are faced with the problem of locating an egg, whether the eggs are free in the environment, such as those released from sea urchins (sea urchin) and toads (toad), or are contained within the female ducts, such as the eggs of humans. In toads and humans, sperm have been shown to make directed movements toward eggs, and there is evidence that they move up the concentration gradient of a small protein released by the egg. In sea urchin sperm, comparable small proteins are detected by receptors in the cell membrane, and this is probably true of all species.
Specialized chemosensory structures
Many invertebrates (invertebrate) have chemoreceptor cells contained in discrete structures called sensilla that are located on the outside of the body. Each sensillum consists of one or a small number of receptor cells together with accessory cells derived from the epidermis. These accessory cells produce a fluid (analogous to vertebrate mucus) that protects the nerve endings from desiccation and provides the constant ionic environment necessary for nerve cells to function properly. In some animals the sensillum and accessory cells form a physical structure around the receptor cells. Chemicals in the environment reach the receptor cells through one or more pores in this protective covering. In some invertebrates sensilla are found all over the body, including on the legs, cerci, and wing margins. In polychaetes (polychaete) the sensilla are often borne on tentacles.
The number of chemoreceptor cells in nematodes (nematode) is very limited. Caenorhabditis elegans, a small soil-inhabiting species, has only 34 chemosensory cells arranged in eight sensilla near the head. This organism also has four sensory cells in the tail, although it is not known whether these cells function as chemoreceptors.
Despite the small number of chemosensory cells, nematodes are capable of responding to many different chemicals, including water-soluble and lipophilic chemicals. As in all other animals, much of their chemoreceptor capability depends on having appropriate receptor proteins in the receptor cells. In C. elegans there may be more than 700 genes controlling receptor protein production. However, because the number of receptor cells is limited, some of the cells must express more than one type of receptor protein. The nature of the connections made by the receptor cells with other components of the nervous system then determines the behaviour that a particular chemical will elicit. By experimentally moving a particular receptor protein from one receptor cell to another, an animal's response can be reversed from being attracted to a particular chemical to being repelled by the chemical.
Animals with separate taste and olfactory systems
Arthropods (arthropod)
Arthropods (arthropod) (e.g., crabs (crab), insects (insect), spiders (spider)) are unique among invertebrates in that they have clearly separate senses of taste and olfaction that are comparable to those of vertebrates. Similar to nematodes, arthropods have a continuous layer of cuticle covering the outside of the body that separates the epidermis from the environment. For chemoreception to occur, the chemosensory cells must be exposed to the environment, and this is achieved through small pores in the cuticle. Most commonly the pores are in hairlike extensions of the cuticle that enclose the outer ends (dendrites) of the receptor cells. Two basic types of structure are recognized: those with a single pore, about 0.15–2 μm in diameter, at the tip of the hair (uniporous) and those with many small pores, about 10 nm in diameter, scattered over the surface of the hair (multiporous). These types are associated with the senses of taste and smell, respectively.
Taste receptor sensilla of arthropods occur mainly on feeding appendages associated with but located outside the mouth. They often occur in groups. In addition, many arthropods have taste receptors on the legs, especially on the ventral surfaces of the tarsi (feet), where they come into contact with whatever the animal is walking on. In some species similar receptors are scattered over the surface of the body and may also be present on egg-laying apparatus.
It is common for four taste receptor cells to be associated with each hair; however, unlike the taste receptor cells of vertebrates, these cells have axons (axon) that extend directly, without any synapses (synapse), to the central nervous system. Arthropods are segmented animals and have a nerve ganglion in each segment, although the ganglia often become fused together. The axons of taste receptor cells extend only as far as the ganglion of the segment on which they occur, and there is no “taste centre,” to which all information concerning taste is conveyed, in the central nervous system.
The taste receptors of insects (insect), which are the most studied of the arthropods, respond mainly to food-related chemicals, and the sensitivities of the cells vary depending on the nature of the insect's food. In most plant-feeding species the four cells within a hair may respond most actively to sugars, amino acids, inorganic salts, and a range of compounds produced by plants that generally inhibit feeding (feeding behaviour). These four categories roughly correspond to the human sweet, sour, salt, and bitter modalities. Bloodsucking insects have receptor cells that are sensitive to adenine nucleotides (nucleotide) (adenosine diphosphate 【ADP】 and adenosine triphosphate 【ATP】), and some insects, such as mosquitoes (mosquito) and blowflies (blow fly), have cells that respond to very low salt concentrations. Apart from bitter-sensitive cells, these cells usually respond to only limited ranges of compounds, even within the class of chemicals to which they are sensitive. For example, a cell may respond to glucose and sucrose but not to fructose, and amino acid-sensitive cells respond to only some amino acids. However, different cells may be sensitive to different groups of these compounds, providing many insects with the capacity to distinguish between suites of amino acids or, sometimes, different sugars. This presumably reflects the occurrence of different receptor proteins in the cell membranes, but little is known about this in insects. In the black blowfly there is evidence that the receptor cells responding to sugars have two receptor proteins, one that recognizes glucose and sucrose and another that recognizes fructose. Since both types of sugar stimulate receptors on the same cell, the fly is unable to distinguish them; a similar arrangement probably occurs in humans. If the receptor proteins were on different cells, the insect would be able to distinguish between the two types of sugar, and this is the case in some insect species.
Some plant-feeding insects that feed on only one or a few closely related plant species have taste receptor cells specialized to perceive chemicals specific to the host. For example, plants in the cabbage family (crucifers) are characterized by a class of compounds called glucosinolates, and some crucifer-feeding insects have cells that respond only to glucosinolates, often exhibiting greatest sensitivity to the specific glucosinolates that occur in their normal hosts. Adult butterflies (butterfly) and adults of some plant-feeding flies (fly) may have similar receptor cells on their tarsi, facilitating the recognition of host plants on which to lay eggs. Thus, this response is not concerned with indicating the nutritional status of a plant; rather, it provides the insect with a stimulus indicating that the plant is taxonomically appropriate. Some insects also have receptor cells in their taste hairs that recognize pheromones (pheromone) on the surface of other members of the species. Because perception of these chemicals may have nothing to do with feeding (in relation to insects), this type of perception is usually referred to as contact chemoreception rather than taste.
Insects (insect) can perceive chemicals on dry surfaces. In this respect, their sense of taste differs from that of vertebrates, which generally perceive compounds in solution. Chemicals on the surface of another insect or on the surface of a leaf are not in solution and are probably conveyed from the insect or leaf surface by carrier proteins in the material covering the nerve endings at the pore.
 Olfactory receptors in arthropods are largely restricted to feelerlike structures at the front end of the animal. In crustaceans (crustacean) most multiporous hairs are on the antennules, and in insects they are on the antennae. However, in arachnids (arachnid) multiporous hairs occur in different positions in different groups. The olfactory receptors of scorpions (scorpion) are found in structures called pectines that project from the ventral surface of the second segment of the opisthosoma, and in sunspiders (sunspider) they are found in small flaps of cuticle called malleoli that hang beneath the basal segments of the legs. However, whip spiders and whip scorpions have the first pair of walking legs modified to form antenna-like structures that are extended in front as them as they move. Multiporous hairs are present on these antenniform legs. Some spiders (spider) are known to have a sense of smell, but the receptors have not been identified.
Olfactory receptors in arthropods are largely restricted to feelerlike structures at the front end of the animal. In crustaceans (crustacean) most multiporous hairs are on the antennules, and in insects they are on the antennae. However, in arachnids (arachnid) multiporous hairs occur in different positions in different groups. The olfactory receptors of scorpions (scorpion) are found in structures called pectines that project from the ventral surface of the second segment of the opisthosoma, and in sunspiders (sunspider) they are found in small flaps of cuticle called malleoli that hang beneath the basal segments of the legs. However, whip spiders and whip scorpions have the first pair of walking legs modified to form antenna-like structures that are extended in front as them as they move. Multiporous hairs are present on these antenniform legs. Some spiders (spider) are known to have a sense of smell, but the receptors have not been identified.The number of multiporous hairs is usually large, since the greater the number, the greater the chance that molecules (molecule) in low concentrations in the air or water will make contact with a sensillum. In insects the length or complexity of the antennae is a reflection of the numbers of multiporous sensilla. In insects requiring increased sensitivity, the antennae are branched, providing a larger surface area on which more sensilla can be accommodated. The featherlike (plumose) antennae of some male moths (moth), compared with the slender antennae of females of the same species, provide a high degree of surface area and thus a high degree of sensitivity. For example, in the polyphemus moth a male with plumose antennae has over 60,000 multiporous sensilla on one antenna, whereas a female with slender antennae has only about 13,000 sensilla on a single antenna. Each of the multiporous hairs contains the dendrites of two or more olfactory receptor cells, and the total number of receptor cells may be very large. An adult male cockroach can have as many as 195,000 olfactory receptor cells on one antenna, and an adult male tobacco hornworm moth may have from 100,000 to more than 300,000 receptor cells on one antenna. Some crabs have similar numbers of olfactory receptor cells on their antennules.
The axons from the olfactory receptor cells run to the central nervous system, where the axons from all the cells with similar sensory properties converge to a single glomerulus, similar to vertebrates. The position of the clusters of glomeruli forming the olfactory lobe varies in the different groups of arthropods according to the body segment on which the multiporous receptors occur. In insects and crustaceans the glomeruli clusters are in the brain, but in arachnids (arachnid) the clusters occur in more-posterior parts of the central nervous system. In addition, the number of glomeruli varies between species. A mosquito has about 10 glomeruli on each side of its brain, whereas a grasshopper has about 1,000 glomeruli in total. A male cockroach has about 125 glomeruli, and a male tobacco hornworm moth has about 60 glomeruli. On average, about 1,500 axons from olfactory receptor neurons converge on each glomerulus in the cockroach, and about 5,000 axons converge on each glomerulus in the moth. These average convergences are high, but much lower than in vertebrates (25,000 axons per glomerulus), although some individual glomeruli in insects may connect with many more axons. For example, in the male tobacco hornworm moth, about 60,000 olfactory receptor cells respond to one component of the female pheromone. The axons of all these cells converge on one large glomerulus, called a macroglomerular complex, resulting in roughly 60,000 axons connecting to a single glomerulus.
Each olfactory receptor cell in arthropods seems to express only one type of receptor protein, similar to vertebrates. As a result, each cell responds to a specific chemical. This is best illustrated by cells that respond to sex pheromones, in which a difference in the position of a double bond between two carbon atoms (atom) can be distinguished.
Many arthropods are able to respond to and differentiate between a wide range of chemical compounds, including pheromones and food-related odours. Many terrestrial species can perceive a range of common compounds with six or seven carbon atoms that are produced by all green plants as metabolic by-products. Bloodsucking insects and some plant-feeders have cells that respond to carbon dioxide, which in blood feeders can provide an important cue to the presence of a host. The characteristic odours of many plants can be perceived and, depending on the insect species, may cause an insect to be attracted to or repelled by the plant. Arthropods also perceive a wide range of odours that have no obvious direct relevance to their lives. This ability is probably necessary for developing learned associations between odours and important but unpredictable factors in the animals' lives.
Vertebrates (vertebrate)
fish
Similar to other vertebrates, fish have discrete taste and smell systems; however, since they live in water, the taste system is not confined to the oral cavity. For example, taste buds occur on the lips, the flanks, and the caudal (tail) fins of some species, as well as on the barbels of catfish. Regardless of where the taste buds occur on the body, they are connected to neurons (neuron) in the same three cranial nerves (facial, glossopharyngeal, and vagus) as the taste buds in the oral cavity. In addition to the taste buds, isolated (solitary) chemoreceptor cells are scattered over the surface of fish. These cells have a similar structure to that of individual taste receptor cells, but their connections to the brain or spinal cord arise from the nerves' providing innervation for the particular part of the body in which the cells occur. Although these cells are isolated from each other, they may occur in densities as high as 4,000 cells per mm2.
The olfactory system of fish is independent of the respiratory system, which is unlike that of terrestrial vertebrates. Gas exchange in fish occurs via the gills (gill), which are bathed in a continual flow of water coming through the mouth. The nasal (olfactory) cavities of sharks (shark) (elasmobranchs) are pits, one on each side of the ventral surface of the snout, located just in front of the mouth, whereas in bony fish (teleosts (teleost)) the pits are usually on the dorsal side of the head, in front of the eyes. Each pit opens to the exterior through anterior and posterior nares; there is no connection with the oral cavity. Water flows into the nasal cavity through the anterior nares and out of the nasal cavity through the posterior nares. In garfish and puffer fish (puffer), the flow is maintained by the action of cilia on accessory cells in the olfactory epithelium. In contrast, in rockfish and some other benthic fish, the volume changes produced in the mouth by respiratory movements compress and expand accessory chambers that are associated with the olfactory epithelium, causing water to move into and out of the nasal cavity. The “coughing” exhibited by certain fish such as flounder cleans the gills and results in an active irrigation of the olfactory epithelium by changing the volume of the nasal cavity. The frequency of coughing increases in the presence of food odours, suggesting that this behaviour may be analogous to sniffing in terrestrial vertebrates.
The floor of the nasal cavity is composed of folds (lamellae) that often form a rosette, with the lamellae radiating from a central point. The effect of the lamellae is to increase the surface area of the olfactory epithelium that lines the nasal cavity. As with terrestrial vertebrates, the number of olfactory receptor cells may be very large, up to 10 million. The axons of olfactory receptor cells run back to glomeruli in the olfactory bulb of the brain. Terrestrial vertebrates appear to have fewer glomeruli than fish. zebra fish, commonly used in laboratory studies, have about 80 glomeruli in each olfactory bulb, and the mitral cells, which synapse with the axons of receptor cells in the glomeruli, have axons extending to several glomeruli, whereas in mammals (mammal) the main connection of each mitral cell is with one glomerulus. Axons from the olfactory bulb form two main tracts, and these may reflect functional differences that in terrestrial vertebrates become separated as the olfactory and vomeronasal systems.
Terrestrial vertebrates
In terrestrial vertebrates the taste receptor system is generally confined to the oral cavity. However, tadpoles (tadpole), being aquatic, retain the external solitary chemosensory cells found in fish, whereas adult amphibians (amphibian) lack these cells. This indicates that the chemoreceptor system of amphibians reflects their evolutionary position as terrestrial animals that are still dependent on an aquatic environment for breeding. The olfactory system is directly associated with the intake of air during breathing and thus is almost continuously exposed to environmental odours. In addition, most terrestrial vertebrates have a third group of chemoreceptors that form the vomeronasal organ (Jacobson organ (Jacobson's organ)). This is a bony or cartilaginous capsule in the nasal cavity, one on each side of the nasal septum. The lumen of the capsule opens through a duct into the nasal cavity or, in some animals, connects with the oral cavity through an opening in the palate. The capsule is filled with fluid and is lined on one side by ciliated receptor cells. The axons from these cells extend to glomeruli, which are separated from those of the primary olfactory system, forming an accessory olfactory bulb. In contrast to the olfactory system, axons from one type of receptor cell project to different glomeruli, and each glomerulus receives input from several types of receptor. In some salamanders (salamander) and rats (rat) the vomeronasal organs are larger in males than in females.
Two families of genes (gene) are concerned with producing receptor proteins in the vomeronasal system. These gene families are different from the primary olfactory gene family. In the mouse there are only 200–300 genes associated with producing vomeronasal receptor proteins. As with other vertebrate chemical receptor proteins whose structures are known, the receptor proteins of the vomeronasal system have seven transmembrane domains. Unlike the receptor cells of the taste and olfactory systems, vomeronasal receptor cells adapt slowly, or sometimes not at all, when continuously stimulated; therefore, the transfer of information to the brain is maintained.
In contrast to the primary olfactory system, in which molecules (molecule) are conveyed to the receptors as an inevitable consequence of breathing, transfer of stimulants to the vomeronasal organ is actively regulated. In addition, different animals exhibit different stimulant regulation mechanisms. Both volatile and nonvolatile compounds may be perceived, though the perception of nonvolatile compounds requires that the animal make direct contact with the source using its nose or tongue. Lungless salamanders (lungless salamander) (family Plethodontidae) rely on the vomeronasal organ (Jacobson's organ) for habitat selection and mating, using the snout to make deliberate contact with the object being investigated. These animals have a narrow groove close to each nostril that connects the upper lip with the nostril. During nose tapping, fluid moves along the grooves by capillary action and is driven, possibly by ciliary movement, into the extensive vomeronasal organs. In another group of amphibians, the burrowing wormlike caecilians (Gymnophiona), chemicals are carried to the vomeronasal organs via tentacles. Directly in front of each eye is a small pore leading to a sac that contains a tentacle. The tentacle can be extended through the pore by hydrostatic pressure to make contact with the surrounding soil. A duct connects the tentacular sac with the vomeronasal organ, and it is believed that this is the path along which chemicals are transported. The connection of the vomeronasal organ to the main olfactory epithelium is greatly reduced in these animals.
 In snakes (snake) and lizards (lizard) the vomeronasal organ is completely isolated from the nasal cavity. As a consequence, environmental chemicals can enter the organ only via the mouth, and the tongue plays an essential part in chemical transport. In snakes (snake) there are no taste buds on the tongue, and chemical transport is probably one of the tongue's major roles. When snakes and lizards flick their tongues in and out, the tongue moves through a vertical arc. In lizards each extension of the tongue usually involves only one such movement, and the lower surface of the tongue often touches the substrate in front of the lizard. However, the tongues of snakes usually make 3–5 vertical oscillations at each extension, and the tongue usually does not touch the substrate. These movements are rapid, being completed in little more than half a second. (Snakes also make much slower tongue flicks that may serve as warning signals.) It is assumed that, during tongue flicking, odour molecules are trapped in the salivary coating of the tongue, and from there they are transferred to the opening of the vomeronasal organ. Various hypotheses have been put forward to account for the transfer of chemicals from the tongue to the vomeronasal organ, which must occur very quickly; however, the mechanism remains unknown.
In snakes (snake) and lizards (lizard) the vomeronasal organ is completely isolated from the nasal cavity. As a consequence, environmental chemicals can enter the organ only via the mouth, and the tongue plays an essential part in chemical transport. In snakes (snake) there are no taste buds on the tongue, and chemical transport is probably one of the tongue's major roles. When snakes and lizards flick their tongues in and out, the tongue moves through a vertical arc. In lizards each extension of the tongue usually involves only one such movement, and the lower surface of the tongue often touches the substrate in front of the lizard. However, the tongues of snakes usually make 3–5 vertical oscillations at each extension, and the tongue usually does not touch the substrate. These movements are rapid, being completed in little more than half a second. (Snakes also make much slower tongue flicks that may serve as warning signals.) It is assumed that, during tongue flicking, odour molecules are trapped in the salivary coating of the tongue, and from there they are transferred to the opening of the vomeronasal organ. Various hypotheses have been put forward to account for the transfer of chemicals from the tongue to the vomeronasal organ, which must occur very quickly; however, the mechanism remains unknown.In male ungulates (ungulate), cats (cat, domestic), elephants (elephant), bats (bat), and some other mammals, access to the vomeronasal organ may be facilitated by curling the lips and exposing the upper teeth, with the nostrils closed. This is called flehmen and is seen during courtship, when it is used by males to assess the estrus state of females, and during the investigation of new odours, when it is used by both males and females to explore their surroundings. Changes in the internal volume of the vomeronasal organ, produced by dilation and compression of blood capillaries, are believed to enhance fluid movement and molecule transport into the lumen. In antelope that exhibit flehmen behaviour, a groove on each side of the hard palate leads to a duct connecting the oral cavity to the vomeronasal organ. hartebeest and topi, animals that do not exhibit flehmen, lack oral connections to their vomeronasal organs.
The vomeronasal organ (Jacobson's organ) is involved in pheromone perception, prey recognition, and habitat selection. Animals such as birds (bird) and the great apes (ape) do not have vomeronasal organs, and in these animals pheromones are of little or no importance. Even in animals that do possess vomeronasal organs, the olfactory system is involved in pheromone perception. A vomeronasal organ does start to develop in human embryos (embryo), and it is present in most, if not all, adults. Its evolutionary development is foreshadowed in fish, in which the vomeronasal gene families are present but are expressed together with olfactory receptor genes in the olfactory epithelium. There is evidence that the nerve pathways from the different receptor types are distinct, though overlapping, in fish.
The significance of the vomeronasal system is that it separates the nervous pathway dealing with innate behavioral and physiological responses from the olfactory pathway that communicates with higher centres of learning and cognition.
Behaviour and chemoreception
Many aspects of animal behaviour involve the perception of chemicals that arise from the environment, such as chemicals produced by plants or predators (predation), or that arise from other members of the same species (pheromones). Because many compounds are volatile, they provide the means for detecting a mate or food from a distance and can serve as an alternative to or work in conjunction with vision and sound. Some odours may have repellent effects. Volatile compounds are perceived via the olfactory system and sometimes via the vomeronasal system. Nonvolatile chemicals are perceived via taste or, in terrestrial vertebrates, via the vomeronasal organ. For the perception of nonvolatile chemicals to be effective, the animal must make direct contact with the chemical's source. These chemicals may have a positive, activating effect on a particular behaviour, or they may have a negative, inhibitory effect. Chemicals that function as signals between organisms of the same or different species are often referred to as semiochemicals. These chemicals may be by-products of basic metabolic pathways, such as alcohols (alcohol) and terpenes (terpene) produced by green plants (plant) or lactic acid produced by mammals. In other cases these chemicals may be specifically produced to provide ecological signals. Some organisms have exocrine glands specifically designed for the production of semiochemicals.
Pheromones (pheromone)
Chemicals produced by an animal (animal communication) to affect the behaviour or physiology of another member of the species are called pheromones (pheromone), and at least some species in all the major animal groups are known to produce pheromones. These chemicals attract a potential mate from a distance, have specific sex or kin recognition, and involve many aspects of social behaviour. Among mammals, pheromones may provide information about sex, age, genetic similarity, reproductive state, sexual arousal, dominance status, territorial boundary, time of last marking, and even emotional state, such as fear or anger. A pheromone may consist of a single compound but usually involves a mixture of different compounds. For the most part, the individual chemicals are not unique to the organism producing them, although the combinations of chemicals may be unique.
Pheromones may be categorized as releasers and primers. A releaser pheromone has an immediate effect on the behaviour of the recipient, whereas a primer pheromone affects the recipient's physiology, producing an effect on behaviour after a period of time. Releaser pheromones are perceived by chemosensory neurons (neuron) in the recipient's peripheral nervous system. This is probably also true of primers, although this is not always known. It is possible that in some cases primers have a direct effect on an animal's metabolism after being taken into the body.
The characteristics of a compound or suite of compounds employed as a pheromone are determined by the pheromone's function and the context in which it is used. To have an effect at a distance from the producer, the compound must be volatile, enabling it to be readily dispersed. In general, within a class of compounds, smaller molecules are more volatile than larger ones. For example, ethanol (ethyl alcohol) (C2H5OH) is about 100 times more volatile than hexanol (C6H13OH) and about 10,000 times more volatile than undecanol (C11H23OH), and formic acid (HCOOH) is about 100 times more volatile than pentanoic acid (C4H9COOH) and 10,000 times more volatile than octanoic acid (C7H15COOH). On the other hand, larger, nonvolatile compounds may be important when animals are in close contact, when taste is important.
A second critical feature of many pheromones is specificity. A sex-attractant pheromone would be disadvantageous if it also attracted individuals of other species. Specificity is dependent to some extent on the degree to which a particular molecular structure can be modified; for example, there are more possible permutations of the structure of a molecule with a backbone of 10 carbon atoms than of a molecule with a backbone of only 2 carbons. The need for volatility may conflict with the need for specificity, and the animal may need to compromise (in an evolutionary sense) to produce molecular structures that meet both requirements. Distance-attractant pheromones require both volatility and specificity. For example, the sex-attractant pheromones of most moths (moth) are molecules containing 12, 14, 16, or 18 carbon atoms, and the aggregation pheromones of bark beetles (bark beetle), which attract huge numbers of conspecifics (members of the same species), comprise molecules with about 8–10 carbon atoms.
An alternative way to achieve specificity is to use mixtures of compounds and to vary the relative proportions of the components. An example of this is seen in moths of the genus Spodoptera. Numerous species in this genus have sex-attractant pheromones with 14-carbon atom compounds, but all these species produce more than one compound, and some are known to produce more than seven compounds. The compounds differ primarily in the presence or absence and position of double bonds located between the carbon atoms that form the backbone of the molecule. By using different proportions of the same compounds, each species can produce its own specific odour. This approach makes it possible to achieve not only species specificity but also individual specificity within a species, which is important in social contexts. Large numbers of compounds, often more than 50, in secretions of the preorbital and pedal glands of antelope and the urine of many mammals appear to reflect the need for individual specificity. Social hymenopterans (hymenopteran) use cuticular hydrocarbons (hydrocarbon) in kin recognition, and there may be 20 or more such compounds on the surface of a single insect.
Alarm pheromones, produced by some animals and best known in insects, have quite different requirements. An alarm pheromone needs high volatility, since it is used to quickly warn other individuals and must rapidly decay from the immediate environment. With a persistent compound the insects would be in a continual state of alarm or would habituate to the odour, thus reducing its value as an alarm pheromone. On the other hand, an alarm pheromone does not require a high degree of specificity, since it is usually not a disadvantage if other species detect the odour. As a consequence, very small molecules are used as alarm pheromones. In formicine ants (ant), formic acid (HCOOH) often serves this function, and, in general, the alarm pheromones of ants and bees (bee) are compounds with 5–9 carbon atoms.
 Marking pheromones require characteristics opposite those of alarm pheromones, since their function is to convey a signal to other members of the species for a relatively long term. Thus, they demand some persistence, though not so much that they remain when their utility is past. Trails marked by pheromones are commonly produced by worker ants as they return to the nest from foraging. The trail persists as long as the food source that it is connected to remains available and as long as the trail pheromone is reinforced by the returning workers. The territorial marks (territorial behaviour) of vertebrates are also maintained by periodic reinforcement. Persistence can also be achieved in other ways. The persistence of territorial marks made by tigers (tiger) is aided by the presence in the pheromone mixture of compounds that delay the loss of volatile compounds. The marking scents of skunks (skunk), which are also used for defense (see below Behaviour and chemoreception: Defensive odours (chemoreception)), may retain persistence by incorporating a chemical that breaks down slowly to produce the dominant effective compound.
Marking pheromones require characteristics opposite those of alarm pheromones, since their function is to convey a signal to other members of the species for a relatively long term. Thus, they demand some persistence, though not so much that they remain when their utility is past. Trails marked by pheromones are commonly produced by worker ants as they return to the nest from foraging. The trail persists as long as the food source that it is connected to remains available and as long as the trail pheromone is reinforced by the returning workers. The territorial marks (territorial behaviour) of vertebrates are also maintained by periodic reinforcement. Persistence can also be achieved in other ways. The persistence of territorial marks made by tigers (tiger) is aided by the presence in the pheromone mixture of compounds that delay the loss of volatile compounds. The marking scents of skunks (skunk), which are also used for defense (see below Behaviour and chemoreception: Defensive odours (chemoreception)), may retain persistence by incorporating a chemical that breaks down slowly to produce the dominant effective compound.Mixtures of compounds have the potential to provide greater information than single compounds. This appears to be true of some antelope markings that change with time, enabling the recipient to adjust its behaviour appropriately. Leafcutter ants (leafcutter ant) (genus Atta) have alarm pheromones consisting of four components with different volatilities. Coupled with differences in the sensitivity of worker ants, the different volatilities produce different areas over which the compounds are most effective, and they stimulate different behaviours. Hexanal, with the greatest effective area, alerts worker ants, and hexanol has an attractant effect. In contrast, 3-undecanone and 2-butyl-2-octenol, the least volatile and thus most concentrated closest to the pheromone source, initiate biting behaviour.
pheromone perception
The specificity of pheromones depends on the specificity of perception as well as production. Little is known of the physiology of individual receptor cells outside the insects, which have receptor cells that are highly specific, at least for the major pheromone components. In many cases, when an attractant pheromone has two major components, the recipient has large numbers of cells specific to each of the compounds, often in the same sensillum. Very often the cells are extremely sensitive, enabling the animal to respond to very low concentrations of compounds.
Primer pheromones
Primer pheromones are important in aspects of social physiology in a range of animals. In mammals they are influential in coordinating reproductive (reproductive behaviour) physiology, and compounds excreted in the urine are especially important (see below Behaviour and chemoreception: Mammals (chemoreception)). For example, the physiology of female mice (mouse) is affected by the odour of urine produced by males and other females. Dominant males have the greatest effect, causing the release of luteinizing hormone in the female, which leads, together with contact with the male, to ovulation. In contrast, the urine of other females tends to delay ovulation. In the presence of a male, a female increases the rate at which she produces urine, and this causes the release of testosterone in the male.
Comparable pheromones are produced by locusts (locust). A mature male desert locust produces a maturation pheromone from glands scattered throughout the epidermis. The pheromone can act via the olfactory system of the recipient or, if the insects come into contact, via the contact chemoreceptor system, although this is not known with certainty. The pheromone speeds up sexual maturation by affecting the endocrine system in individuals of both sexes, with the result that in a swarm of locusts sexual maturation tends to be synchronized.
Primer pheromones are especially important in the maintenance of colony structure in social insects (social insect). Queen honeybees (honeybee) secrete “queen substance” from their mandibular glands. When an unfertilized queen leaves the colony, queen substance acts as an olfactory attractant for males. The same compound within the colony modifies the behaviour of workers, preventing them from rearing more queens, and also affects their physiology, disrupting the development of their ovaries (ovary).
Movement toward an odour source
Attraction to the source of an odour poses problems for all animals using the sense of smell. It had been supposed that animals simply moved up a concentration gradient, from an area of low odour concentration to an area of high odour concentration, ending near the source of an odour. However, consideration of the movement of odour molecules (molecule) in air or water showed that, in general, such gradients do not exist under natural conditions. Wind flow varies in both direction and strength. In addition, during the day, when the ground is heated, rising and falling air movements contribute to turbulence. As a result, odour molecules, even when continuously released at the source, become dispersed as a series of wisps, similar to the way that smoke from a chimney becomes dispersed. As a consequence, a stationary animal (animal communication) or an animal moving toward an odour source in a straight line will encounter bursts of odour with relatively long intervals between bursts. This is true whatever the distance from the source, although at short distances bursts contain more peaks with high concentrations of odour molecules. Only by averaging the concentration over a period of time and distance is it possible to follow a gradient of odour. Some animals may do this, but insects (and probably many other organisms) use a different strategy. In these organisms an odour has the effect of switching on a behavioral program that uses some signal other than odour to locate the source. In many cases the other signal is wind direction, and the animal moves upwind, ultimately arriving at the source of an odour. This mechanism is called odour-modulated anemotaxis. It is used by male moths to locate females, by moths flying to a flower odour to obtain nectar, and by cabbage root flies flying toward a cabbage plant (cabbage) to lay eggs.
Wind direction may be determined by its mechanical effect on the body, and in insects this involves structures at the bases of the antennae and mechanosensory hairs on the head. The behaviour involved in moving upwind varies. Larval insects such as those of the desert locust walk directly upwind if they smell food after having been without it for some time, and adult golden rod beetles exhibit similar behaviour. Cabbage root flies, when they perceive the host odour, orient into the wind while still on the ground and then make a short, straight flight of perhaps one metre before landing. The arrival of a new puff of odour causes them to reorient to the wind and repeat the process. Thus, their movement toward the odour source involves a series of short flights. However, in many insects (insect) odour causes takeoff into the wind, followed by a zigzagging flight toward the source, much as a sailboat might tack into the wind. During most of the movement, the insect is flying across the wind with its body oriented obliquely upwind. As a result, it drifts sideways, as an airplane does in high winds. This sideways drift produces a flow of images of the ground across the insect's eyes, and the insect adjusts its power output to maintain its general upwind movement. The same mechanism is used by nocturnal insects. However, in some day-flying insects such as tsetse flies (tsetse fly), the flight toward an odour source may be much more direct, with the odour causing takeoff but flight being directed toward any moving object that is visible upwind.
Odour gradients, in which the concentration declines progressively with increasing distance from the source, probably do exist in very still environments such as those occurring in the soil. The soil-dwelling larvae of some insects that feed on roots, such as the corn root worm (the larva of a beetle), have been shown to move along chemical gradients.
reproductive behaviour
Insects
Sex-attractant pheromones
 Many insects produce a sex-attractant pheromone, by which one sex attracts the other from a distance. Among moths (moth), it is common for the female to produce a sex-attractant pheromone. For example, female gypsy moths (gypsy moth), which are flightless despite having fully developed wings, and female bagworms (bagworm moth), which do not have wings, depend wholly on the power of their sexual odour to attract a mate. Female moth sex-attractant pheromones are produced in glands in the moth's abdomen. When the female is ready to mate, she exposes the glands and disperses the pheromone into the air. This behaviour, known as calling, typically occurs at a time of day or night that is characteristic of the mating pattern of the species.
Many insects produce a sex-attractant pheromone, by which one sex attracts the other from a distance. Among moths (moth), it is common for the female to produce a sex-attractant pheromone. For example, female gypsy moths (gypsy moth), which are flightless despite having fully developed wings, and female bagworms (bagworm moth), which do not have wings, depend wholly on the power of their sexual odour to attract a mate. Female moth sex-attractant pheromones are produced in glands in the moth's abdomen. When the female is ready to mate, she exposes the glands and disperses the pheromone into the air. This behaviour, known as calling, typically occurs at a time of day or night that is characteristic of the mating pattern of the species.Sex-attractant pheromones can sometimes have unfortunate side effects for the insect producing them, because they can be used by other organisms to locate the insects. For example, males of the stinkbug genus Podisus produce a pheromone that attracts females as well as other males and immatures. It also attracts female parasitic (parasitism) flies of the family Tachinidae (tachinid fly), providing the flies with an easy way to find their hosts, on which they lay their eggs. In some instances other organisms produce some of the sex-attractant pheromones of moths to mislead the moths. Late-stage immature and adult female bolas spiders (spider) in the genus Mastophora are known to produce some of the same components of the sex-attractant pheromone produced by females of some noctuid moths (owlet moth). The spider is active at night and hangs from a horizontal silk line. It produces a vertical thread, which it holds with one leg, and secretes a viscous fluid that forms a globule at the lower end of the thread. Male moths are attracted by the odour of what appears to be a potential mate, and the spider, apparently stimulated by the vibrations of the moth's wings, uses its leg holding the thread to hurl the viscous globule at the moth. If the globule hits the moth, the moth becomes trapped, and the spider immobilizes it with venom by attaching the vertical thread to the horizontal line and moving down the thread or by pulling the thread up. The moth may be eaten immediately or wrapped in silk before being eaten.
 Some orchids (orchid) produce chemicals that mimic the sex-attractant pheromones of the wasps (wasp) that pollinate them. In this instance the orchid flower also bears some visual resemblance to the female, giving rise to some of the common orchid names—for example, bee orchids. The male is first attracted by the odour and then attempts to copulate with the presumed female. The dummy female is positioned in such a way that the male picks up the pollen-containing masses, known as pollinia, on its head before flying off.
Some orchids (orchid) produce chemicals that mimic the sex-attractant pheromones of the wasps (wasp) that pollinate them. In this instance the orchid flower also bears some visual resemblance to the female, giving rise to some of the common orchid names—for example, bee orchids. The male is first attracted by the odour and then attempts to copulate with the presumed female. The dummy female is positioned in such a way that the male picks up the pollen-containing masses, known as pollinia, on its head before flying off.Aphrodisiac pheromones
The males of some insects produce aphrodisiac pheromones that induce females to mate once the two sexes have come together. One of the most remarkable and fully understood examples of this concerns monarch butterflies (monarch butterfly) (although not the well-known North American monarch). Males of these insects seek out plants containing a particular type of alkaloid known as a pyrrolizidine, which is highly toxic to mammals. The insect licks the plant with its tongue and accumulates small quantities of the alkaloid. Concealed on either side of its abdomen are structures called hair pencils that contain the alkaloids and that are formed from modified scales (basically similar to those that cover the wings and other parts of the body, although different in form). The pencils, when everted out of the abdomen, separate to form elegant brushlike structures, somewhat resembling feather dusters. Eversion only occurs in the presence of the female, but before doing this the male thrusts the pencils (not yet expanded) into glandular pockets on the hind wings. The contents of the pockets effect a slight chemical modification of the alkaloid to produce the pheromone. Some of the scales break into minute fragments impregnated with the pheromone, and these fragments are dusted onto the female antennae as the male hovers over the female during courtship. The odour of the pheromone, perceived by cells on the female's antennae, induce her to permit the male to copulate.
Sex recognition
In houseflies (housefly) and their relatives, compounds in the layer of wax covering the outside of the insect are important in sexual recognition. Males and females have different chemical profiles that allow a male to distinguish unmated from mated females. In tsetse flies, some of the male's wax rubs off onto the female during mating, and this changes her wax chemistry so that she is no longer attractive. Females of the vinegar fly, Drosophila, lose their attractiveness after mating by secreting wax with a different chemical profile.
Mammals (mammal)
Pheromones are also of great importance in reproduction (reproductive behaviour) among mammals, acting both as releasers, thereby influencing behaviour, and as primers, thereby altering the physiology of other members of the same and the opposite sex. Among rats and mice, and probably many other species, odours from the urine have a major role. Mammalian urine contains many different volatile compounds. For example, over 60 volatile compounds have been identified in the urine of the house mouse and the white-tailed deer. By repeated marking, house mice produce accretions of urine at “marking posts,” and a dominant male may mark 100–200 times in an hour. It is probable that mixtures of these compounds are important in individual recognition, but specific compounds may also be important.
territorial behaviour
Territorial behaviour occurs in many animals and is especially widespread in mammals. Both visual and chemical signals may be used to advertise the territory to other animals. Antelope have a variety of exocrine glands, the secretions of which may be used in communication. However, the preorbital glands, located on the side of the face with an opening just in front of the eyes, are the best known in relation to territorial behaviour. In species such as the South African bontebok (blesbok), the preorbital glands are larger in males than in females. The secretions of these glands are extremely complex, containing over 40 compounds, and are deposited on grass culms (stems) or twigs at territory borders by pressing the head down onto the culm so that it enters the opening of a pore, alternating between left and right glands. In species such as Thomson's gazelle, this results in an appreciable accumulation of the secretion on the grass or twig. Bontebok appear to transfer the secretion to their horns and forehead by waving the head from side to side across the stalk bearing the secretion.
For scents (odour) to be effective as territorial markers, individuals must be able to distinguish their own scent from the scents of other species and from the scents of individuals of the same species. The scents must persist for some time and must also change with time, enabling a recipient to judge whether a scent derives from a recent intruder or a past intruder. The complexity of the secretions probably contributes both to individual variation and to changes with time. It is likely that volatile components are lost more rapidly than nonvolatile components, causing the quantitative composition of the scent to change in a predictable way.
In addition to scent marking from the preorbital glands, many antelope mark territorial boundaries with fecal middens. These serve both as visual markers and as substrates for glandular secretions. Animals often urinate at the same time that they defecate. In addition, territorial male bontebok paw dung patches, possibly adding the secretion of the pedal glands to the dung. Similar to the preorbital gland secretions, the pedal gland secretions are very complex, and bontebok contain over 80 compounds of different classes. Territorial males habitually defecate at the same sites, and they do so frequently. Male oribi may defecate up to eight times in an hour, presumably to maintain the odour quality of the middens.
Carnivores (carnivore) also mark their territories by scent. Civets (civet), found in Africa, southern Europe, and Asia, secrete material from anal glands. The major ingredient, called civet, or civetone, is an unusual compound, with 17 carbon atoms that form a ring. musk deer produce a similar compound (with 15 carbon atoms in a ring), and both compounds were widely used in perfumery until similar synthetic compounds were produced.
Little is known about the perception of chemical marker compounds, although the vomeronasal organ (Jacobson organ (Jacobson's organ)) is suspected to play an important role. As with sex-attractant pheromones, marking pheromones can provide cues that animals use to locate prey or hosts. For example, the klipspringer, a South African antelope, is the host for a bloodsucking tick called Ixodes matopi. The antelope marks its territory with secretion from its preorbital gland, and adult ticks aggregate on these marks, presumably using odour to find them. This behaviour increases their chances of finding the appropriate host.
Individual recognition
Among social animals it is very common for individuals to be able to recognize each other, and chemoreception plays an important role in this behaviour. Social insects (social insect), such as termites (termite), bees (bee), wasps (wasp), and ants (ant), are able to distinguish between nest mates and individuals from other colonies. This often depends on small differences in the proportions of different components in the insects' surface wax. Social wasps make their nests of paper, which is produced by chewing wood. Some of the wax rubs off the bodies of the workers and onto the nest. The composition of this wax plays a key role in enabling workers to distinguish members of their own colony from intruders. Other insects called inquilines, which habitually live with ants, depend on acquiring the wax characteristics of the ant colony in order to avoid being attacked by the ants.
In mammals, individual recognition is often achieved via the odour of urine. Urine and other body odours are partly controlled by genes (gene) in the major histocompatibility complex (MHC), which also governs certain immune responses. Mice have about 50 linked genetic variations (polymorphisms (polymorphism)) in this complex. Some of the proteins (protein) produced by these genes occur in the urine and contribute to the chemical signature of each individual. However, because the proteins are not volatile, they cannot contribute directly to the odour, and their precise role is not understood. In rats, bacteria from the gut play a key role in the development of odour specificity. This does not appear to be the case in mice. Rats, mice, and humans prefer the odours of individuals with a histocompatibility complex different from their own; thus, mating tends to occur between individuals with different MHCs. In order to detect different MHCs, an individual must be aware that a potential partner has a distinct smell. In mice the odour of the family in which they are reared becomes imprinted early in development. ( imprinting is the process by which young animals develop a lasting association with a particular feature in the environment.) If a pup is reared by a foster mother with her own pups, the pup imprints onto the odour of the foster family. This family odour is the odour against which the pup will compare the odour of a potential mate, once the pup is mature. This means that the pup does not make the comparison with its own genetically determined odour.
homing
Many animals have specific places, such as nests or dens or, on a larger scale, geographical locations, to which they return periodically, often to breed. This homing behaviour may involve vision or an electromagnetic sense. However, in some animals olfaction plays a significant role, often in conjunction with one of the other senses. These instances depend on a learned knowledge and memory of environmental odours, although, despite multiple studies, in no case has the nature of the odour been well characterized. Animals known to use odour in homing include fishes (fish), reptiles (reptile), amphibians (amphibian), and birds (bird).
 salmon breed in fresh water, usually in the upper reaches of streams or in lakes. They remain in fresh water, generally for a year or more, varying to some extent with the species, and then they migrate to the sea. They remain in the sea to feed, often for two or three years, before returning to fresh water to breed. The most extraordinary aspect of this migration is that the vast majority—more than 90 percent—of fish return to the streams in which they passed their early development. This is important because, over many generations, the fish become adapted to the particular characteristics of their home stream, increasing the probability that the young will survive. (Today, because of a number of environmental factors, such as dams and overfishing, the number of fish returning to their home streams is decreasing.)
salmon breed in fresh water, usually in the upper reaches of streams or in lakes. They remain in fresh water, generally for a year or more, varying to some extent with the species, and then they migrate to the sea. They remain in the sea to feed, often for two or three years, before returning to fresh water to breed. The most extraordinary aspect of this migration is that the vast majority—more than 90 percent—of fish return to the streams in which they passed their early development. This is important because, over many generations, the fish become adapted to the particular characteristics of their home stream, increasing the probability that the young will survive. (Today, because of a number of environmental factors, such as dams and overfishing, the number of fish returning to their home streams is decreasing.)The factors involved in directing the salmon to the correct stream system from the sea are not known, although geomagnetic orientation (geomagnetic field) or steering by the position of the Sun may be involved. However, once the fish has entered its own stream system, olfaction is involved in finding the original spawning sites. During early development, the chemical characteristics of the home stream become imprinted on the young salmon. The chemicals arise from the substrate and vegetation of the stream and from the immediate environment—factors that give every stream a specific chemical signature. In addition, chemicals produced by other salmon may contribute to the chemical signature of a stream, since chemical production is known to vary between salmon populations. Research has shown that Coho salmon can become imprinted to specific chemicals.
imprinting only occurs during a specific period of an animal's life and is usually thought to be something that occurs in the animal's brain. However, in the case of salmon, changes in the sensitivity of the olfactory receptors (receptor) are important, but the increase in sensitivity to the environment-specific odour does not occur until the salmon is ready to return to its home waters, two or more years after imprinting occurred.
Two hormones (hormone) regulate these processes. The timing of imprinting coincides with an increase in activity of thyroxine, a thyroid (thyroid gland) hormone, early in the fish's life, and it is presumed that this leads to changes in the sensory cells of the olfactory epithelium. The level of this hormone in the blood depends partly on the age of the fish and partly on environmental conditions. As a result, the timing of imprinting may vary from place to place. However, the increase in sensitivity of these cells does not occur until the salmon makes its return journey. The timing coincides with an increase in levels of reproductive (reproductive behaviour) hormones in the fish's blood, and these hormones probably regulate the changes in the olfactory system. The increased sensitivity results, at least partly, from an increase in the activity of the second-messenger (second messenger) system involved in the transduction of a specific chemical signal into an electrical signal. Changes may also occur in the type or number of receptor proteins involved in detection of the chemicals, but this is not known with certainty. It is probable that homing by sea turtles (sea turtle) is dependent on imprinting of some chemical characteristics of the natal beach in the hatchling stages.
Homing pigeons (pigeon) use olfaction as part of their navigation system, apparently depending on trace amounts of gases. However, if they have previous experience of an area, they appear able to navigate using visual and geomagnetic signals alone. It is likely that other migrant birds (bird) also use their memories of odours in navigation.
Finding and recognizing food
A wide variety of odours from potential food resources are known to attract or repel animals from a distance. After location of a possible food item, the close-range odours and taste together determine acceptability, although, in many predators (predation) and most birds, visual cues tend to predominate. Each animal (animal communication) group and some species have particular characteristic preferences determined by the overall mixture of volatile and nonvolatile nutrients and nonnutrients. Choices are also influenced by varying nutritional needs and by experience.
There are some generalizations that can be made. For example, a preference for foods containing sugars (sugar) is common in herbivores (herbivore) and omnivores (omnivore) and uncommon in carnivores. An ability to taste substances perceived by humans as bitter may be used to detect substances that are poisonous after ingestion. This ability appears to be more highly developed in herbivores than in carnivores. Carnivores are stimulated by flavours (flavour) characteristic of animal protein, especially certain amino acids (amino acid) and their breakdown products. Extreme generalists, such as rats and some primates (primate), typically sample novel foods and then eat more of those foods or reject them, depending on postingestive effects. Most animals learn to use odour, flavour, or other cues to improve or balance their nutrient uptake and to reduce the intake of poisons. Specialists, such as koalas (koala) and monarch butterflies, tend to be specifically attracted to or stimulated by chemicals in their foods.
Different foraging strategies may involve correlated chemosensory characteristics. For example, relative to lizards that sit and wait for their prey, lizards (lizard) that are active foragers rely heavily on chemical discrimination using their vomeronasal organs. Herbivores generally discriminate against plants with high concentrations of plant secondary metabolites, such as alkaloids (alkaloid), phenols (phenol), and terpenoids (terpene). High levels of tannins (tannin), which are astringent to humans, are commonly deterrent to herbivores, and plants with alkaloids, which are often bitter to humans, tend to be rejected by herbivores. The tassel-eared squirrel, which hoards twigs of ponderosa pines (pine) for winter food, prefers to collect twigs low in α-pinene (a monoterpene). There are many such individual examples recorded, some of which may result from learning, but many of which are innate. Chemoreception in combination with behavioral responses has been best characterized among insects.
Plant chemicals

 The chemicals (chemical compound) in plants (plant) include a range of nutrient compounds, such as sugars, proteins, and lipids (lipid). In addition, plants produce a great variety of chemicals that are not derived from primary metabolic pathways and that have some ecological signaling function. These secondary compounds may be volatile and therefore may affect animals at some distance from the plant. In contrast, other compounds are nonvolatile and are not detected until the animal makes contact with (e.g., bites into) a plant. The compounds belong to several different chemical classes, including alkaloids, nonprotein amino acids, cyanogenic glycosides (glycoside), terpenoids, glucosinolates, and phenolic compounds. Many thousands of these compounds are known, and their distribution among plants is often limited to particular taxa. For example, the cabbage family is characterized by glucosinolates and their breakdown products, which include the volatile thiocyanates; lupines (lupine) contain quinolizidine alkaloids; onions (onion) contain thiosulfinates; and mint contains suites of monoterpenes. These compounds are largely responsible for the odours and flavours of these plants that are perceived by humans and other animals. Often a particular chemical is found only in a genus or a particular species of plant. Because these chemicals affect the behaviour and fitness of animals that encounter them, they are of major importance in determining the range of plants eaten by an animal.
The chemicals (chemical compound) in plants (plant) include a range of nutrient compounds, such as sugars, proteins, and lipids (lipid). In addition, plants produce a great variety of chemicals that are not derived from primary metabolic pathways and that have some ecological signaling function. These secondary compounds may be volatile and therefore may affect animals at some distance from the plant. In contrast, other compounds are nonvolatile and are not detected until the animal makes contact with (e.g., bites into) a plant. The compounds belong to several different chemical classes, including alkaloids, nonprotein amino acids, cyanogenic glycosides (glycoside), terpenoids, glucosinolates, and phenolic compounds. Many thousands of these compounds are known, and their distribution among plants is often limited to particular taxa. For example, the cabbage family is characterized by glucosinolates and their breakdown products, which include the volatile thiocyanates; lupines (lupine) contain quinolizidine alkaloids; onions (onion) contain thiosulfinates; and mint contains suites of monoterpenes. These compounds are largely responsible for the odours and flavours of these plants that are perceived by humans and other animals. Often a particular chemical is found only in a genus or a particular species of plant. Because these chemicals affect the behaviour and fitness of animals that encounter them, they are of major importance in determining the range of plants eaten by an animal.Plant-feeding (feeding behaviour) animals may be polyphagous, if they eat plants from several different families, oligophagous, if they are restricted to feeding on members of one plant family, or monophagous, if they feed on only one genus or one species of plant. These differences depend on plant chemistry and, to a very large extent, on what the animal smells and tastes.
Attraction
 plant odours are usually complex mixtures, but often they are characterized by particular chemicals. For example, the characteristic odour (to humans) of plants in the cabbage family is produced by sulfur-containing compounds called isothiocyanates. The odours of mint, lavender, and pine are dominated by different terpenoids. Since most insects are monophagous or oligophagous, the distinctness of the odours of different plants often enables them to locate their specific hosts from a distance. Flight toward host-plant odours is known to occur in a number of different butterflies, moths, flies, beetles, and aphids (aphid). The insect has receptor cells on its antennae that respond to the appropriate compounds. For example, the antennal sensory system of cabbage root flies responds strongly to isothiocyanates from cabbage but weakly to most of the disulfides produced by onions. In contrast, the antennal sensory system of the closely related onion fly responds strongly to disulfides from onions and weakly to isothiocyanates from cabbage. These differences in antennal response mediate the differences in movement by these insects toward their respective host plants.
plant odours are usually complex mixtures, but often they are characterized by particular chemicals. For example, the characteristic odour (to humans) of plants in the cabbage family is produced by sulfur-containing compounds called isothiocyanates. The odours of mint, lavender, and pine are dominated by different terpenoids. Since most insects are monophagous or oligophagous, the distinctness of the odours of different plants often enables them to locate their specific hosts from a distance. Flight toward host-plant odours is known to occur in a number of different butterflies, moths, flies, beetles, and aphids (aphid). The insect has receptor cells on its antennae that respond to the appropriate compounds. For example, the antennal sensory system of cabbage root flies responds strongly to isothiocyanates from cabbage but weakly to most of the disulfides produced by onions. In contrast, the antennal sensory system of the closely related onion fly responds strongly to disulfides from onions and weakly to isothiocyanates from cabbage. These differences in antennal response mediate the differences in movement by these insects toward their respective host plants. Many flowers (flower) produce characteristic odours that attract pollinators. These odours are blends of volatile secondary compounds, including terpenes, derivatives of fatty acids (fatty acid), and aromatic phenolic compounds. Receptors on the insects' antennae respond to these compounds. However, while the response to odours of foliage is often innate, the response to odours of specific flowers is for the most part learned, since most insect pollinators are not specific to particular plants. In order to enhance the probability that an insect will visit plants of the same species, the insect must associate the presence of nectar with the odour of the flower that it last visited. This improves the foraging efficiency of the pollinator and increases the chance of cross pollination within a plant species. Thus, the odours of flowers are distinct and are species-specific mixtures of compounds. Some flowers, such as arums (Arum) and carrion flowers (carrion flower), have qualitatively different odours that, to humans, are unpleasant. These flowers are pollinated by flies, moths, or beetles that are normally attracted to carrion, on which they lay their eggs.
Many flowers (flower) produce characteristic odours that attract pollinators. These odours are blends of volatile secondary compounds, including terpenes, derivatives of fatty acids (fatty acid), and aromatic phenolic compounds. Receptors on the insects' antennae respond to these compounds. However, while the response to odours of foliage is often innate, the response to odours of specific flowers is for the most part learned, since most insect pollinators are not specific to particular plants. In order to enhance the probability that an insect will visit plants of the same species, the insect must associate the presence of nectar with the odour of the flower that it last visited. This improves the foraging efficiency of the pollinator and increases the chance of cross pollination within a plant species. Thus, the odours of flowers are distinct and are species-specific mixtures of compounds. Some flowers, such as arums (Arum) and carrion flowers (carrion flower), have qualitatively different odours that, to humans, are unpleasant. These flowers are pollinated by flies, moths, or beetles that are normally attracted to carrion, on which they lay their eggs.Phagostimulation
All plants (plant) contain carbohydrates (carbohydrate), proteins, amino acids, and various lipids that are potential nutrients for animals. Some of these compounds can be tasted by animals and generally stimulate feeding (feeding behaviour) and thus are called phagostimulants (based on the Greek phagein, meaning “to eat”). In general, the taste of nutrient compounds is often essential for feeding and is used to adjust the amount eaten so that an organism maintains a suitable balance of nutrients. However, phagostimulants do not play a major role in determining the range of plants an animal will eat. Instead, the range of plants that animals feed on is determined to a very large extent by plant secondary compounds.
Although most secondary compounds are deterrent to the vast majority of species, there are some cases in which these compounds act as essential sign stimuli for an animal, indicating that it has the correct food. This is true for many insects that are oligophagous or monophagous on plants that contain characteristic chemicals. For example, plants in the cabbage family contain sulfur-containing compounds that act as sign stimuli for insects that habitually feed on only these plants. In the absence of the compounds, these insects will not feed. This is not because the compounds contain a chemical that provides some essential nutrient. In a few cases, it is known that the insects have receptor cells in the sensilla on their mouthparts or tarsi that are specifically sensitive to the sulfur-containing compounds, and this may be common in insects with chemically defined host-plant ranges. These same chemicals may be deterrents for insects that do not feed on these plants, as well as for insects that do feed on them.
Deterrents and repellents
 Many secondary compounds have low volatility and usually serve to reduce or completely inhibit feeding by most plant-feeding insects. Secondary compounds only affect an animal when it makes contact with the plant, which generally occurs when the animal bites into the plant. quinine and other alkaloids are examples of deterrents, as are glucosinolates and iridoid glycosides. In mammals these compounds are detected by the bitter taste receptors. Grasshoppers (grasshopper), butterflies, and moths also have cells that respond to a range of secondary compounds. The activity of these cells correlates with aversive behaviour, and they are usually called deterrent cells. Phytophagous beetles may not have these cells, and their host plant choice may depend on the indirect effect of secondary chemicals on the activity of sensory cells that signal acceptability.
Many secondary compounds have low volatility and usually serve to reduce or completely inhibit feeding by most plant-feeding insects. Secondary compounds only affect an animal when it makes contact with the plant, which generally occurs when the animal bites into the plant. quinine and other alkaloids are examples of deterrents, as are glucosinolates and iridoid glycosides. In mammals these compounds are detected by the bitter taste receptors. Grasshoppers (grasshopper), butterflies, and moths also have cells that respond to a range of secondary compounds. The activity of these cells correlates with aversive behaviour, and they are usually called deterrent cells. Phytophagous beetles may not have these cells, and their host plant choice may depend on the indirect effect of secondary chemicals on the activity of sensory cells that signal acceptability.Only a few instances are known in which a plant odour causes an insect to move away from the source. Linalool, a very common component of flower odours, is known to have a repellent effect on the carrot aphid, Cavariella. This may be a common phenomenon, but it has been little studied.
Feeding (feeding behaviour) decisions
Whether or not an animal eats a plant depends on phagostimulatory effects, mainly caused by nutrient compounds and sign stimulants, and on deterrent effects, caused by a variety of secondary chemicals. Polyphagous insects eat many plants that are unpalatable to oligophagous or monophagous species, even though all these insects may receive the same sensory information about plant chemistry. In the polyphagous species, deterrent compounds are less important in the interpretation of information by the central nervous than is true for selective feeders. An insect that is deprived of food or water tends to place less emphasis on deterrent signals and thus will eat a wide range of plants. The longer the period of deprivation, the greater the variety of plants that will be eaten.
Blood-feeding insects
 Insects that feed on vertebrate blood, such as mosquitoes (mosquito) and tsetse flies (tsetse fly), employ similar responses when locating and identifying their hosts. However, the chemical signals they use are different. Host odours cause takeoff, followed by upwind flight or, as in some tsetse flies, by visually oriented flight. lactic acid from human sweat is an important attractant for some mosquitoes, and octenol and acetone from cattle breath odours are also attractants. Blood-feeding insects have receptors on their antennae that are sensitive to these compounds. carbon dioxide is also an activator and attractant for several species of bloodsucking insects. Receptors for carbon dioxide have been demonstrated in not only insects that feed on blood but also several other kinds of insects, and these receptors are often on the maxillary palps (sensory structures associated with the mouthparts), rather than on the antennae. When an insect arrives at a potential host, chemicals on the host's skin likely influence the insect's behaviour, although the role of these chemicals is poorly understood. Once the insect starts to probe host tissues, adenosine diphosphate (ADP) and adenosine triphosphate (ATP) are released from red blood cells (erythrocyte) and may act as phagostimulants, causing the insects to gorge on blood. While many bloodsucking insects have receptors that are sensitive to ADP and ATP, others have receptors that are sensitive to different compounds.
Insects that feed on vertebrate blood, such as mosquitoes (mosquito) and tsetse flies (tsetse fly), employ similar responses when locating and identifying their hosts. However, the chemical signals they use are different. Host odours cause takeoff, followed by upwind flight or, as in some tsetse flies, by visually oriented flight. lactic acid from human sweat is an important attractant for some mosquitoes, and octenol and acetone from cattle breath odours are also attractants. Blood-feeding insects have receptors on their antennae that are sensitive to these compounds. carbon dioxide is also an activator and attractant for several species of bloodsucking insects. Receptors for carbon dioxide have been demonstrated in not only insects that feed on blood but also several other kinds of insects, and these receptors are often on the maxillary palps (sensory structures associated with the mouthparts), rather than on the antennae. When an insect arrives at a potential host, chemicals on the host's skin likely influence the insect's behaviour, although the role of these chemicals is poorly understood. Once the insect starts to probe host tissues, adenosine diphosphate (ADP) and adenosine triphosphate (ATP) are released from red blood cells (erythrocyte) and may act as phagostimulants, causing the insects to gorge on blood. While many bloodsucking insects have receptors that are sensitive to ADP and ATP, others have receptors that are sensitive to different compounds.Chemical defense
Defensive odours
 The best-known example of a vertebrate that uses odour for defense is the North American skunk. When threatened, skunks perform a visual warning. However, if this fails to deter a potential attacker, they produce an odorous spray from anal glands that are located on each side of the anus. The secretion contains several major and minor components that vary slightly among species. The compounds most offensive to humans are thiols (thiol). In addition, two of the three species whose secretions have been analyzed produce secretions containing acetates of thiols. These acetates slowly break down in air, giving rise to thiols and contributing to the persistence of the odour.
The best-known example of a vertebrate that uses odour for defense is the North American skunk. When threatened, skunks perform a visual warning. However, if this fails to deter a potential attacker, they produce an odorous spray from anal glands that are located on each side of the anus. The secretion contains several major and minor components that vary slightly among species. The compounds most offensive to humans are thiols (thiol). In addition, two of the three species whose secretions have been analyzed produce secretions containing acetates of thiols. These acetates slowly break down in air, giving rise to thiols and contributing to the persistence of the odour. Many insects also produce compounds that volatilize in contact with air and are effective repellents for potential predators (predation). The glands producing the compounds are distributed on various parts of the body. Many adult plant-sucking bugs (bug) have glands that open in front of the hind legs, and the products of these glands are released if the insect is touched, producing an unpleasant smell and giving rise to the common name “ stinkbug.” Many beetles also produce defensive compounds, and some stick insects (walkingstick) and a few grasshoppers produce compounds in a spray that can be ejected a distance of 40 cm (16 inches). Many different compounds are employed by different species to produce these defensive compounds. Often, strong odours are conspicuous in species that produce poisons, and the odour plays an important role in learning by predators, thus enhancing the protective effect of the poisons.
Many insects also produce compounds that volatilize in contact with air and are effective repellents for potential predators (predation). The glands producing the compounds are distributed on various parts of the body. Many adult plant-sucking bugs (bug) have glands that open in front of the hind legs, and the products of these glands are released if the insect is touched, producing an unpleasant smell and giving rise to the common name “ stinkbug.” Many beetles also produce defensive compounds, and some stick insects (walkingstick) and a few grasshoppers produce compounds in a spray that can be ejected a distance of 40 cm (16 inches). Many different compounds are employed by different species to produce these defensive compounds. Often, strong odours are conspicuous in species that produce poisons, and the odour plays an important role in learning by predators, thus enhancing the protective effect of the poisons.Defensive tastes
A wide variety of plants, marine animals, arthropods (arthropod), and vertebrates produce chemicals that are bitter to humans and distasteful to other vertebrate predators. Some of the animals acquire the chemicals from plants. Alkaloids are commonly used by all these groups, although a variety of other chemicals may be found. Iridoid glycosides, occurring in a number of plant families, are sequestered by checkerspot butterfly larvae and other insects that feed on the plants. These compounds are highly deterrent to ants and mammals. However, it should be noted that not all nonvolatile defensive chemicals are detected by the animals that encounter these plants and animals, and, if the chemicals are toxic, avoidance must depend on learning to associate illness with the flavour of the food that has been most recently eaten. In arthropods some defensive chemicals, such as quinones, phenols, acids (acid), and bases (base), have deterrent effects that stimulate vertebrate receptors involved in conveying sensations of burning or irritation to the brain via the trigeminal nerve.
Predator (predation) chemical cues and prey escape
Predator chemicals may be detected by some animals, although in most cases it is not known exactly how the chemicals are detected. For example, rabbits (rabbit) detect and move considerable distances away from feces of carnivorous mammals, and kangaroo rats (kangaroo rat) drum with their hind feet, probably as a warning to others, if they detect the odour of a predator. Salamanders (salamander) move away from substrates that are tainted by chemicals deposited by their snake predators, and they move out of waters that contain chemicals from fish predators. The anal sac secretions and urine of foxes (fox) have a range of volatile sulfur-containing compounds. The main compound studied is trimethyl triazoline, which causes freezing behaviour in rats. stoat anal sac chemicals cause alarm in snowshoe hares (hare).
 Among aquatic invertebrates (invertebrate), such as rotifers (rotifer), crustaceans (crustacean), and insects, there are many examples of sensitivity to predator chemicals that induce adaptive changes in behaviour or morphology. For example, in the water flea genus Daphnia, chemicals from predatory fish influence vertical migration patterns that reduce predation by fish. Chemicals from the predatory back swimmer bug in the genus Notonecta act as a predation cue by altering the response to light of Daphnia. This cue warns Daphnia of Notonecta's presence, giving it an opportunity to escape predation by the bugs. Barnacles (barnacle) on intertidal rocks normally produce a volcano-shaped armour. However, a specialist gastropod predator can breach this armour, unless the barnacle grows in a bent shape with the opening to the side. Young barnacles will develop either the volcano or the bent shape, depending on whether chemicals from the predator are absent or present in the water. How the chemicals induce these effects is unclear.
Among aquatic invertebrates (invertebrate), such as rotifers (rotifer), crustaceans (crustacean), and insects, there are many examples of sensitivity to predator chemicals that induce adaptive changes in behaviour or morphology. For example, in the water flea genus Daphnia, chemicals from predatory fish influence vertical migration patterns that reduce predation by fish. Chemicals from the predatory back swimmer bug in the genus Notonecta act as a predation cue by altering the response to light of Daphnia. This cue warns Daphnia of Notonecta's presence, giving it an opportunity to escape predation by the bugs. Barnacles (barnacle) on intertidal rocks normally produce a volcano-shaped armour. However, a specialist gastropod predator can breach this armour, unless the barnacle grows in a bent shape with the opening to the side. Young barnacles will develop either the volcano or the bent shape, depending on whether chemicals from the predator are absent or present in the water. How the chemicals induce these effects is unclear.Effects of experience
Changes in response to odour and taste may occur very rapidly. For example, a tendency to respond to an attractive food odour will decline if the food is out of reach, and many animals habituate to flavours (flavour) that are mildly distasteful on first encounter. On repeated encounters the flavours no longer elicit repellence or deterrence. However, most marked effects of chemosensory experience are of longer duration, lasting days, weeks, years, or in some cases a lifetime. Sometimes the chemoreceptive capacity is affected by experience, whereas other times the olfactory lobe structure or other integrative centres of the brain are affected.
Early experience
 Effects of early experience on odour and taste preferences have been studied in many animals, especially insects and mammals. For example, some caterpillars (caterpillar) that feed on only one of several equally acceptable host plant species will subsequently ignore or refuse the alternatives. In the larvae of the cabbage butterfly, the taste receptors develop a reduced sensitivity to mild deterrents in the experienced host and an enhanced sensitivity to the plant-specific phagostimulants.
Effects of early experience on odour and taste preferences have been studied in many animals, especially insects and mammals. For example, some caterpillars (caterpillar) that feed on only one of several equally acceptable host plant species will subsequently ignore or refuse the alternatives. In the larvae of the cabbage butterfly, the taste receptors develop a reduced sensitivity to mild deterrents in the experienced host and an enhanced sensitivity to the plant-specific phagostimulants.In several species of mammals, food preferences have been shown to be influenced in utero by the mother's diet. Chemicals from the maternal diet reach the fetus and cause long-lasting increases in the acceptance of foods containing the same chemicals. For example, young rabbits, whose mothers ate food containing juniper in the late stages of pregnancy, will, when subsequently weaned, exhibit a preference for juniper and even for the odour of juniper. This occurs regardless of whether, during weaning, they are fed by a different female who has not experienced juniper, indicating that the effect is not the result of a compound in the mother's milk. The effect results from an increase in the sensitivity to the odour of juniper in the young rabbit's olfactory epithelium. However, whether this arises through an increase in the frequency of a particular receptor type or an increase in sensitivity of existing receptors is not known. Comparable changes have been shown to occur in the preference of human babies for carrots (carrot), although the precise nature of the underlying mechanism has not been demonstrated.
Lactating females also can influence the later food preference of their offspring via chemicals ingested in the milk. This has been demonstrated in rats, ruminants (ruminant), and other animals; the food preferences of young livestock are conditioned before the young begin to eat solid food. In rats the process continues after weaning, with weanlings preferring to eat foods with odours accumulated on the mother's fur or in her breath. Such imprinting has been found in other contexts. For example, homing animals make use of odours experienced early in life to help them return to their natal place (see above Behaviour and chemoreception: Homing (chemoreception)).
associative learning
 A more plastic experiential change is seen in associations that develop at least to some extent in all animals with a central nervous system. An individual develops an association between sensory inputs (e.g., chemicals) and the important positive or negative effects experienced. Most studies have involved foraging and feeding behaviour. Parasitic (parasitism) wasps (wasp) learn to associate the presence of a host such as a caterpillar with the more prominent odours of the host's substrate (i.e., accumulated feces). Honeybees (honeybee) learn to associate particular floral odours with the presence of nectar rewards. Such learning often involves visual cues as well as chemical cues and increases foraging efficiency, minimizing time spent on fruitless searching when suitable resources are abundant. Among bees (bee), nest mates learn the floral odours picked up by foragers returning with food. The bees can use these odours to localize the food source in the field, after other signals have brought them to the general area.
A more plastic experiential change is seen in associations that develop at least to some extent in all animals with a central nervous system. An individual develops an association between sensory inputs (e.g., chemicals) and the important positive or negative effects experienced. Most studies have involved foraging and feeding behaviour. Parasitic (parasitism) wasps (wasp) learn to associate the presence of a host such as a caterpillar with the more prominent odours of the host's substrate (i.e., accumulated feces). Honeybees (honeybee) learn to associate particular floral odours with the presence of nectar rewards. Such learning often involves visual cues as well as chemical cues and increases foraging efficiency, minimizing time spent on fruitless searching when suitable resources are abundant. Among bees (bee), nest mates learn the floral odours picked up by foragers returning with food. The bees can use these odours to localize the food source in the field, after other signals have brought them to the general area.Specific nutritional learning of flavours (flavour) has also been demonstrated in various animal groups. For example, chemicals associated with complementary food sources, such as high protein and high carbohydrates (carbohydrate), can be learned. This enables locusts (locust), rats, cattle, and humans to choose the food type most needed at a particular time and thus, over a period of time, achieve a suitable balance between the two classes of nutrients. This ability is often combined with learned aversions to foods lacking specific nutrients. In the laboratory, slugs (slug) learn to reject a food lacking a single nontasted essential amino acid on the basis of the food flavour, and rats learn to reject a food lacking a single vitamin. Typically, the aversion to the flavour of the nutritionally inadequate food is accompanied by an increased attractiveness of novel flavours. Thus, aversion learning helps to increase the nutritional quality of the overall diet. In obtaining an ideal diet, generalist feeders are thought to use positive associative learning, aversion learning, and attraction to novel flavours. Over time, as conditions and needs change, new associations can develop.
How an animal (animal communication) determines that it has some specific nutritional deficiency is uncertain in most cases. In locusts the concentrations of some amino acids in the blood are of particular importance. In these insects the sensitivity of taste receptors to sugars and amino acids varies. If these insects are not ingesting enough protein, the responses of their receptors to amino acids are enhanced; if they are not ingesting enough carbohydrate, responses to sucrose are enhanced. If these nutrients are reliable indicators of carbohydrate and protein levels in food, variable sensitivity to them adds to the value of learned associations.
A danger for many omnivorous or polyphagous species is that potential food items may be poisonous. When an herbivore encounters a novel food that smells and tastes acceptable, the animal eats small amounts of it. If illness occurs, the illness is associated with the novel flavour or the flavour of the most recently eaten food, which is excluded from the diet thenceforth. This kind of aversion learning has been demonstrated in many species of insects, mollusks (mollusk), fish, mammals, and other animals that have brains (brain); it apparently does not occur in the phylum Cnidaria (cnidarian), since these organisms have only simple nerve nets. In mammals the senses of taste and smell play somewhat different roles in aversion learning. A novel odour alone is relatively ineffective and must be followed immediately by an aversive feedback to produce strong odour-aversion learning. However, strong aversions to flavours (taste and smell together) can be conditioned even when aversive feedback is delayed by up to 12 hours. When a weak odour is combined with a distinctive flavour and is followed by illness, the weak odour itself becomes a strong and long-term aversive stimulus.
Thus, the learned association between flavour and post-feeding distress occurs with respect to diets lacking important nutrients and foods that are poisonous. Apart from foraging and food selection, certain animals learn chemical cues associated with predators (predation), competitors, mates, and kin or social group, enabling them to behave in the most appropriate ways.
Influence of chemoreception in humans
Humans use a knowledge of the chemical senses to modify their own behaviour or physiology and to modify these properties in other animals.
Food (food processing) additives
 Probably the greatest knowledge of the influence of chemicals in human feeding (feeding behaviour) control relates to artificial sweeteners (sweetener). Sugars are phagostimulants; however, sugars and especially complex carbohydrates (e.g., starch), from which simple sugars may be derived in the oral cavity, are a source of fats, the primary storage form of carbohydrates. The accumulation of these fats can lead to obesity. As a result, humans have searched for substances that taste sweet but do not result in excessive fat storage. Such compounds are known to occur naturally in some plants and represent a range of structurally different chemical classes. For example, thaumatin is a sweet-tasting protein extracted from the tropical flowering plant Thaumatococcus daniellii, commonly called miracle fruit, or katempfe, and glycyrrhizin is a triterpene glycoside extracted from Glycyrrhiza glabra, or licorice.
Probably the greatest knowledge of the influence of chemicals in human feeding (feeding behaviour) control relates to artificial sweeteners (sweetener). Sugars are phagostimulants; however, sugars and especially complex carbohydrates (e.g., starch), from which simple sugars may be derived in the oral cavity, are a source of fats, the primary storage form of carbohydrates. The accumulation of these fats can lead to obesity. As a result, humans have searched for substances that taste sweet but do not result in excessive fat storage. Such compounds are known to occur naturally in some plants and represent a range of structurally different chemical classes. For example, thaumatin is a sweet-tasting protein extracted from the tropical flowering plant Thaumatococcus daniellii, commonly called miracle fruit, or katempfe, and glycyrrhizin is a triterpene glycoside extracted from Glycyrrhiza glabra, or licorice.In addition, many sweet-tasting compounds have been synthesized in the laboratory. In order to elicit the same response induced by a natural compound, the corresponding synthetic compound's molecular conformation must match the natural compound's receptor. More than 1,000 compounds have been synthesized following the discovery that l-aspartyl-l-phenylalanine methyl ester, which subsequently became known as aspartame, was found to taste sweet. In similar molecular quantities, some of the subsequent compounds taste much sweeter to humans than does sucrose. For example, aspartame is 200 times more potent than sucrose, whereas some modifications of aspartame are 50,000 times more potent. Artificial sweeteners that have been tested on Old World monkeys (monkey) have similar effects on humans, who are close relatives of these monkeys. However, these same sweeteners have variable effects on New World monkeys, which are more distantly related to humans, relative to Old World monkeys. These substances are presumed to stimulate the taste receptors using the same receptor proteins as sugars. Since a nerve carries electrical information in the form of action potentials (action potential), irrespective of the nature of the stimulating molecule, these substances are perceived as sweet.
Various food additives (food additive) are used by different societies. Chemicals are added to foods to influence the flavours (flavour) of foods, often stimulating appetite and digestive processes. monosodium glutamate (MSG) is commonly added to increase the umami, or meaty taste, of cooked dishes, and the flavour of many spices and herbs (spice and herb) increases production of saliva and other digestive juices or stimulates digestive processes. For example, the perception of peppermint increases saliva production, and the taste of cinnamon increases peristalsis in the gut. Individuals vary greatly in their olfactory sensitivity and in their chemosensory and cultural backgrounds, with the result that the use of additional flavours in foods is highly idiosyncratic. Nevertheless, flavour additives provide many people with pleasurable food experiences. In advanced commercial developments, use is made of the knowledge that minute amounts of key odour ingredients that typify favoured foods and beverages can be added to enhance the attractiveness of these products—for example, key components of the odour of freshly baked bread or of freshly roasted coffee.
Odour and culture
 The sense of smell has more important connections with the limbic system and hypothalamus in the brain than does hearing or vision. The close association between smell and the hypothalamus underlies the relationship of odour with emotion. Odour memory is long, and specific smells can vividly revive a past situation and emotion. Furthermore, pleasant or unpleasant odours may induce mild changes in mood, arousal, or cognition and may even reduce muscle tension. Many of these effects are at least partly a result of the circumstances of the use of odours. Odour compounds are used in a variety of human rituals, such as religious ceremonies and initiation ceremonies. For example, the burning of many fragrant woods and resins (resin) has been practiced for thousands of years in religious ceremonies, including in ancient Egyptian practices relating to Nefertem, the god of perfume, perfection, and beauty. A modern example of the use of odours in religious ceremonies is the burning of incense in the Roman Catholic (Roman Catholicism) mass. Many naturally derived fragrances are also used for aromatherapy, where, common to many human behaviours, the effects can often be explained more by expectations than by a direct effect on health.
The sense of smell has more important connections with the limbic system and hypothalamus in the brain than does hearing or vision. The close association between smell and the hypothalamus underlies the relationship of odour with emotion. Odour memory is long, and specific smells can vividly revive a past situation and emotion. Furthermore, pleasant or unpleasant odours may induce mild changes in mood, arousal, or cognition and may even reduce muscle tension. Many of these effects are at least partly a result of the circumstances of the use of odours. Odour compounds are used in a variety of human rituals, such as religious ceremonies and initiation ceremonies. For example, the burning of many fragrant woods and resins (resin) has been practiced for thousands of years in religious ceremonies, including in ancient Egyptian practices relating to Nefertem, the god of perfume, perfection, and beauty. A modern example of the use of odours in religious ceremonies is the burning of incense in the Roman Catholic (Roman Catholicism) mass. Many naturally derived fragrances are also used for aromatherapy, where, common to many human behaviours, the effects can often be explained more by expectations than by a direct effect on health.While the sense of smell is less important than vision or hearing in human interactions, odours do play an important role in influencing human behaviour. Every person has an individual odour, largely derived from apocrine secretions and epithelial flora, including bacteria and yeasts (yeast). Individuals may be recognized by their odour, and, a few days following birth, a baby is able to recognize its mother by her specific odour. Related people have more-similar odours, and the sexes have identifiably different odours that may play a role in sexual interactions. Some of the chemicals involved are used in perfumery.
Different cultures employ various means to reduce or enhance body odours, depending on the perceived unpleasantness or pleasantness of an odour. For example, deodorants may be used, particularly on the axillae and feet, that block secretions, kill bacteria, inhibit bacterial enzymes (enzyme), combine with unwanted odorants (e.g., isovaleric acid), or overpower odorants. Perfumes may employ particular body odour components that are considered attractive or may use floral, fruity, minty, or other fragrances, depending on the society. In certain cultures, perfumery is a multibillion-dollar industry.
In medicine, odours are employed in various ways. For example, in diagnostics, acetone on the breath is characteristic of diabetes mellitus, o-toluidine and aniline are characteristic of lung cancer, and sulfides (sulfide) are indicative of cirrhosis of the liver and dental disease. In the elderly, in people with damaged nasal epithelia from industrial pollutants, and in people with certain disease conditions, olfactory ability is decreased, and added food flavours can improve the experience of eating.
Human uses of chemoreception in other animals
Dogs (dog)
 Humans often employ the ability of dogs (dog) to learn specific odours in order to locate odour sources. Thus, dogs can be trained to help find missing or suspect persons by the odour associated with the person's clothing. Dogs can also be trained to locate drugs (drug) and are sometimes seen in this capacity sniffing at baggage as it is unloaded at airports.
Humans often employ the ability of dogs (dog) to learn specific odours in order to locate odour sources. Thus, dogs can be trained to help find missing or suspect persons by the odour associated with the person's clothing. Dogs can also be trained to locate drugs (drug) and are sometimes seen in this capacity sniffing at baggage as it is unloaded at airports.Monitoring and controlling pests
 The most widely used human applications of animal chemoreception involve attempts to control animals regarded as pests (pest). For example, sex-attractant pheromones (pheromone) of many moths (moth) of economic importance have been used to monitor and control moth populations. For monitoring, a synthetically produced pheromone is exposed in a trap, to which male moths are attracted and from which they are unable to escape. The lure of the pheromone is so strong that individual moths may be attracted even when the population is very low. By monitoring changes in the numbers caught, which are presumed to reflect the size of the population, the buildup of damaging populations can be predicted and potentially prevented. Population control using sex-attractant pheromones usually employs a different approach that is dependent on confusing the males. The object is to saturate an area with so much synthetic pheromone that the males are unable to locate calling females. To achieve this, the pheromone is dispersed over the area in small capsules or fibres of plastic, often dropped by aircraft. The capsules are designed so that the pheromone is released very slowly and persists in the environment for some weeks before a new application is required. This method has been used with some success against the corn earworm (or cotton bollworm) in the United States.
The most widely used human applications of animal chemoreception involve attempts to control animals regarded as pests (pest). For example, sex-attractant pheromones (pheromone) of many moths (moth) of economic importance have been used to monitor and control moth populations. For monitoring, a synthetically produced pheromone is exposed in a trap, to which male moths are attracted and from which they are unable to escape. The lure of the pheromone is so strong that individual moths may be attracted even when the population is very low. By monitoring changes in the numbers caught, which are presumed to reflect the size of the population, the buildup of damaging populations can be predicted and potentially prevented. Population control using sex-attractant pheromones usually employs a different approach that is dependent on confusing the males. The object is to saturate an area with so much synthetic pheromone that the males are unable to locate calling females. To achieve this, the pheromone is dispersed over the area in small capsules or fibres of plastic, often dropped by aircraft. The capsules are designed so that the pheromone is released very slowly and persists in the environment for some weeks before a new application is required. This method has been used with some success against the corn earworm (or cotton bollworm) in the United States.Altering pest behaviour
Chemicals are also used to inhibit feeding (feeding behaviour) by various animals on crops or ornamental plants. Some fungicides (fungicide) and other compounds have been shown experimentally to inhibit feeding by deer and granivorous (feeding on grain or seed) birds (bird), although it is not generally clear whether the effects are a consequence of distastefulness or olfactory repellence. A plant compound called azadirachtin has been widely used to inhibit feeding by herbivorous and granivorous insects. Azadirachtin is produced by the neem tree, which is native to northwestern India, although today it is widely grown in other parts of the world. The inhibitory effect of azadirachtin results from its taste. However, the efficacy of such methods is limited. For example, the compounds may be washed from foliage by rain, as plants grow, new growth is not protected; and, as an herbivore becomes increasingly hungry, it becomes less affected by the inhibitory effects. To overcome these obstacles, some compounds are injected into plants and some plants have been genetically engineered (genetic engineering) to produce deterrent substances (see genetically modified organism).
 The use of chemicals to repel nuisance insects is widely used in various human societies. For example, in Ethiopia, leaves of the pepper tree, Schinus molle, are used to repel houseflies (housefly), and two compounds from the leaves have been shown to produce the repellent effects. Citronella (citronella oil) extracted from plants is often used to repel mosquitoes (mosquito). In some countries, certain synthetic compounds may be used. For example, in the United States many people periodically use the compound commercially known as DEET to repel biting arthropods, especially mosquitoes and ticks. The active ingredient is N,N-diethyl-m-toluamide, which is mixed with other compounds to produce appropriate patterns of release in different circumstances.
The use of chemicals to repel nuisance insects is widely used in various human societies. For example, in Ethiopia, leaves of the pepper tree, Schinus molle, are used to repel houseflies (housefly), and two compounds from the leaves have been shown to produce the repellent effects. Citronella (citronella oil) extracted from plants is often used to repel mosquitoes (mosquito). In some countries, certain synthetic compounds may be used. For example, in the United States many people periodically use the compound commercially known as DEET to repel biting arthropods, especially mosquitoes and ticks. The active ingredient is N,N-diethyl-m-toluamide, which is mixed with other compounds to produce appropriate patterns of release in different circumstances.Additional Reading
Neurobiology of chemoreception
A clear, simple account of the molecular biology of olfaction and taste is Scott Brady et al., Basic Neurochemistry: Molecular, Cellular, and Medical Aspects, 7th ed. (2006). A book dealing specifically with the senses of taste and smell is Gary K. Beauchamp and Linda Bartoshuk (eds.), Tasting and Smelling (1997). The chemical senses in a range of organisms, including bacteria and humans, are summarized in Thomas E. Finger, Wayne L. Silver, and Diego Restrepo (eds.), The Neurobiology of Taste and Smell, 2nd ed. (2000).
Pheromones
A basic account of pheromone chemistry and associated behaviour is given in William C. Agosta, Chemical Communication: The Language of Pheromones (1992). An illustrated work providing information on pheromones and the importance of these compounds in chemical communication in animals is Tristram D. Wyatt, Pheromones and Animal Behaviour: Communication by Smell and Taste (2003).
Chemical interactions among plants and animals
The chemistry of plant and animal interactions is covered in Jeffrey B. Harborne, Introduction to Ecological Biochemistry, 4th ed. (1993). Aspects of semiochemistry are dealt with in Eric S. Albone and Stephen G. Shirley, Mammalian Semiochemistry: The Investigation of Chemical Signals Between Mammals (1984). A work covering the study of chemoreception in insects that includes information on the chemical interactions among plants and insects is Ring T. Cardé and William J. Bell, Chemical Ecology of Insects 2 (1995).
Perfumes and flavours
An account of the chemistry of perfumes that covers both natural and synthetic products is Charles Sell (ed.), The Chemistry of Fragrances: From Perfumer to Consumer (2006). A work providing insight on the perception of taste and flavour in humans is Andrew J. Taylor and Deborah D. Roberts (eds.), Flavor Perception (2004).
- Feast of Orthodoxy
- Feast of the Holy Family
- Feast of the Holy Innocents
- Feast of the Holy Trinity
- Feast of the Transfiguration
- feather
- featherbedding
- feather-duster worm
- Feathered Serpent
- feather moss
- feather star
- feathertail
- Feather, Victor, Baron Feather of the City of Bradford
- feather-winged beetle
- featherwork
- February
- February Revolution
- Febvre, Lucien Paul Victor
- feces
- Fechner, Gustav Theodor
- Feckenham, John de
- fedayee
- Fedchenko Glacier
- Fed Cup
- Fed Cup Table