dinosaur
extinct reptile
Introduction
 the common name given to a group of reptiles (reptile), often very large, that first appeared in the Late Triassic Period about 215 million years ago and thrived worldwide for some 150 million years. Most died out by the end of the Cretaceous Period, about 65 million years ago, but many lines of evidence now show that one lineage evolved into birds (bird) about 150 million years ago.
the common name given to a group of reptiles (reptile), often very large, that first appeared in the Late Triassic Period about 215 million years ago and thrived worldwide for some 150 million years. Most died out by the end of the Cretaceous Period, about 65 million years ago, but many lines of evidence now show that one lineage evolved into birds (bird) about 150 million years ago.The name dinosaur comes from the Greek words deinos (“terrible” or “fearfully great”) and sauros (“reptile” or “lizard”). The English anatomist Richard Owen (Owen, Sir Richard) proposed the formal term Dinosauria in 1842 to include three giant extinct animals ( Megalosaurus, Iguanodon, and Hylaeosaurus) represented by large fossilized bones that had been unearthed at several locations in southern England during the early part of the 19th century. Owen recognized that these reptiles were far different from other known reptiles of the present and the past for three reasons: they were large yet obviously terrestrial, unlike the aquatic ichthyosaurs (ichthyosaur) and plesiosaurs (plesiosaur) that were already known; they had five vertebrae in their hips, whereas most known reptiles have only two; and, rather than holding their limbs sprawled out to the side in the manner of lizards (lizard), dinosaurs held their limbs under the body in columnar fashion, like elephants (elephant) and other large mammals.
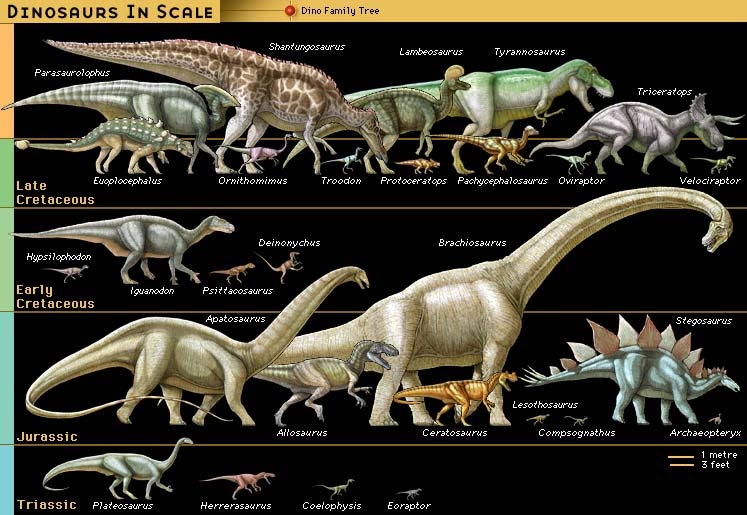
 Originally applied to just a handful of incomplete specimens, the category Dinosauria now encompasses more than 800 generic names and at least 1,000 species, with new names being added to the roster every year as the result of scientific explorations around the world. Not all of these names are valid taxa (taxon), however. A great many of them have been based on fragmentary or incomplete material that may actually have come from two or more different dinosaurs. In addition, bones have sometimes been misidentified as dinosaurian when they are not from dinosaurs at all. Nevertheless, dinosaurs are well documented by abundant fossil remains recovered from every continent on Earth, and the number of known dinosaurian taxa is estimated to be 10–25 percent of actual past diversity.
Originally applied to just a handful of incomplete specimens, the category Dinosauria now encompasses more than 800 generic names and at least 1,000 species, with new names being added to the roster every year as the result of scientific explorations around the world. Not all of these names are valid taxa (taxon), however. A great many of them have been based on fragmentary or incomplete material that may actually have come from two or more different dinosaurs. In addition, bones have sometimes been misidentified as dinosaurian when they are not from dinosaurs at all. Nevertheless, dinosaurs are well documented by abundant fossil remains recovered from every continent on Earth, and the number of known dinosaurian taxa is estimated to be 10–25 percent of actual past diversity. The extensive fossil record of genera and species is testimony that dinosaurs were diverse animals, with widely varying lifestyles and adaptations. Their remains are found in sedimentary rock layers (strata) dating to the Late Triassic Period (227 million to 206 million years ago). The abundance of their fossilized bones is substantive proof that dinosaurs were the dominant form of terrestrial animal life during the Mesozoic Era (248 million to 65 million years ago). It is likely that the known remains represent a very small fraction (probably less than 0.0001 percent) of all the individual dinosaurs that once lived.
The extensive fossil record of genera and species is testimony that dinosaurs were diverse animals, with widely varying lifestyles and adaptations. Their remains are found in sedimentary rock layers (strata) dating to the Late Triassic Period (227 million to 206 million years ago). The abundance of their fossilized bones is substantive proof that dinosaurs were the dominant form of terrestrial animal life during the Mesozoic Era (248 million to 65 million years ago). It is likely that the known remains represent a very small fraction (probably less than 0.0001 percent) of all the individual dinosaurs that once lived.The search for dinosaurs
The first finds
Before Richard Owen introduced the term Dinosauria in 1842, there was no concept of anything even like a dinosaur. Large fossilized bones quite probably had been observed long before that time, but there is little record—and no existing specimens—of such findings much before 1818. In any case, people could not have been expected to understand what dinosaurs were even if they found their remains. For example, some classical scholars now conclude that the Greco-Roman legends of griffins (griffin) from the 7th century BC were inspired by discoveries of protoceratopsian (Protoceratops) dinosaurs in the Altai (Altai Mountains) region of Mongolia. In 1676 Robert Plot of the University of Oxford (Oxford, University of) included, in a work of natural history, a drawing of what was apparently the knee-end of the thigh bone of a dinosaur, which he thought might have come from an elephant taken to Britain in Roman times. Fossil bones of what were undoubtedly dinosaurs were discovered in New Jersey in the late 1700s and were probably discussed at the meetings of the American Philosophical Society in Philadelphia. Soon thereafter, Lewis and Clark's expedition (Lewis and Clark Expedition) encountered dinosaur fossils in the western United States.
The earliest verifiable published record of dinosaur remains that still exists is a note in the 1820 American Journal of Science and Arts by Nathan Smith. The bones described had been found in 1818 by Solomon Ellsworth, Jr., while he was digging a well at his homestead in Windsor, Connecticut. At the time, the bones were thought to be human, but much later they were identified as Anchisaurus. Even earlier (1800), large birdlike footprints had been noticed on sandstone slabs in Massachusetts. Pliny Moody, who discovered these tracks, attributed them to “Noah's raven,” and Edward Hitchcock of Amherst College, who began collecting them in 1835, considered them to be those of some giant extinct bird. The tracks are now recognized as having been made by several different kinds of dinosaurs, and such tracks are still commonplace in the Connecticut River Valley today.
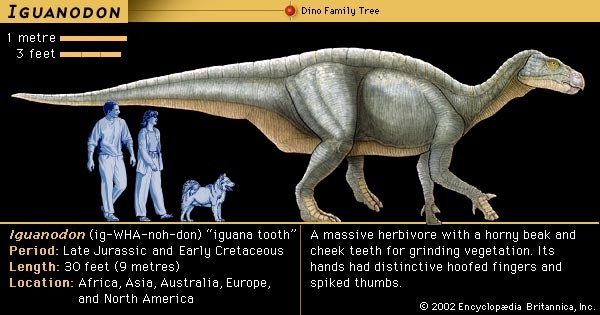
Better known are the finds in southern England during the early 1820s by William Buckland (Buckland, William) (a clergyman) and Gideon Mantell (Mantell, Gideon Algernon) (a physician), who described Megalosaurus and Iguanodon, respectively. In 1824 Buckland published a description of Megalosaurus, fossils of which consisted mainly of a lower jawbone with a few teeth. The following year Mantell published his “Notice on the Iguanodon, a Newly Discovered Fossil Reptile, from the Sandstone of Tilgate Forest, in Sussex,” on the basis of several teeth and some leg bones. Both men collected fossils as an avocation and are credited with the earliest published announcements in England of what later would be recognized as dinosaurs. In both cases their finds were too fragmentary to permit a clear image of either animal. In 1834 a partial skeleton was found near Brighton that corresponded with Mantell's fragments from Tilgate Forest. It became known as the Maidstone Iguanodon, after the village where it was discovered. The Maidstone skeleton provided the first glimpse of what these creatures might have looked like.
 Two years before the Maidstone Iguanodon came to light, a different kind of skeleton was found in the Weald (Weald, The) of southern England. It was described and named Hylaeosaurus by Mantell in 1832 and later proved to be one of the armoured dinosaurs. Other fossil bones began turning up in Europe: fragments described and named as Thecodontosaurus and Palaeosaurus by two English students, Henry Riley and Samuel Stutchbury, and the first of many skeletons named Plateosaurus by the naturalist Hermann von Meyer in 1837. Richard Owen (Owen, Sir Richard) identified two additional dinosaurs, albeit from fragmentary evidence: Cladeiodon, which was based on a single large tooth, and Cetiosaurus, which he named from an incomplete skeleton composed of very large bones. Having carefully studied most of these fossil specimens, Owen recognized that all of these bones represented a group of large reptiles that were unlike any living varieties. In a report to the British Association for the Advancement of Science in 1841, he described these animals, and the word Dinosauria was first published in the association's proceedings in 1842.
Two years before the Maidstone Iguanodon came to light, a different kind of skeleton was found in the Weald (Weald, The) of southern England. It was described and named Hylaeosaurus by Mantell in 1832 and later proved to be one of the armoured dinosaurs. Other fossil bones began turning up in Europe: fragments described and named as Thecodontosaurus and Palaeosaurus by two English students, Henry Riley and Samuel Stutchbury, and the first of many skeletons named Plateosaurus by the naturalist Hermann von Meyer in 1837. Richard Owen (Owen, Sir Richard) identified two additional dinosaurs, albeit from fragmentary evidence: Cladeiodon, which was based on a single large tooth, and Cetiosaurus, which he named from an incomplete skeleton composed of very large bones. Having carefully studied most of these fossil specimens, Owen recognized that all of these bones represented a group of large reptiles that were unlike any living varieties. In a report to the British Association for the Advancement of Science in 1841, he described these animals, and the word Dinosauria was first published in the association's proceedings in 1842.Reconstruction and classification (taxonomy)
 During the decades that followed Owen's announcement, many other kinds of dinosaurs were discovered and named in England and Europe: Massospondylus in 1854, Scelidosaurus in 1859, Bothriospondylus in 1875, and Omosaurus in 1877. Popular fascination with the giant reptiles grew, reaching a peak in the 1850s with the first attempts to reconstruct the three animals on which Owen based Dinosauria—Iguanodon, Megalosaurus, and Hylaeosaurus—for the first world exposition, the Great Exhibition of 1851 in London's Crystal Palace. A sculptor under Owen's direction (Waterhouse Hawkins) created life-size models of these two genera, and in 1854 they were displayed together with models of other extinct and living reptiles, such as plesiosaurs, ichthyosaurs, and crocodiles.
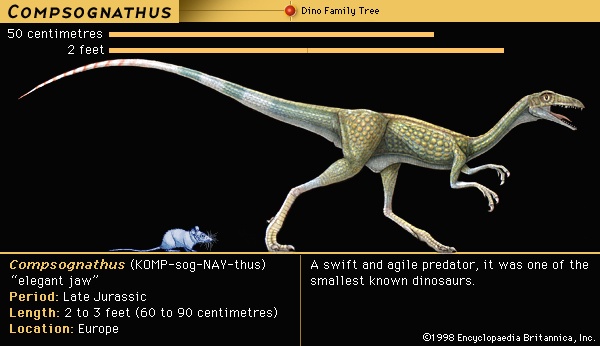
During the decades that followed Owen's announcement, many other kinds of dinosaurs were discovered and named in England and Europe: Massospondylus in 1854, Scelidosaurus in 1859, Bothriospondylus in 1875, and Omosaurus in 1877. Popular fascination with the giant reptiles grew, reaching a peak in the 1850s with the first attempts to reconstruct the three animals on which Owen based Dinosauria—Iguanodon, Megalosaurus, and Hylaeosaurus—for the first world exposition, the Great Exhibition of 1851 in London's Crystal Palace. A sculptor under Owen's direction (Waterhouse Hawkins) created life-size models of these two genera, and in 1854 they were displayed together with models of other extinct and living reptiles, such as plesiosaurs, ichthyosaurs, and crocodiles.By the 1850s it had become evident that the reptile fauna of the Mesozoic Era was far more diverse and complex than it is today. The first important attempt to establish an informative classification of the dinosaurs was made by the English biologist T.H. Huxley (Huxley, T.H.) as early as 1868. Because he observed that these animals had legs similar to birds as well as other birdlike features, he established a new order called Ornithoscelida. He divided the order into two suborders. Dinosauria was the first and included the iguanodonts, the large carnivores (or megalosaurids), and the armoured forms (including Scelidosaurus). Compsognatha was the second order, named for the very small birdlike carnivore Compsognathus.
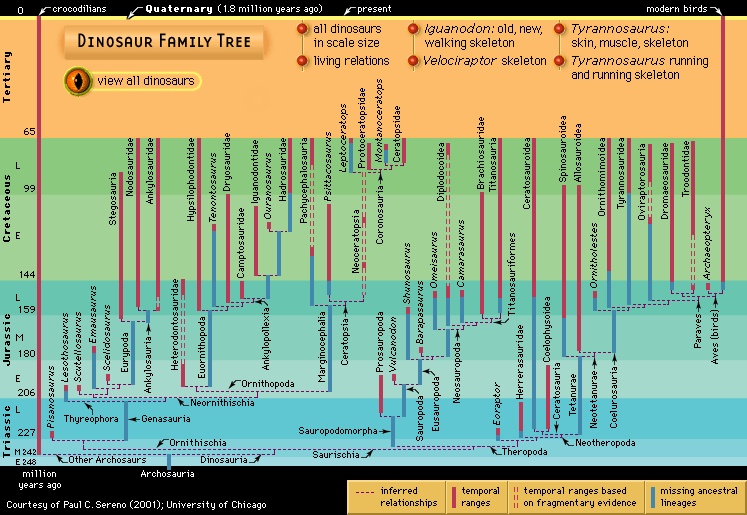
 Huxley's classification was replaced by a radically new scheme proposed in 1887 by his fellow Englishman H.G. Seeley, who noticed that all dinosaurs possessed one of two distinctive pelvic designs, one like that of birds and the other like that of reptiles. Accordingly, he divided the dinosaurs into the orders Ornithischia (ornithischian) (having a birdlike pelvis) and Saurischia (saurischian) (having a reptilian pelvis). Ornithischia included four suborders: Ornithopoda (ornithopod) (Iguanodon and similar herbivores), Stegosauria (plated forms), Ankylosauria (Hylaeosaurus and other armoured forms), and Ceratopsia (ceratopsian) (horned dinosaurs, just then being discovered in North America). Seeley's second order, the Saurischia (saurischian), included all the carnivorous dinosaurs, such as Megalosaurus and Compsognathus, as well as the giant herbivorous sauropods, including Cetiosaurus and several immense “brontosaur” types that were turning up in North America. In erecting Saurischia and Ornithischia, Seeley cast doubt on the idea that Dinosauria was a natural grouping of these animals. This uncertainty persisted for a century thereafter, but it is now understood that the two groups share unique features that indeed make the Dinosauria a natural group.
Huxley's classification was replaced by a radically new scheme proposed in 1887 by his fellow Englishman H.G. Seeley, who noticed that all dinosaurs possessed one of two distinctive pelvic designs, one like that of birds and the other like that of reptiles. Accordingly, he divided the dinosaurs into the orders Ornithischia (ornithischian) (having a birdlike pelvis) and Saurischia (saurischian) (having a reptilian pelvis). Ornithischia included four suborders: Ornithopoda (ornithopod) (Iguanodon and similar herbivores), Stegosauria (plated forms), Ankylosauria (Hylaeosaurus and other armoured forms), and Ceratopsia (ceratopsian) (horned dinosaurs, just then being discovered in North America). Seeley's second order, the Saurischia (saurischian), included all the carnivorous dinosaurs, such as Megalosaurus and Compsognathus, as well as the giant herbivorous sauropods, including Cetiosaurus and several immense “brontosaur” types that were turning up in North America. In erecting Saurischia and Ornithischia, Seeley cast doubt on the idea that Dinosauria was a natural grouping of these animals. This uncertainty persisted for a century thereafter, but it is now understood that the two groups share unique features that indeed make the Dinosauria a natural group.In 1878 a spectacular discovery was made in the town of Bernissart, Belgium, where several dozen complete articulated skeletons of Iguanodon were accidentally uncovered in a coal mine during the course of mining operations. Under the direction of the Royal Institute of Natural Science of Belgium, thousands of bones were retrieved and carefully restored over a period of many years. The first skeleton was placed on exhibit in 1883, and today the public can view an impressive herd of Iguanodon. The discovery of these multiple remains gave the first hint that at least some dinosaurs may have traveled in groups and showed clearly that some dinosaurs were bipedal (walking on two legs). The supervisor of this extraordinary project was Louis Dollo, a zoologist who was to spend most of his life studying Iguanodon, working out its structure, and speculating on its living habits.
American hunting expeditions
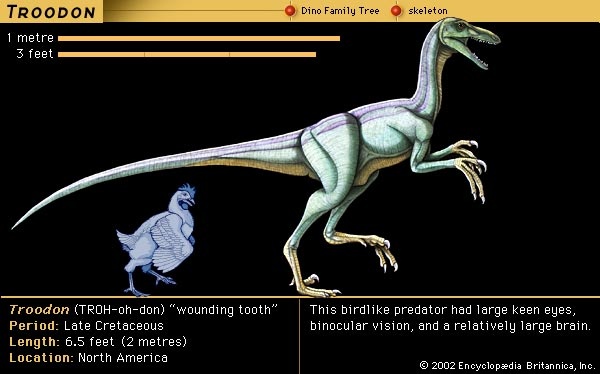 England and Europe produced most of the early discoveries and students of dinosaurs, but North America soon began to contribute a large share of both. One leading student of fossils was Joseph Leidy (Leidy, Joseph) of the Academy of Natural Sciences in Philadelphia, who named some of the earliest dinosaurs found in America, including Palaeoscincus, Trachodon, Troodon, and Deinodon. Unfortunately, some names given by Leidy are no longer used, because they were based on such fragmentary and undiagnostic material. Leidy is perhaps best known for his study and description of the first dinosaur skeleton to be recognized in North America, that of a duckbill, or hadrosaur, found at Haddonfield, New Jersey, in 1858, which he named Hadrosaurus foulkii. Leidy's inference that this animal was probably amphibious influenced views of dinosaur life for the next century.
England and Europe produced most of the early discoveries and students of dinosaurs, but North America soon began to contribute a large share of both. One leading student of fossils was Joseph Leidy (Leidy, Joseph) of the Academy of Natural Sciences in Philadelphia, who named some of the earliest dinosaurs found in America, including Palaeoscincus, Trachodon, Troodon, and Deinodon. Unfortunately, some names given by Leidy are no longer used, because they were based on such fragmentary and undiagnostic material. Leidy is perhaps best known for his study and description of the first dinosaur skeleton to be recognized in North America, that of a duckbill, or hadrosaur, found at Haddonfield, New Jersey, in 1858, which he named Hadrosaurus foulkii. Leidy's inference that this animal was probably amphibious influenced views of dinosaur life for the next century.Two Americans whose work during the second half of the 19th century had worldwide impact on the science of paleontology in general, and the growing knowledge of dinosaurs in particular, were O.C. Marsh (Marsh, Othniel Charles) of Yale (Yale University) College and E.D. Cope (Cope, Edward Drinker) of Haverford College, the University of Pennsylvania (Pennsylvania, University of), and the Academy of Natural Sciences in Philadelphia. All previous dinosaur remains had been discovered by accident in well-populated regions with temperate, moist climates, but Cope and Marsh astutely focused their attention on the wide arid expanses of bare exposed rock in western North America. In their intense quest to find and name new dinosaurs, these scientific pioneers became fierce and unfriendly rivals.
Marsh's field parties explored widely, exploiting dozens of now famous areas, among them Yale's sites at Morrison and Canon City, Colorado, and, most important, Como Bluff in southeastern Wyoming. The discovery of Como Bluff in 1877 was a momentous event in the history of paleontology that generated a burst of exploration and study as well as widespread public enthusiasm for dinosaurs. Como Bluff brought to light one of the greatest assemblages of dinosaurs, both small and gigantic, ever found. For decades the site went on producing the first known specimens of Late Jurassic Period (159 million to 144 million years ago) dinosaurs such as Stegosaurus, Camptosaurus, Camarasaurus, Laosaurus, Coelurus, and others. From the Morrison site came the original specimens of Allosaurus, Diplodocus, Atlantosaurus, and Brontosaurus (later renamed Apatosaurus). Canon City provided bones of a host of dinosaurs, including Stegosaurus, Brachiosaurus (brachiosaur), Allosaurus, and Camptosaurus.
Another major historic site was the Lance Creek (Lance Formation) area of northeastern Wyoming, where J.B. Hatcher discovered and collected dozens of Late Cretaceous (Cretaceous Period) horned dinosaur remains for Marsh and for Yale College, among them the first specimens of Triceratops and Torosaurus. Marsh was aided in his work at these and other localities by the skills and efforts of many other collaborators like Hatcher—William Reed, Benjamin Mudge, Arthur Lakes, William Phelps, and Samuel Wendell Williston, to name a few. Marsh's specimens now form the core of the Mesozoic collections at the National Museum of Natural History of the Smithsonian Institution and the Peabody Museum of Natural History at Yale University.
 Cope's dinosaur explorations ranged as far as, or farther than, Marsh's, and his interests encompassed a wider variety of fossils. Owing to a number of circumstances, however, Cope's dinosaur discoveries were fewer and his collections far less complete than those of Marsh. Perhaps his most notable achievement was finding and proposing the names for Coelophysis and Monoclonius. Cope's dinosaur explorations began in the eastern badlands (badland) of Montana, where he discovered Monoclonius in the Judith River Formation of the Cretaceous Period (99 million to 65 million years ago). Accompanying him there was a talented young assistant, Charles H. Sternberg. Later Sternberg and his three sons went on to recover countless dinosaur skeletons from the Oldman and Edmonton formations of the Late Cretaceous along the Red Deer River of Alberta, Canada.
Cope's dinosaur explorations ranged as far as, or farther than, Marsh's, and his interests encompassed a wider variety of fossils. Owing to a number of circumstances, however, Cope's dinosaur discoveries were fewer and his collections far less complete than those of Marsh. Perhaps his most notable achievement was finding and proposing the names for Coelophysis and Monoclonius. Cope's dinosaur explorations began in the eastern badlands (badland) of Montana, where he discovered Monoclonius in the Judith River Formation of the Cretaceous Period (99 million to 65 million years ago). Accompanying him there was a talented young assistant, Charles H. Sternberg. Later Sternberg and his three sons went on to recover countless dinosaur skeletons from the Oldman and Edmonton formations of the Late Cretaceous along the Red Deer River of Alberta, Canada.Dinosaur ancestors
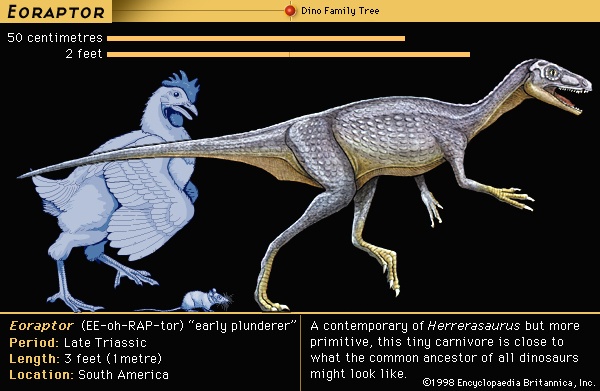 During the early decades of dinosaur discoveries, little thought was given to their evolutionary ancestry. Not only were the few specimens known unlike any living animal, but they were so different from any other reptiles that it was difficult to discern much about their relationships. Early on it was recognized that, as a group, dinosaurs appear to be most closely allied to crocodilians (crocodile), though T.H. Huxley had proposed in the 1860s that dinosaurs and birds must have had a very close common ancestor in the distant past. Three anatomic features—socketed teeth, a skull with two large holes (diapsid), and another hole in the lower jaw—are present in both crocodiles and dinosaurs. The earliest crocodilians occurred nearly simultaneously with the first known dinosaurs, so neither could have given rise to the other. It was long thought that the most likely ancestry of dinosaurs could be found within a poorly understood group of Triassic reptiles termed thecodontians (thecodontian) (“socket-toothed reptiles”). Today it is recognized that “thecodontian” is simply a name for the basal, or most primitive, members of the archosaurs (archosaur) (“ruling reptiles”), a group that is distinguished by the three anatomic features mentioned above and that includes dinosaurs, pterosaurs (pterosaur) (flying reptiles), crocodiles, and their extinct relatives. An early candidate for the ancestor of dinosaurs was a small basal archosaur from the Early Triassic Period (248 million to 242 million years ago) of South Africa called Euparkeria. New discoveries suggest creatures that are even more dinosaur-like from the Middle Triassic (242 million to 227 million years ago) and from an early portion of the Late Triassic (227 million to 206 million years ago) of South America; these include Lagerpeton, Lagosuchus, Pseudolagosuchus, and Lewisuchus. Other South American forms such as Eoraptor and Herrerasaurus are particularly dinosaurian in appearance and are sometimes considered dinosaurs.
During the early decades of dinosaur discoveries, little thought was given to their evolutionary ancestry. Not only were the few specimens known unlike any living animal, but they were so different from any other reptiles that it was difficult to discern much about their relationships. Early on it was recognized that, as a group, dinosaurs appear to be most closely allied to crocodilians (crocodile), though T.H. Huxley had proposed in the 1860s that dinosaurs and birds must have had a very close common ancestor in the distant past. Three anatomic features—socketed teeth, a skull with two large holes (diapsid), and another hole in the lower jaw—are present in both crocodiles and dinosaurs. The earliest crocodilians occurred nearly simultaneously with the first known dinosaurs, so neither could have given rise to the other. It was long thought that the most likely ancestry of dinosaurs could be found within a poorly understood group of Triassic reptiles termed thecodontians (thecodontian) (“socket-toothed reptiles”). Today it is recognized that “thecodontian” is simply a name for the basal, or most primitive, members of the archosaurs (archosaur) (“ruling reptiles”), a group that is distinguished by the three anatomic features mentioned above and that includes dinosaurs, pterosaurs (pterosaur) (flying reptiles), crocodiles, and their extinct relatives. An early candidate for the ancestor of dinosaurs was a small basal archosaur from the Early Triassic Period (248 million to 242 million years ago) of South Africa called Euparkeria. New discoveries suggest creatures that are even more dinosaur-like from the Middle Triassic (242 million to 227 million years ago) and from an early portion of the Late Triassic (227 million to 206 million years ago) of South America; these include Lagerpeton, Lagosuchus, Pseudolagosuchus, and Lewisuchus. Other South American forms such as Eoraptor and Herrerasaurus are particularly dinosaurian in appearance and are sometimes considered dinosaurs.The earliest appearance of “true dinosaurs” is almost impossible to pinpoint, since it can never be known with certainty whether the very first (or last) specimen of any kind of organism has been found. The succession of deposits containing fossils is discontinuous and contains many gaps; even within these deposits, the fossil record of dinosaurs and other creatures contained within is far from complete. Further complicating matters is that evolution from ancestral to descendant form is usually a stepwise process. Consequently, as more and more gaps are filled between the first dinosaurs and other archosaurs, the number of features distinguishing them becomes smaller and smaller. Currently, paleontologists define dinosaurs as Triceratops (representing Ornithischia), birds (the most recent representatives of the Saurischia (saurischian)), and all the descendants of their most recent common ancestor. That common ancestor apparently had a suite of features not present in other dinosaur relatives, including the loss of the prefrontal bone above the eye, a long deltopectoral crest on the humerus, three or fewer joints on the fourth finger of the hand, three or more hip vertebrae, a fully open hip socket, and a cnemial crest on the shin bone ( tibia). These features were passed on and modified in the descendants of the first dinosaurs. Compared with most of their contemporaries, dinosaurs had an improved stance and posture with a resulting improved gait and, in several independent lineages, an overall increase in size. They also were more efficient at gathering food and processing it and apparently had higher metabolic rates and cardiovascular nourishment. All these trends, individually or in concert, probably contributed to the collective success of dinosaurs, which resulted in their dominance among the terrestrial animals of the Mesozoic.
Modern studies
 During the first century or more of dinosaur awareness, workers in the field more or less concentrated on the search for new specimens and new types. Their discoveries then required detailed description and analysis, followed by comparisons with other known dinosaurs in order to classify the new finds and develop hypotheses about evolutionary relationships. These pursuits continue, but newer methods of exploration and analysis have been adopted. Emphasis has shifted from purely descriptive procedures to analyses of relationships by using the methods of cladistics, which dispenses with the traditional taxonomic (taxonomy) hierarchy in favour of “phylogenetic trees (phylogenetic tree)” that are more explicit about evolutionary relationships. Phylogenetic analyses also help us to understand how certain features evolved in groups of dinosaurs and give us insight into their possible functions. For example, in the evolution of horned dinosaurs (ceratopsians (ceratopsian)), it can be seen that the beak evolved first, followed by the frill, and finally the nose and eye horns, which were differently developed in different groups. The hypothesis that the frill was widely used in defense by ceratopsians such as Protoceratops can thus be tested phylogenetically. On this basis, the idea is now generally rejected because the frill was basically just an open rim of bone in nearly all ceratopsians except Triceratops, which is often pictured charging like a rhinoceros.
During the first century or more of dinosaur awareness, workers in the field more or less concentrated on the search for new specimens and new types. Their discoveries then required detailed description and analysis, followed by comparisons with other known dinosaurs in order to classify the new finds and develop hypotheses about evolutionary relationships. These pursuits continue, but newer methods of exploration and analysis have been adopted. Emphasis has shifted from purely descriptive procedures to analyses of relationships by using the methods of cladistics, which dispenses with the traditional taxonomic (taxonomy) hierarchy in favour of “phylogenetic trees (phylogenetic tree)” that are more explicit about evolutionary relationships. Phylogenetic analyses also help us to understand how certain features evolved in groups of dinosaurs and give us insight into their possible functions. For example, in the evolution of horned dinosaurs (ceratopsians (ceratopsian)), it can be seen that the beak evolved first, followed by the frill, and finally the nose and eye horns, which were differently developed in different groups. The hypothesis that the frill was widely used in defense by ceratopsians such as Protoceratops can thus be tested phylogenetically. On this basis, the idea is now generally rejected because the frill was basically just an open rim of bone in nearly all ceratopsians except Triceratops, which is often pictured charging like a rhinoceros.Functional anatomic studies extensively use analogous traits (analogy) of present-day animals that, along with both mechanical and theoretical models, make it possible to visualize certain aspects of extinct animals. For example, estimates of normal walking (locomotion) and maximum running speeds can be calculated on the basis of the analysis of trackways, which can then be combined with biomechanical (biomechanics) examination of the legs and joints and reconstruction of limb musculature. Similar methods have been applied to jaw mechanisms and tooth wear patterns to obtain a better understanding of feeding habits and capabilities.
The soft parts of dinosaurs are only imperfectly known. Original colours and patterns cannot be known, but skin textures have occasionally been preserved. Most show a knobby or pebbly surface rather than a scaly texture as in most living reptiles. Impressions of internal organs are rarely preserved, but, increasingly, records of filaments and even feathers have been found on some dinosaurs. Gastroliths (“stomach stones”) used for processing food in the gizzard have been recovered from a variety of dinosaurs.
extinction
A misconception commonly portrayed in popular books and media is that all the dinosaurs died out at the same time—and apparently quite suddenly—at the end of the Cretaceous Period 65 million years ago. This is not entirely correct, and not only because birds are a living branch of dinosaurian lineage. The best records, which are almost exclusively from North America, show that dinosaurs were already in decline during the latest portion of the Cretaceous. The causes of this decline, as well as the fortunes of other groups at the time, are complex and difficult to attribute to a single source. In order to understand extinction, it is necessary to understand the basic fossil record of dinosaurs.
Faunal changes
During the 160 million years or so of the Mesozoic Era (248 million to 65 million years ago) from which dinosaurs are known, there were constant changes in dinosaur communities. Different species evolved rapidly and were quickly replaced by others throughout the Mesozoic; it is rare that any particular type of dinosaur survived from one geologic formation into the next. The fossil evidence shows a moderately rich fauna of plateosaurs (Plateosaurus) and other prosauropods, primitive ornithopods (ornithopod), and theropods (theropod) during the Late Triassic Period (227 million to 206 million years ago). Most of these kinds of dinosaurs are also represented in strata of the Early Jurassic (Jurassic Period) Period (206 million to 180 million years ago), but following a poorly known Middle Jurassic, the fauna of the Late Jurassic (159 million to 144 million years ago) was very different. By this time sauropods (sauropod), more advanced ornithopods, stegosaurs, and a variety of theropods predominated. The Early Cretaceous (144 million to 99 million years ago) then contained a few sauropods (albeit they were all new forms), a few stegosaurian (dinosaur) holdovers, new kinds of theropods and ornithopods, and some of the first well-known ankylosaurs (dinosaur). By the Late Cretaceous (99 million to 65 million years ago), sauropods, which had disappeared from the northern continents through most of the Early and mid-Cretaceous, had reinvaded the northern continents from the south, and advanced ornithopods (duckbills) had become the dominant browsers. A variety of new theropods of all sizes were widespread; stegosaurs no longer existed; and the ankylosaurs were represented by a collection of new forms that were prominent in the North America and Asia. New groups of dinosaurs, the pachycephalosaurs (dinosaur) and ceratopsians (ceratopsian), had appeared in Asia and had successfully colonized North America. The overall picture is thus quite clear: throughout Mesozoic time there was a continuous dying out and renewal of dinosaurian life.
It is important to note that extinction is a normal, universal occurrence. Mass extinctions often come to mind when the term extinction is mentioned, but the normal background extinctions that occur throughout geologic time probably account for most losses of biodiversity. Just as new species constantly split from existing ones, existing species are constantly becoming extinct. The speciation rate of a group must, on balance, exceed the extinction rate in the long run, or that group will become extinct. The history of animal and plant life is replete with successions as early forms are replaced by new and often more advanced forms. In most instances the layered (stratigraphic) nature of the fossil record gives too little information to show whether the old forms were actually displaced by the new successors (from the effects of competition, predation, or other ecological processes) or if the new kinds simply expanded into the declining population's ecological niches.
Because the fossil record is episodic rather than continuous, it is very useful for asking many kinds of questions, but it is not possible to say precisely how long most dinosaur species or genera actually existed. Moreover, because the knowledge of the various dinosaur groups is somewhat incomplete, the duration of any particular dinosaur can be gauged only approximately—usually by stratigraphic boundaries and presumed “first” and “last” occurrences. The latter often coincide with geologic age boundaries; in fact, the absence of particular life-forms has historically defined most geologic boundaries ever since the geologic record was first compiled and analyzed in the late 18th century. The “moments” of apparently high extinction levels among dinosaurs were near the ends of two stages of the Triassic (about 221 million and 210 million years ago), perhaps at the end of the Jurassic (144 million years ago), and of course at the end of the Cretaceous (65 million years ago). Undoubtedly, there were lesser extinction peaks at other times in between, but there are poor terrestrial records for most of the world in the Middle Triassic, Middle Jurassic, and mid-Cretaceous.
The K–T boundary event
It was not only the dinosaurs that disappeared 65 million years ago at the Cretaceous–Tertiary, or K–T, boundary. Many other organisms became extinct or were greatly reduced in abundance and diversity, and the extinctions were quite different between, and even among, marine and terrestrial organisms. Land plants did not respond in the same way as land animals, and not all marine organisms showed the same patterns of extinction. Some groups died out well before the K–T boundary, including flying reptiles (pterosaurs (pterosaur)) and sea reptiles (plesiosaurs (plesiosaur), mosasaurs (mosasaur), and ichthyosaurs (ichthyosaur)). Strangely, turtles (turtle), crocodilians (crocodile), lizards (lizard), and snakes (snake) were either not affected or affected only slightly. Effects on amphibians and mammals were mild. These patterns seem odd, considering how environmentally sensitive and habitat-restricted many of these groups are today. Many marine groups—such as the molluscan ammonites (ammonoid), the belemnites (belemnoid), and certain bivalves (bivalve)—were decimated. Other greatly affected groups were the moss animals (moss animal) (phylum Bryozoa), the crinoids (crinoid), and a number of planktonic life-forms such as foraminifera (foraminiferan), radiolarians (radiolarian), coccolithophores, and diatoms (diatom).
Whatever factors caused it, there was undeniably a major, worldwide biotic change near the end of the Cretaceous. But the extermination of the dinosaurs is the best-known change by far, and it has been a puzzle to paleontologists, geologists, and biologists for two centuries. Many hypotheses have been offered over the years to explain dinosaur extinction, but only a few have received serious consideration. Proposed causes have included everything from disease to heat waves and resulting sterility, freezing cold spells, the rise of egg-eating mammals, and X rays from a nearby exploding supernova. Since the early 1980s, attention has focused on the so-called asteroid theory put forward by the American geologist Walter Alvarez, his father, physicist Luis Alvarez (Alvarez, Luis W.), and their coworkers. This theory is consistent with the timing and magnitude of some extinctions, especially in the oceans, but it does not fully explain the patterns on land and does not eliminate the possibility that other factors were at work on land as well as in the seas.
One important question is whether the extinctions were simultaneous and instantaneous or whether they were nonsynchronous and spread over a long time. The precision with which geologic time can be measured leaves much to be desired no matter what means are used (radiometric, paleomagnetic, or the more traditional measuring of fossil content of stratigraphic layers). Only rarely does an “instantaneous” event leave a worldwide—or even regional—signature in the geologic record in the way that a volcanic eruption does locally. Attempts to pinpoint the K–T boundary event, even by using the best radiometric dating techniques, result in a margin of error on the order of 50,000 years. Consequently, the actual time involved in this, or any of the preceding or subsequent extinctions, has remained undetermined.
The asteroid theory
The discovery of an abnormally high concentration of the rare metal iridium at, or very close to, the K–T boundary provides what has been recognized as one of those rare instantaneous geologic time markers that seem to be worldwide. This iridium anomaly, or spike, was first found by Walter Alvarez in the Cretaceous–Tertiary stratigraphic sequence at Gubbio, Italy, in the 1970s. The spike has subsequently been detected at hundreds of localities in Denmark and elsewhere, both in rock outcrops on land and in core samples drilled from ocean floors. Iridium normally is a rare substance in rocks of the Earth's crust (about 0.3 part per billion). At Gubbio the iridium concentration is more than 20 times greater (6.3 parts per billion), and it exceeds this concentration at other sites.
Because the levels of iridium are higher in meteorites than on the Earth, the Gubbio anomaly is thought to have an extraterrestrial explanation. If this is true, such extraterrestrial signatures will have a growing influence on the precision with which geologic time boundaries can be specified. The level of iridium in meteorites (meteorite) has been accepted as representing the average level throughout the solar system and, by extension, the universe. Accordingly, the iridium concentration at the K–T boundary is widely attributed to a collision between the Earth and a huge meteor (meteor and meteoroid) or asteroid. The size of the object is estimated at about 10 km (6.2 miles) in diameter and one quadrillion metric tons in weight; the velocity at the time of impact is reckoned to have been several hundreds of thousands of kilometres per hour. The crater resulting from such a collision would be some 100 km or more in diameter. Such an impact site (called an astrobleme), known as the Chicxulub crater, may have been identified in the Yucatán Peninsula.
The asteroid theory is widely accepted as the most probable explanation of the K–T iridium anomaly, but it does not appear to account for all the paleontological data. An impact explosion of this kind would have ejected an enormous volume of terrestrial and asteroid material into the atmosphere, producing a cloud of dust and solid particles that would have encircled the Earth and blocked out sunlight for many months, possibly years. The loss of sunlight could have eliminated photosynthesis and resulted in the death of plants and the subsequent extinction of herbivores, their predators, and scavengers.
The K–T mass extinctions, however, do not seem to be fully explained by this hypothesis. The stratigraphic record is most complete for extinctions of marine life—foraminifera, ammonites, coccolithophores, and the like. These apparently died out suddenly and simultaneously, and their extinction accords best with the asteroid theory. The fossil evidence of land dwellers, however, suggests a gradual rather than a sudden decline in dinosaurian diversity (and possibly abundance). Alterations in terrestrial life seem to be best accounted for by environmental factors, such as the consequences of seafloor spreading and continental drift, resulting in continental fragmentation, climatic deterioration, increased seasonality, and perhaps changes in the distributions and compositions of terrestrial communities. But one phenomenon does not preclude another. It is entirely possible that a culmination of ordinary biological changes and some catastrophic events, including increased volcanic activity, took place around the end of the Cretaceous.
Dinosaur descendants
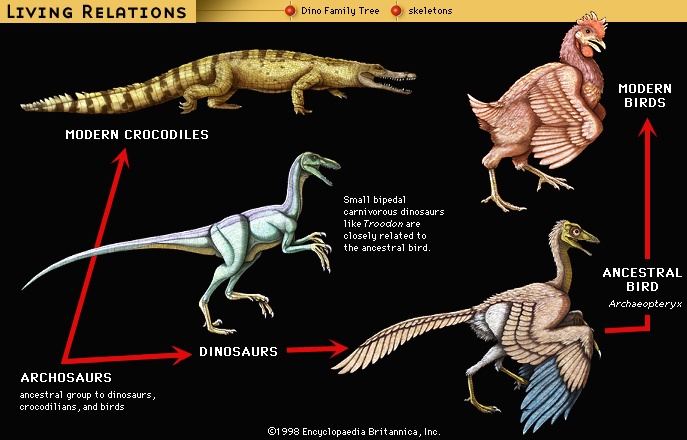 Contrary to the commonly held belief that the dinosaurs left no descendants, the rare (seven) specimens of Archaeopteryx (the earliest bird known) provide compelling evidence that birds (bird) (class Aves) evolved from small theropod dinosaurs. Following the principles of genealogy that are applied to humans as much as to other organisms, organisms are classified at a higher level within the groups from which they evolved. Archaeopteryx is therefore classified as both a dinosaur and a bird, just as humans are both primates and mammals.
Contrary to the commonly held belief that the dinosaurs left no descendants, the rare (seven) specimens of Archaeopteryx (the earliest bird known) provide compelling evidence that birds (bird) (class Aves) evolved from small theropod dinosaurs. Following the principles of genealogy that are applied to humans as much as to other organisms, organisms are classified at a higher level within the groups from which they evolved. Archaeopteryx is therefore classified as both a dinosaur and a bird, just as humans are both primates and mammals.The specimens of Archaeopteryx contain particular anatomic features that also are exclusively present in certain theropods (theropod) ( Oviraptor, Velociraptor, Deinonychus, and Troodon, among others). These animals share long arms and hands, a somewhat shorter, stiffened tail, a similar pelvis, and an unusual wrist joint in which the hand is allowed to flex sideways instead of up and down. This wrist motion is virtually identical to the motion used by birds (and bats (bat)) in flight, though in these small dinosaurs its initial primary function was probably in catching prey.
Beginning in the 1990s, several specimens of small theropod dinosaurs from the Early Cretaceous of Liaoning province, China, were unearthed. These fossils are remarkably well preserved, and because they include impressions of featherlike (feather), filamentous structures that covered the body, they have shed much light on the relationship between birds and Mesozoic dinosaurs. Such structures are now known in a compsognathid (Sinosauropteryx), a therizinosaurid (Beipiaosaurus), a dromaeosaur (Sinornithosaurus), and an alvarezsaurid (Shuvuuia). The filamentous structures on the skin of Sinosauropteryx are similar to the barbs of feathers, which suggests that feathers evolved from a much simpler structure that probably functioned as an insulator. True feathers (feather) of several types, including contour and body feathers, have been found in the 125-million-year-old feathered oviraptorid Caudipteryx and the apparently related Protarchaeopteryx. Because these animals were not birds and did not fly, it is now evident that true feathers neither evolved first in birds nor developed for the purpose of flight. Instead, feathers may have evolved for insulation, display, camouflage, species recognition, or some combination of these functions and only later became adapted for flight. In the case of Caudipteryx, for example, it has been established that these animals not only sat on nests but probably protected the eggs with their feathers.
 Until comparatively recent times, the two groups of birds from Cretaceous time that received the most attention because of their strange form were the divers, such as , (Hesperornis) and the strong-winged , (Ichthyornis) a more ternlike (tern) form. Because they were the first well-known Cretaceous birds, having been described by American paleontologist O.C. Marsh (Marsh, Othniel Charles) in 1880, they were thought to represent typical Cretaceous birds. Recent discoveries, however, have changed this view. For example, members of one Early Cretaceous bird group, the Confuciusornithidae, showed very little advancement compared with Archaeopteryx and the Enantiornithes (a major group of birds widely distributed around the world through most of the Cretaceous Period). Because representatives of living bird groups have long been known among the fossil species from the Paleocene (Paleocene Epoch) and Eocene (Eocene Epoch) epochs (65 million to 33.7 million years ago), it has seemed evident that bird groups other than those including Hesperornis and Ichthyornis must have existed during the Cretaceous. Knowledge of these, based on fragments of fossil bone, has slowly come to light, and there is now a fairly definite record from Cretaceous rock strata of other ancestral birds related to the living groups of loons (loon), grebes (grebe), flamingos (flamingo), cranes (crane), parrots (parrot), and shorebirds (shorebird)—and thus indication of early avian diversity. Therefore, it is clear that birds did not go through a “bottleneck” of extinction at the end of the Cretaceous that separated the archaic groups from the extant groups. Rather, the living groups were mostly present by the latest Cretaceous, and by this time the archaic groups seem to have died out.
Until comparatively recent times, the two groups of birds from Cretaceous time that received the most attention because of their strange form were the divers, such as , (Hesperornis) and the strong-winged , (Ichthyornis) a more ternlike (tern) form. Because they were the first well-known Cretaceous birds, having been described by American paleontologist O.C. Marsh (Marsh, Othniel Charles) in 1880, they were thought to represent typical Cretaceous birds. Recent discoveries, however, have changed this view. For example, members of one Early Cretaceous bird group, the Confuciusornithidae, showed very little advancement compared with Archaeopteryx and the Enantiornithes (a major group of birds widely distributed around the world through most of the Cretaceous Period). Because representatives of living bird groups have long been known among the fossil species from the Paleocene (Paleocene Epoch) and Eocene (Eocene Epoch) epochs (65 million to 33.7 million years ago), it has seemed evident that bird groups other than those including Hesperornis and Ichthyornis must have existed during the Cretaceous. Knowledge of these, based on fragments of fossil bone, has slowly come to light, and there is now a fairly definite record from Cretaceous rock strata of other ancestral birds related to the living groups of loons (loon), grebes (grebe), flamingos (flamingo), cranes (crane), parrots (parrot), and shorebirds (shorebird)—and thus indication of early avian diversity. Therefore, it is clear that birds did not go through a “bottleneck” of extinction at the end of the Cretaceous that separated the archaic groups from the extant groups. Rather, the living groups were mostly present by the latest Cretaceous, and by this time the archaic groups seem to have died out.Natural history
Habitats
Dinosaurs lived in many kinds of terrestrial environments, and although some remains, such as footprints, indicate where dinosaurs actually lived, their bones tell us only where they died (assuming that they have not been scattered or washed far from their place of death). Not all environments are equally well preserved in the fossil record. Upland environments, forests, and plains tend to experience erosion or decomposition of organic remains, so remains from these environments are rarely preserved in the geologic record. As a result, most dinosaur fossils are known from lowland environments, usually floodplains, deltas, lake beds, stream bottoms, and even some marine environments, where their bones apparently washed in after death. Much about the environments dinosaurs lived in can be learned from studying the pollen and plant remains preserved with them and from geochemical isotopes (isotope) that indicate temperature and precipitation levels. These climates, although free from the extensive ice caps of today and generally more equable, suffered extreme monsoon seasons and made much of the globe arid.
Only a few specimens represent the meagre beginning of the dinosaurian reign. This is probably because of a highly incomplete fossil record. Before dinosaurs appeared, all the continents of the world were joined to form one very large supercontinent called Pangea. Movements of the Earth's great crustal plates then began changing Earth's geography. By the Early Triassic Period (248 million to 206 million years ago), as dinosaurs were beginning to gain a foothold, Pangea had started to split apart at a rate averaging a few centimetres a year.
As the dinosaur line arose and experienced its initial diversification during the Late Triassic Period (227 million to 206 million years ago), the land areas of the world were in motion and drifting apart. Their respective inhabitants were consequently isolated from each other. Throughout the remainder of the Mesozoic Era, ocean barriers grew wider and the separate faunas became increasingly different. As the continents drifted apart, successive assemblages arose on each landmass and then diversified, waned, and disappeared, to be replaced by new fauna. By the Late Cretaceous Period (99 million to 65 million years ago), each continent occupied its own unique geographic position and climatic zone, and its fauna reflected that separation.
Food and feeding
The plant eaters
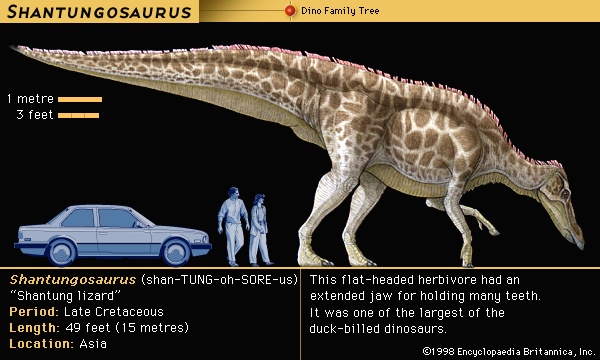 From the Triassic through the Jurassic and into the Cretaceous, the Earth's vegetation changed slowly but fundamentally from forests rich in gymnosperms (gymnosperm) (cycadeoids (Cycadeoidophyta), cycads (cycad), and conifers (conifer)) to angiosperm-dominated forests of palmlike trees and magnolia-like hardwoods. Although conifers continued to flourish at high latitudes, palms (palm) were increasingly confined to subtropical and tropical regions. These forms of plant life, the vast majority of them low in calories and proteins and made largely of hard-to-digest cellulose, became the foods of changing dinosaur communities. Accordingly, certain groups of dinosaurs, such as the ornithopods (ornithopod), included a succession of types that were increasingly adapted for efficient food processing. At the peak of the ornithopod lineage, the hadrosaurs (duck-billed dinosaurs of the Late Cretaceous) featured large dental batteries in both the upper and lower jaws, which consisted of many tightly compressed teeth that formed a long crushing or grinding surface. The preferred food of the duckbills cannot be certified, but at least one specimen found in Wyoming offers an intriguing clue: fossil plant remains in the stomach region have been identified as pine needles.
From the Triassic through the Jurassic and into the Cretaceous, the Earth's vegetation changed slowly but fundamentally from forests rich in gymnosperms (gymnosperm) (cycadeoids (Cycadeoidophyta), cycads (cycad), and conifers (conifer)) to angiosperm-dominated forests of palmlike trees and magnolia-like hardwoods. Although conifers continued to flourish at high latitudes, palms (palm) were increasingly confined to subtropical and tropical regions. These forms of plant life, the vast majority of them low in calories and proteins and made largely of hard-to-digest cellulose, became the foods of changing dinosaur communities. Accordingly, certain groups of dinosaurs, such as the ornithopods (ornithopod), included a succession of types that were increasingly adapted for efficient food processing. At the peak of the ornithopod lineage, the hadrosaurs (duck-billed dinosaurs of the Late Cretaceous) featured large dental batteries in both the upper and lower jaws, which consisted of many tightly compressed teeth that formed a long crushing or grinding surface. The preferred food of the duckbills cannot be certified, but at least one specimen found in Wyoming offers an intriguing clue: fossil plant remains in the stomach region have been identified as pine needles.The hadrosaurs' Late Cretaceous contemporaries, the ceratopsians (horned dinosaurs), had similar dental batteries that consisted of dozens of teeth. In this group the upper and lower batteries came together and acted as serrated shearing blades rather than crushing or grinding surfaces. Ordinarily, slicing teeth are found only in flesh-eating animals, but the bulky bodies and the unclawed, hooflike feet of dinosaurs such as Triceratops clearly are those of plant eaters. The sharp beaks and specialized shearing dentition of the ceratopsians suggest that they probably fed on tough, fibrous plant tissues, perhaps palm or cycad fronds.
 The giant sauropods (sauropod) such as Diplodocus and Apatosaurus must have required large quantities of plant food, but there is no direct evidence as to the particular plants they preferred. Because angiosperms rich in calories and proteins did not exist during most of the Mesozoic Era, it must be assumed that these sauropods fed on the abundant conifers and palm trees. Such a cellulose-heavy diet would have required an unusual bacterial population in the intestines to break down the fibre. A digestive tract with one or more crop chambers containing stones might have aided in the food-pulverizing process, but such gastroliths, or “stomach-stones,” are only rarely found in association with dinosaur skeletons. (A Seismosaurus specimen found with several hundred such stones is an important exception.)
The giant sauropods (sauropod) such as Diplodocus and Apatosaurus must have required large quantities of plant food, but there is no direct evidence as to the particular plants they preferred. Because angiosperms rich in calories and proteins did not exist during most of the Mesozoic Era, it must be assumed that these sauropods fed on the abundant conifers and palm trees. Such a cellulose-heavy diet would have required an unusual bacterial population in the intestines to break down the fibre. A digestive tract with one or more crop chambers containing stones might have aided in the food-pulverizing process, but such gastroliths, or “stomach-stones,” are only rarely found in association with dinosaur skeletons. (A Seismosaurus specimen found with several hundred such stones is an important exception.)The food preference of herbivorous dinosaurs can be inferred to some extent from their general body plan and from their teeth. It is probable, for example, that low-built animals such as the ankylosaurs, stegosaurs, and ceratopsians fed on low shrubbery. The tall ornithopods (ornithopod), especially the duckbills, and the long-necked sauropods (sauropod) probably browsed on high branches and treetops. No dinosaurs could have fed on grasses (family Poaceae), as these plants had not yet evolved.
The flesh eaters
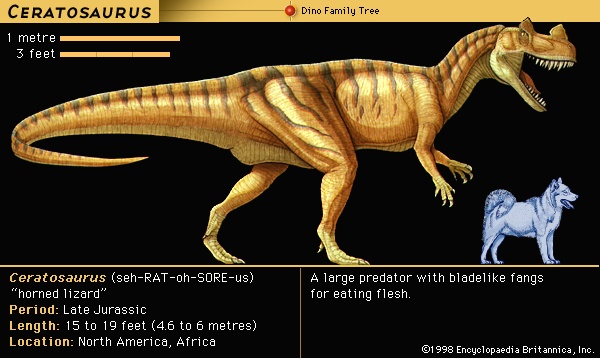 The flesh-eating dinosaurs came in all shapes and sizes and account for about 40 percent of the diversity of Mesozoic dinosaurs. They must have eaten anything they could catch, because predation is a highly opportunistic lifestyle. In several instances the prey victim of a particular carnivore has been established beyond much doubt. Remains were found of the small predator Compsognathus containing a tiny skeleton of the lizard Bavarisaurus in its stomach region. In Mongolia two different dinosaur skeletons were found together, a nearly adult-size Protoceratops in the clutches of its predator Velociraptor. Two of the many skeletons of Coelophysis discovered at Ghost Ranch in New Mexico, U.S., contained bones of several half-grown Coelophysis, apparently an early Mesozoic example of cannibalism. Fossilized feces (coprolites (coprolite)) from a large tyrannosaur contained crushed bone of another dinosaur. Skeletons of Deinonychus unearthed in Montana, U.S., were mixed with fragmentary bones of a much larger victim, the herbivore Tenontosaurus. This last example is significant because the multiple remains of the predator Deinonychus, associated with the bones of a single large prey animal, Tenontosaurus, strongly suggest that Deinonychus hunted in packs.
The flesh-eating dinosaurs came in all shapes and sizes and account for about 40 percent of the diversity of Mesozoic dinosaurs. They must have eaten anything they could catch, because predation is a highly opportunistic lifestyle. In several instances the prey victim of a particular carnivore has been established beyond much doubt. Remains were found of the small predator Compsognathus containing a tiny skeleton of the lizard Bavarisaurus in its stomach region. In Mongolia two different dinosaur skeletons were found together, a nearly adult-size Protoceratops in the clutches of its predator Velociraptor. Two of the many skeletons of Coelophysis discovered at Ghost Ranch in New Mexico, U.S., contained bones of several half-grown Coelophysis, apparently an early Mesozoic example of cannibalism. Fossilized feces (coprolites (coprolite)) from a large tyrannosaur contained crushed bone of another dinosaur. Skeletons of Deinonychus unearthed in Montana, U.S., were mixed with fragmentary bones of a much larger victim, the herbivore Tenontosaurus. This last example is significant because the multiple remains of the predator Deinonychus, associated with the bones of a single large prey animal, Tenontosaurus, strongly suggest that Deinonychus hunted in packs.Herding behaviour
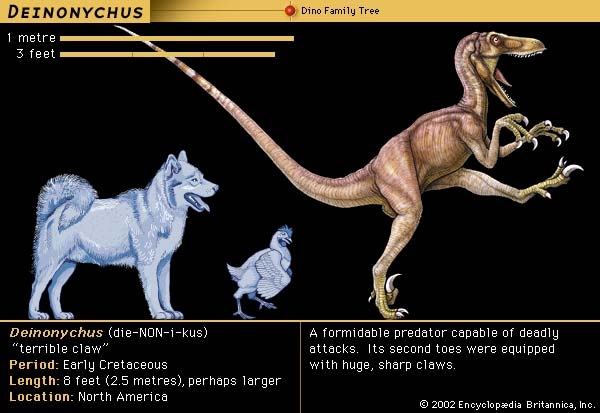 It should not come as a surprise that Deinonychus was a social animal, because many animals today are gregarious and form groups. Fossil evidence documents similar herding behaviour in a variety of dinosaurs. The mass assemblage in Bernissart, Belgium, for example, held at least three groups of Iguanodon. Group association and activity is also indicated by the dozens of Coelophysis skeletons of all ages recovered in New Mexico, U.S. The many specimens of Allosaurus at the Cleveland-Lloyd Quarry in Utah, U.S., may denote a herd of animals attracted to the site for the common purpose of scavenging. In the last two decades, several assemblages of ceratopsians and duckbills containing thousands of individuals have been found. Even Tyrannosaurus rex (tyrannosaur) is now known from sites where a group has been preserved together.
It should not come as a surprise that Deinonychus was a social animal, because many animals today are gregarious and form groups. Fossil evidence documents similar herding behaviour in a variety of dinosaurs. The mass assemblage in Bernissart, Belgium, for example, held at least three groups of Iguanodon. Group association and activity is also indicated by the dozens of Coelophysis skeletons of all ages recovered in New Mexico, U.S. The many specimens of Allosaurus at the Cleveland-Lloyd Quarry in Utah, U.S., may denote a herd of animals attracted to the site for the common purpose of scavenging. In the last two decades, several assemblages of ceratopsians and duckbills containing thousands of individuals have been found. Even Tyrannosaurus rex (tyrannosaur) is now known from sites where a group has been preserved together.These rare occurrences of multiple skeletal remains have repeatedly been reinforced by dinosaur footprints as evidence of herding. Trackways were first noted by Roland T. Bird in the early 1940s along the Paluxy riverbed in central Texas, U.S., where numerous washbasin-size depressions proved to be a series of giant sauropod footsteps preserved in limestone of the Early Cretaceous Period (144 million to 99 million years ago). Because the tracks are nearly parallel and all progress in the same direction, Bird concluded that “all were headed toward a common objective” and suggested that the sauropod trackmakers “passed in a single herd.” Large trackway sites also exist in the eastern and western United States, Canada, Australia, England, Argentina, South Africa, and China, among other places. These sites, dating from the Late Triassic Period (227 million to 206 million years ago) to the latest Cretaceous (65 million years ago), document herding as common behaviour among a variety of dinosaur types.
Some dinosaur trackways record hundreds, perhaps even thousands, of animals, possibly indicating mass migrations. The existence of so many trackways suggests the presence of great populations of sauropods, prosauropods, ornithopods (ornithopod), and probably most other kinds of dinosaurs. The majority must have been herbivores, and many of them were huge, weighing several tons or more. The impact of such large herds on the plant life of the time must have been great, suggesting constant migration in search of food.
Nesting sites discovered in the late 20th century also establish herding among dinosaurs. Nests and eggs numbering from dozens to thousands are preserved at sites that were possibly used for thousands of years by the same evolving populations of dinosaurs.
Growth and life span
Much attention has been devoted to dinosaurs as living animals—moving, eating, growing, reproducing biological machines. But how fast did they grow? How long did they live? How did they reproduce? The evidence concerning growth and life expectancy is sparse but growing. In the 1990s histological (histology) studies of fossilized bone by Armand de Ricqlès in Paris and R.E.H. Reid in Ireland showed that dinosaur skeletons grew quite rapidly. The time required for full growth has not been quantified for most dinosaurs, but de Ricqlès and his colleagues have shown that duckbills (hadrosaurs) such as Hypacrosaurus and Maiasaura reached adult size in seven or eight years and that the giant sauropods reached nearly full size in as little as 12 years. How long dinosaurs lived after reaching adult size is difficult to determine, but it is thought that the majority of known skeletons are not fully grown, because their bone ends and arches are very often not fused; in mature individuals these features would be fused.
reproduction
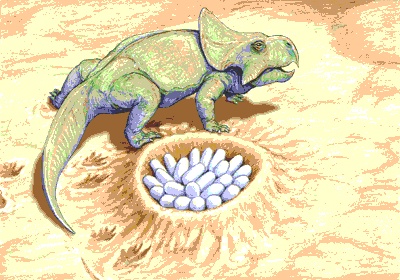 The idea that dinosaurs, like most living reptiles and birds, built nests and laid eggs had been widely debated even before the 1920s, when a team of scientists from the American Museum of Natural History, New York, made an expedition to Mongolia. Their discovery of dinosaur eggs in the Gobi Desert (Gobi) proved conclusively that at least one kind of dinosaur had been an egg layer and nest builder. These eggs were at first attributed to Protoceratops, but they are now known to have been those of Oviraptor. In 1978 John R. Horner and his field crews from Princeton University discovered dinosaur nests in western Montana. A few other finds, mostly of eggshell fragments from a number of sites, established oviparity as the only known mode of reproduction. In recent years an increasing number of dinosaur eggshells have been found and identified with the dinosaurs that laid them, and embryos have been found inside some eggs.
The idea that dinosaurs, like most living reptiles and birds, built nests and laid eggs had been widely debated even before the 1920s, when a team of scientists from the American Museum of Natural History, New York, made an expedition to Mongolia. Their discovery of dinosaur eggs in the Gobi Desert (Gobi) proved conclusively that at least one kind of dinosaur had been an egg layer and nest builder. These eggs were at first attributed to Protoceratops, but they are now known to have been those of Oviraptor. In 1978 John R. Horner and his field crews from Princeton University discovered dinosaur nests in western Montana. A few other finds, mostly of eggshell fragments from a number of sites, established oviparity as the only known mode of reproduction. In recent years an increasing number of dinosaur eggshells have been found and identified with the dinosaurs that laid them, and embryos have been found inside some eggs.The almost complete absence of juvenile dinosaur remains was puzzling until the 1980s. Horner, having moved to Montana State University, demonstrated that most paleontologists simply had not been exploring the right territory. After a series of intensive searches for the remains of immature dinosaurs, he succeeded beyond all expectations. The first such bones were unearthed near Choteau, Montana, and thereafter Horner and his crews discovered hundreds of nests, eggs, and newly hatched dinosaurs (mostly duckbills). Horner observed that previous explorations had usually concentrated on lowland areas, where sediments were commonly deposited and where most fossil remains were preserved. He recognized that such regions were not likely to produce dinosaur nests and young because they would have been hazardous places for nesting and raising the hatchlings. Upland regions would have been safer, but they were subject to erosion rather than deposition and were therefore less likely to preserve nests and eggs. However, it was exactly in such upland areas, close to the young and still-rising Rocky Mountains, that Horner made his discoveries.
Egg Mountain, as the area was named, produced some of the most important clues to dinosaurian habits yet found. For example, the sites show that a number of different dinosaur species made annual treks to this same nesting ground (though perhaps not all at the same time). Because of the succession of similar nests and eggs lying one on top of the other, it is thought that particular species returned to the same site year after year to lay their clutches. As Horner concluded, “site fidelity” was an instinctive part of dinosaurian reproductive strategy. This was confirmed more recently with the discovery of sauropod nests and eggs spread over many square kilometres in Patagonia, Argentina.
Body temperature
Beyond eating, digestion, assimilation, reproduction, and nesting, many other processes and activities went into making the dinosaur a successful biological machine. Breathing, fluid balance, temperature regulation (thermoregulation), and other such capabilities are also required. Dinosaurian body temperature regulation, or lack thereof, has been a hotly debated topic among students of dinosaur biology. Because it is obviously not possible to take an extinct dinosaur's temperature, all aspects of their metabolism and thermophysiology can be assessed only indirectly.
Ectothermy (cold-bloodedness) and endothermy
All animals thermoregulate (thermoregulation). The internal environment of the body is under the influence of both external and internal conditions. Land animals thermoregulate in several ways. They do so behaviorally, by moving to a colder or warmer place, by exercising to generate body heat, or by panting or sweating to lose it. They also thermoregulate physiologically, by activating internal metabolic processes that warm or cool the blood. But these efforts have limits, and, as a result, external temperatures and climatic conditions are among the most important factors controlling the geographic distribution of animals.
Today's so-called warm-blooded animals are the mammals (mammal) and birds (bird); reptiles (reptile), amphibians (amphibian), and most fishes (fish) are called cold-blooded. These two terms, however, are imprecise and misleading. Some “cold-blooded” lizards have higher normal body temperatures than do some mammals, for instance. Another pair of terms, ectothermy and endothermy, describes whether most of an animal's heat is absorbed from the environment (“ecto-”) or generated by internal processes (“endo-”). A third pair of terms, poikilothermy and homeothermy, describes whether the body temperature tends to vary with that of the immediate environment or remains relatively constant.
Today's mammals and birds have a high metabolism and are considered endotherms, which produce body heat internally. They possess biological temperature sensors that control heat production and switch on heat-loss mechanisms such as perspiration. Today's reptiles and amphibians, on the other hand, are ectotherms that mostly gain heat energy from sunlight, a heated rock surface, or some other external source. The endothermic state is effective but metabolically expensive, as the body must produce heat continuously, which requires correspondingly high quantities of fuel in the form of food. On the other hand, endotherms can be more active and survive lower external temperatures. Ectotherms do not require as much fuel, but most cannot deal as well with cold surroundings.
From the time of the earliest discoveries in the 19th century, dinosaur remains were classified as reptilian because their anatomic features are typical of living reptiles such as turtles (turtle), crocodiles (crocodile), and lizards (lizard). Because dinosaurs all have lower jaws constructed of several bones, a reptilian jaw joint, and a number of other nonmammalian, nonbirdlike characteristics, it was assumed that living dinosaurs were similar to living reptiles—scaly, cold-blooded, ectothermic egg layers (predominantly), not furry, warm-blooded live-bearers. A chauvinistic attitude seems to prevail that the warm-bloodedness of mammals is better than the cold-blooded reptilian state, even though turtles, snakes (snake), and other reptiles do very well regulating their body temperature in a different way. Moreover, both birds and mammals evolved from ectothermic, poikilothermic ancestors. At what point did metabolism heat up?
Clues to dinosaurian metabolism
The question of whether any extinct dinosaur was a true endotherm or homeotherm cannot be answered, but some interesting anatomic facts suggest these “warmer” possibilities. Probably the most direct evidence of dinosaurian physiology comes from bones themselves, particularly in regard to how they grew. The long bones (such as arm and leg bones) of most dinosaurs are composed almost exclusively of a well-vascularized type of bone matrix (fibro-lamellar) also found in most mammals and large birds. This type of bone tissue always indicates rapid growth, and it is very different from the more compact, poorly vascularized, parallel-fibred bone found in crocodiles and other reptiles and amphibians. It is generally thought that well-vascularized, rapidly growing bone can be sustained only by high metabolic rates that bring a continual source of nutrients and minerals to the growing tissues. It is difficult to explain these histological features in any other metabolic terms. On the other hand, most dinosaurs retain lines of arrested growth (LAGs) in most of their long bones. LAGs are found in other reptiles, amphibians, and fishes, and they often reflect a seasonal period during which metabolism slows, usually because of environmental stresses. This slowdown produces “rest lines” as LAGs in the bones. The presence of these lines in dinosaur bones has been taken as an indication that they were metabolically incapable of growing throughout the year. However, LAGs in dinosaurs are less pronounced than in other reptiles; LAGs can also appear in different numbers in different bones of the same skeleton, and they are sometimes even completely absent. Finally, some living birds and mammals, which are clearly endotherms, have LAGs very much like those of dinosaurs, so LAGs are probably not strong indicators of metabolism in any of these animals.
Other, less direct lines of evidence may reveal other clues about dinosaurian metabolism. Two dinosaurian groups, the hadrosaurs and the ceratopsians (ceratopsian), had highly specialized sets of teeth that were obviously effective at processing food. Both groups were herbivorous, but unlike living reptiles they chopped and ground foliage thoroughly. Such highly efficient dentitions may suggest a highly effective digestive process that would allow more energy to be extracted from the food. This feature by itself, however, may not be crucial. Pandas (panda), for example, are not very efficient in digesting plant material, but they survive quite well on a diet of almost nothing but bamboo.
Another line of evidence is that dinosaurs had anatomic features reflecting a high capacity for activity. The first dinosaurs walked upright, holding their legs under their bodies; they could not sprawl. This indicates that, by standing and walking all day, they probably expended more energy than reptiles, which typically sit and wait for prey. As some lineages of dinosaurs grew larger, they reverted to four-legged (quadrupedal) locomotion, but their stance was still upright. They also put one foot directly in front of the other when they walked (parasagittal gait), instead of swinging the limbs to the side. Such posture and gait are present in all nonaquatic endotherms (mammals and birds) today, whereas a sprawling or semierect posture is typical of all ectotherms (reptiles and amphibians). Bipedal stance and parasagittal gait are not sustained in any living ectotherm, perhaps because they require a relatively higher level of sustained energy.
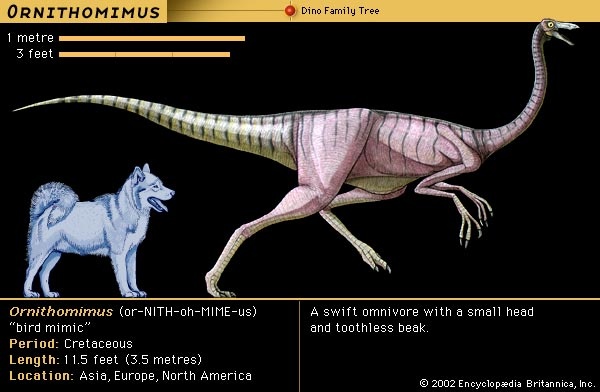 The high speeds at which some dinosaurs must have traveled have also been invoked as evidence of high metabolic levels. For example, the ostrichlike dinosaurs, such as Struthiomimus, Ornithomimus, Gallimimus, and Dromiceiomimus, had long hind legs and must have been very fleet. The dromaeosaurs (dromaeosaur), such as Deinonychus, Velociraptor, and Dromaeosaurus, also were obligatory bipeds. They killed prey with talons on their feet, and one can argue that it must have taken a high level of metabolism to generate the degree of activity and agility required of such a skill. However, most ectotherms can move very rapidly in bursts of activity such as running and fighting, so this feature may not provide conclusive evidence either.
The high speeds at which some dinosaurs must have traveled have also been invoked as evidence of high metabolic levels. For example, the ostrichlike dinosaurs, such as Struthiomimus, Ornithomimus, Gallimimus, and Dromiceiomimus, had long hind legs and must have been very fleet. The dromaeosaurs (dromaeosaur), such as Deinonychus, Velociraptor, and Dromaeosaurus, also were obligatory bipeds. They killed prey with talons on their feet, and one can argue that it must have taken a high level of metabolism to generate the degree of activity and agility required of such a skill. However, most ectotherms can move very rapidly in bursts of activity such as running and fighting, so this feature may not provide conclusive evidence either.
 Related to the upright posture of many dinosaurs is the fact that the head was often positioned well above the level of the heart. In some sauropods (sauropod) ( Apatosaurus, Diplodocus, Brachiosaurus (brachiosaur), and Barosaurus, for instance), the brain must have been several metres above the heart. The physiological importance of this is that a four-chambered heart would be required for pumping freshly oxygenated blood to the brain. Brain death follows very quickly when nerve cells are deprived of oxygen, and to prevent it most dinosaurs must have required two ventricles (ventricle). In a four-chambered heart, one ventricle pumps oxygen-poor venous blood at low pressure to the lungs to absorb fresh oxygen (high pressure would rupture capillaries (capillary) of the lungs). A powerful second ventricle pumps freshly oxygenated blood to all other parts of the body at high pressure. To overcome the weight of the column of blood that must be moved from the heart to the elevated brain, high pressure is certainly needed. In short, like birds and mammals, many dinosaurs apparently had the required four-chambered heart necessary for an animal with a high metabolism.
Related to the upright posture of many dinosaurs is the fact that the head was often positioned well above the level of the heart. In some sauropods (sauropod) ( Apatosaurus, Diplodocus, Brachiosaurus (brachiosaur), and Barosaurus, for instance), the brain must have been several metres above the heart. The physiological importance of this is that a four-chambered heart would be required for pumping freshly oxygenated blood to the brain. Brain death follows very quickly when nerve cells are deprived of oxygen, and to prevent it most dinosaurs must have required two ventricles (ventricle). In a four-chambered heart, one ventricle pumps oxygen-poor venous blood at low pressure to the lungs to absorb fresh oxygen (high pressure would rupture capillaries (capillary) of the lungs). A powerful second ventricle pumps freshly oxygenated blood to all other parts of the body at high pressure. To overcome the weight of the column of blood that must be moved from the heart to the elevated brain, high pressure is certainly needed. In short, like birds and mammals, many dinosaurs apparently had the required four-chambered heart necessary for an animal with a high metabolism.The significance of thermoregulation can be seen by comparing today's reptiles with mammals. The rate of metabolism is usually measured in terms of oxygen consumed per unit of body weight per unit of time. The resting metabolic rate for most mammals is about 10 times that of modern reptiles, and the range of metabolic rates among living mammals is about double that seen among reptiles. These differences mean that endothermic mammals have much more endurance than their cold-blooded counterparts. Some dinosaurs may have been so endowed, and although they seem to have possessed the cardiovascular system (human cardiovascular system) necessary for endothermy, that capacity does not conclusively prove that they were endothermic. There exists the possibility that dinosaurs were neither complete ectotherms nor complete endotherms. Rather, they may have evolved a range of metabolic strategies, much as mammals have (as is illustrated by the differences between sloths (sloth) and cheetahs (cheetah), bats (bat) and whales (whale), for example).
Classification (taxonomy)
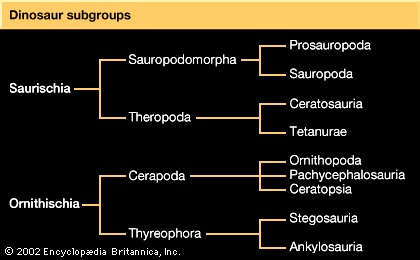 The chief difference between the two major groups of dinosaurs is in the configuration of the pelvis (pelvic girdle). It was primarily on this distinction that the English biologist H.G. Seeley established the two dinosaurian orders and named them Saurischia (saurischian) (“lizard hips”) and Ornithischia (ornithischian) (“bird hips”) in 1887; this differentiation is still maintained.
The chief difference between the two major groups of dinosaurs is in the configuration of the pelvis (pelvic girdle). It was primarily on this distinction that the English biologist H.G. Seeley established the two dinosaurian orders and named them Saurischia (saurischian) (“lizard hips”) and Ornithischia (ornithischian) (“bird hips”) in 1887; this differentiation is still maintained. As in all four-legged animals, the dinosaurian pelvis was a paired structure consisting of three separate bones on each side that attached to the sacrum of the backbone. The ilium was attached to the spine, and the pubis and ischium were below, forming a robust bony plate. At the centre of each plate was a deep cup—the hip socket (acetabulum). The hip socket faced outward and was open at its centre for the articulation of the thighbone. The combined saurischian pelvic bones presented a triangular outline as seen from the side, with the pubis extending down and forward and the ischium projecting down and backward from the hip socket. The massive ilium formed a deep vertical plate of bone to which the muscles of the pelvis, hind leg, and tail were attached. The pubis had a stout shaft, commonly terminating in a pronounced expansion or bootlike structure (presumably for muscle attachment) that solidly joined its opposite mate. The ischium was slightly less robust than the pubis, but it too joined its mate along a midline. There were minor variations in this structure between the various saurischians.
As in all four-legged animals, the dinosaurian pelvis was a paired structure consisting of three separate bones on each side that attached to the sacrum of the backbone. The ilium was attached to the spine, and the pubis and ischium were below, forming a robust bony plate. At the centre of each plate was a deep cup—the hip socket (acetabulum). The hip socket faced outward and was open at its centre for the articulation of the thighbone. The combined saurischian pelvic bones presented a triangular outline as seen from the side, with the pubis extending down and forward and the ischium projecting down and backward from the hip socket. The massive ilium formed a deep vertical plate of bone to which the muscles of the pelvis, hind leg, and tail were attached. The pubis had a stout shaft, commonly terminating in a pronounced expansion or bootlike structure (presumably for muscle attachment) that solidly joined its opposite mate. The ischium was slightly less robust than the pubis, but it too joined its mate along a midline. There were minor variations in this structure between the various saurischians.The ornithischian pelvis was constructed of the same three bones on each side of the sacral vertebrae, to which they were attached. The lateral profile of the pelvis was quite different from that of the saurischians, which had a long but low iliac blade above the hip socket and a modified ischium-pubis structure below. Here the long, thin ischium extended backward and slightly downward from the hip socket. In the most primitive, or basal, ornithischians, the pubis had a moderately long anterior blade, but this was reduced in later ornithischians. Posteriorly it stretched out into a long, thin postpubic process lying beneath and closely parallel to the ischium. The resulting configuration superficially resembled that of birds, whose pubis is a thin process extending backward beneath the larger ischium. These anatomic dissimilarities are thought to reflect important differences in muscle arrangements in the hips and hind legs of these two orders. However, the soft parts of these dinosaurs are not well enough understood to reveal any functional or physiological basis for the differences. Other marked dissimilarities between saurischians and ornithischians are found in their jaws and teeth, their limbs, and especially their skulls. Details regarding these differences are given in the following discussions of the major dinosaur groups.
The classification shows how the groups are subdivided. This classification is based on their relationships to each other, as far as they are known. Fossil remains are often difficult to interpret, especially when only a few fragmentary specimens of a type have been found. No universally accepted classification of dinosaurs exists. Occasionally, for example, the Sauropodomorpha have been divided into more or fewer lower-rank categories (e.g., families, subfamilies), and the suborder Theropoda has been divided into two infraorders, the Carnosauria and the Coelurosauria. Increasingly, taxonomists (taxonomy) have abandoned the traditional Linnaean ranks of family, order, and so on because they are cumbersome and not comparable among different kinds of organisms. Instead, the names of the groups alone are used without denoting a category. Generally, a phylogeny such as the accompanying diagram clearly shows which groups are subsumed under others. Additionally, words with similar roots but different endings may indicate more or less inclusive groups. Ornithomimosauria, for example, denotes a more inclusive group than Ornithomimidae. Because the results of different phylogenetic analyses vary among researchers, and will continue to change as new specimens and taxa (taxon) are discovered, the classification can be expected to change accordingly. This is a normal part of scientific activity and reflects continuing growth of knowledge and reappraisal of current understanding.Saurischia (saurischian)
Saurischians are known from specimens ranging from the Late Triassic to the present day, because, as will be seen, birds are highly derived saurischian dinosaurs. Two distinctly different groups are traditionally included in the saurischians—the Sauropodomorpha (herbivorous sauropods and prosauropods) and the Theropoda (carnivorous dinosaurs). These groups are placed together on the basis of a suite of features that they share uniquely. These include elongated posterior neck vertebrae, accessory articulations on the trunk vertebrae, and a hand that is nearly half as long as the rest of the arm (or longer). In addition, the second finger of the hand (not the third, as in other animals) is invariably the longest; the thumb is borne on a short metacarpal bone that is offset at its far end, so that the thumb diverges somewhat from the other fingers. The first joint of the thumb, which bears a robust claw, is longer than any other joint in the hand.
Sauropodomorpha
Included in this group are the well-known sauropods, or “brontosaur” types, and their probable ancestral group, the prosauropods. All were plant eaters, though their relationship to theropods, along with the fact that the closest relatives of dinosaurs were evidently carnivorous, suggests that they evolved from meat eaters. Sauropodomorpha are distinguished by leaf-shaped tooth crowns, a small head, and a neck that is at least as long as the trunk of the body and longer than the limbs.
Prosauropoda
Most generalized of the Sauropodomorpha were the so-called prosauropods. Found from the Late Triassic (Triassic Period) to Early Jurassic (Jurassic Period) periods (227 million to 180 million years ago), their remains are probably the most ubiquitous of all Triassic dinosaurs. They have been found in Europe (Germany), North America (New England, Arizona, and New Mexico), South America (Argentina), Africa (South Africa, Lesotho, Zimbabwe), China (Yunnan), and Antarctica. The best-known examples include Plateosaurus of Germany and Massospondylus of South Africa. Prosauropods were not especially large; they ranged from less than 2 metres (7 feet) in length up to about 8 metres (26 feet) and up to several tons in maximum weight. Many of these animals are known from very complete skeletons (especially the smaller, more lightly built forms). Because their forelimbs are conspicuously shorter than their hind limbs, they have often been reconstructed poised on their hind legs in a bipedal stance. Their anatomy, however, clearly indicates that some of them could assume a quadrupedal (four-footed) position. Footprints generally attributed to prosauropods appear to substantiate both forms of locomotion.Prosauropods have long been seen as including the first direct ancestors of the giant sauropods, probably among the melanorosaurids. That view has long prevailed largely because of their distinctly primitive sauropod-like appearance and also because of their Late Triassic–Early Jurassic occurrence. No better candidate has been discovered, and the first true sauropods are not found until the Early Jurassic, so the transition between prosauropods and sauropods has been generally accepted. In the 1990s, however, several studies have suggested that prosauropods may be a distinct group that shared common ancestors with sauropods earlier in the Triassic. If this view is correct, it is mystifying why the smaller prosauropods are so widespread throughout the Late Triassic, yet none of the larger and more conspicuous sauropods have been found from that period.
In general body form, prosauropods were mostly rather stocky, with a long, moderately flexible neck containing surprisingly long and flexible cervical ribs. The head was small in comparison with the body. The jaw was long and contained rows of thin, leaflike teeth suited for chopping up (but not grinding or crushing) plant tissues, although there is an indication of direct tooth-on-tooth occlusion.
Prosauropod forelimbs were stout, with five complete digits. The hind limbs were about 50 percent longer than the forelimbs and even more heavily built. The foot was of primitive design, and its five-toed configuration could be interpreted as a forerunner of the sauropod foot. Walking apparently was done partly on the toes (semidigitigrade), with the metatarsus (metatarsal) held well off the ground. The vertebral column was unspecialized and bore little indication of the cavernous excavations that were to come in later sauropod vertebrae, nor did it show projections that were to buttress the sauropod vertebral column. The long tail probably served as a counterweight or stabilizer whenever the animal assumed a bipedal position.
Sauropoda (sauropod)
The more widely known sauropods—the huge “brontosaurs” and their relatives—varied in length from 6 or 7 metres (about 20 feet) in the primitive ancestral sauropod Vulcanodon of Africa, Barapasaurus of India, and Ohmdenosaurus of Germany, up to 28 to 30 metres (90 to 100 feet) or more in Late Jurassic North American forms such as Apatosaurus (formerly known as Brontosaurus), Diplodocus, Seismosaurus, and Sauroposeidon. Weights ranged from about 20 tons or less in Barapasaurus to 80 tons or more for the gigantic Brachiosaurus (brachiosaur) of Africa and North America. Sauropods were worldwide in distribution but have not as yet been found in Antarctica. In geologic time they ranged from the Late Triassic (Triassic Period) Riojasaurus to the Late Cretaceous (Cretaceous Period) Alamosaurus of North America and Laplatasaurus of South America. Their greatest diversity and abundance took place 120 million–150 million years ago, during the Late Jurassic (Jurassic Period) and Early Cretaceous periods.
Sauropods are notable for their body form as well as their enormous size. Their large bodies were heart-shaped in cross section, like elephants, with long (sometimes extremely long) necks and tails. Their columnar legs, again like those of elephants, had little freedom to bend at the knee and elbow. The legs were maintained in a nearly vertical position beneath the shoulder and hip sockets. Because of their great bulk, sauropods unquestionably were obligate quadrupeds.
The sauropod limb bones were heavy and solid. The feet were broad, close to plantigrade (adapted for walking on the soles), and graviportal (adapted for bearing great weight). The toes were generally short, blunt, and broad, but some sauropods had a large straight claw on the first digit of the forefoot and the first and second toes of the hind foot. These animals must have moved relatively slowly and with only short steps because of the comparative inflexibility of the limbs. Running must have been stiff-legged at no better than an elephantine pace of 16 km (10 miles) per hour, if that. Their tremendous bulk placed them out of the reach of predators and eliminated any need for speed. Evidently their fast growth was adaptive to predator avoidance.
The vertebrae of the backbone were highly modified, with numerous excavations and struts to reduce bone weight. Complex spines and projections for muscle and ligament attachment compensated for any loss of skeletal strength that resulted from reductions in bone density and mass. The long and sometimes massive tail, characteristic of so many sauropods, would appear to have been carried well off the ground. Tail drag marks associated with sauropod trackways are not known, and damaged (stepped-on) tails are also not known, even though these animals apparently traveled in herds (albeit of undetermined density). Another possible use of the tail, like the neck, may have been thermal regulation, as improved heat loss through its large surface area could have been a result. The tail was also the critical anchor of the large, powerful hind leg muscles that produced most of the walking force required for moving the many tons of sauropod weight. The muscle arrangement of the tail was precisely that of modern alligators and lizards.
The most important part of any skeleton is the skull because it provides the most information about an animal's mode of life and general biology. Sauropod skulls were of several main types, including the high, boxy Camarasaurus type (often incorrectly associated with Apatosaurus); the shoe-shaped Brachiosaurus type, with its large, delicately arched nasal bones; and the low, narrow, streamlined, almost horselike Diplodocus type. The first had broad, spatulate teeth, while the latter two had narrow, pencil-shaped teeth largely confined to the front parts of the jaws, especially in diplodocids.
Until recently, sauropods were visualized as swamp or lake dwellers because their legs were thought to be incapable of supporting their great weights or because such huge creatures would naturally prefer the buoyancy of watery surroundings. The 19th-century English biologist Richard Owen (Owen, Sir Richard), in fact, identified the first known sauropods as giant aquatic crocodiles and called them cetiosaurs (whale lizards) because they were so large and because they were found in aquatic sediments. Eventually enough skeletal remains were discovered to show that these animals were neither crocodiles nor aquatic. However, the image of amphibious habits, thought necessary to support the great weights of sauropods, persisted for a long time, however incorrectly. Experiments with fresh bone samples have shown that bone of the type that composed the sauropods' limb bones could easily have supported their estimated weights. Moreover, there is no feature in their skeletons that suggests an aquatic, or even amphibious, existence. In addition, numerous trackway sites clearly prove that sauropods could navigate on land, or at least where the water was too shallow to buoy up their weight. Accordingly, newer interpretations see these animals as floodplain and forest inhabitants.
Still another blow has been dealt to the old swamp image by the physical laws of hydrostatic pressure, which prohibit the explanation that the long neck enabled a submerged animal to raise its head to the surface for a breath of fresh air. The depth at which the lungs would be submerged would not allow them to be expanded by normal atmospheric pressure, the only force that fills the lungs. Consequently, the long necks of sauropods must be explained in terms of terrestrial functions such as elevating the feeding apparatus or the eyes. On all counts, sauropods are best seen as successful giraffelike browsers and only occasional waders.
Theropoda (theropod)
 This group includes all the known carnivorous dinosaurs as well as the birds (bird). No obviously adapted herbivores are recognized in the group, but some theropods, notably the toothless oviraptorids and ornithomimids, may well have been relatively omnivorous like today's ostriches (ostrich). Mesozoic Era theropods ranged in size from the smallest known adult Mesozoic nonavian dinosaur, the crow-sized Microraptor, up to the great Tyrannosaurus and Giganotosaurus, which were 15 or more metres (50 feet) long, more than 5 metres (16 to 18 feet) tall, and weighed 6 tons or more. Theropods have been recovered from deposits of the Late Triassic through the latest Cretaceous and from all continents.
This group includes all the known carnivorous dinosaurs as well as the birds (bird). No obviously adapted herbivores are recognized in the group, but some theropods, notably the toothless oviraptorids and ornithomimids, may well have been relatively omnivorous like today's ostriches (ostrich). Mesozoic Era theropods ranged in size from the smallest known adult Mesozoic nonavian dinosaur, the crow-sized Microraptor, up to the great Tyrannosaurus and Giganotosaurus, which were 15 or more metres (50 feet) long, more than 5 metres (16 to 18 feet) tall, and weighed 6 tons or more. Theropods have been recovered from deposits of the Late Triassic through the latest Cretaceous and from all continents.Theropods may be defined as birds and all saurischians more closely related to birds than to sauropods (sauropod). They have a carnivorous dentition and large, recurved claws on the fingers. They also share many other characteristics, such as a distinctive joint in the lower jaw, epipophyses on the neck vertebrae, and a unique “transition point” in the tail where the vertebrae become longer and more lightly built. Other similarities include the reduction or loss of the outer two fingers, long end joints of the fingers, and a straplike fibula attached to a crest on the side of the tibia.
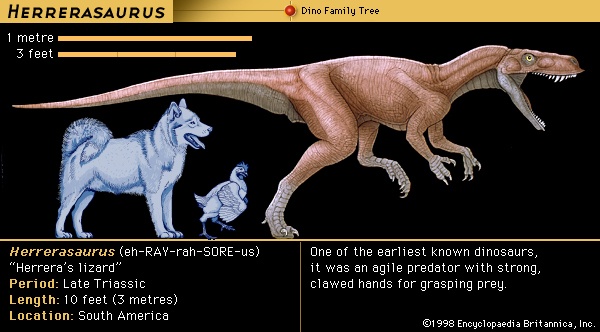 Herrerasaurus and several fragmentary taxa from South America, including Staurikosaurus and Ischisaurus, from the Middle to Late Triassic of Argentina are carnivores that have often been included in the Dinosauria, specifically in Theropoda. Whereas these animals closely resemble dinosaurs and have many carnivorous features, they also lack a number of features present in dinosaurs, saurischians, and theropods. For example, they have only two sacral vertebrae, unlike dinosaurs; their hips are more primitive than those of saurischians, as are their wrists; and the second finger is not the longest, unlike those of all saurischians. It remains probable that the features they seem to share with theropod dinosaurs are simply primitive and related to carnivory, the general habit of archosaurs (archosaur). Future discoveries and analyses may help to resolve these questions.
Herrerasaurus and several fragmentary taxa from South America, including Staurikosaurus and Ischisaurus, from the Middle to Late Triassic of Argentina are carnivores that have often been included in the Dinosauria, specifically in Theropoda. Whereas these animals closely resemble dinosaurs and have many carnivorous features, they also lack a number of features present in dinosaurs, saurischians, and theropods. For example, they have only two sacral vertebrae, unlike dinosaurs; their hips are more primitive than those of saurischians, as are their wrists; and the second finger is not the longest, unlike those of all saurischians. It remains probable that the features they seem to share with theropod dinosaurs are simply primitive and related to carnivory, the general habit of archosaurs (archosaur). Future discoveries and analyses may help to resolve these questions.In all theropods the hind leg bones were hollow to varying degrees—extremely hollow and lightly built in small to medium-size members ( Compsognathus, Coelurus, and Ornitholestes, among others) and more solid in the larger forms (such as Allosaurus, Daspletosaurus, and Tarbosaurus).
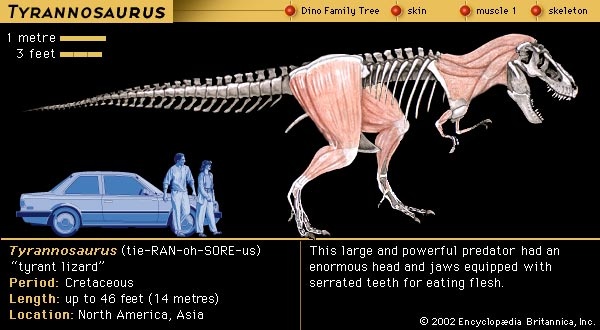 In stance and gait, theropods were obligatory bipeds. Their bodies conformed to a common shape in which the hind legs were dominant and designed for support and locomotion. The forelimbs, on the other hand, had been modified from the primitive design and entirely divested of the functions of locomotion and body support. Hind limbs were either very robust and of graviportal (weight-bearing) proportions, as in Allosaurus, Megalosaurus, and the tyrannosaurids (tyrannosaur), or very slender, elongated, and of cursorial (adapted for running) proportions, as in Coelurus, Coelophysis, Ornitholestes, and the ornithomimids. Theropod feet, despite the group's name, which means “beast (i.e., mammal) foot,” usually looked much like those of birds, which is not surprising, because birds inherited their foot structure from these dinosaurs. Three main toes were directed forward and splayed in a V-shaped arrangement; an additional inside toe was directed medially or backward. The whole foot was supported by the toes (digitigrade), with the “heel” elevated well above the ground. Toes usually bore sharp, somewhat curved claws.
In stance and gait, theropods were obligatory bipeds. Their bodies conformed to a common shape in which the hind legs were dominant and designed for support and locomotion. The forelimbs, on the other hand, had been modified from the primitive design and entirely divested of the functions of locomotion and body support. Hind limbs were either very robust and of graviportal (weight-bearing) proportions, as in Allosaurus, Megalosaurus, and the tyrannosaurids (tyrannosaur), or very slender, elongated, and of cursorial (adapted for running) proportions, as in Coelurus, Coelophysis, Ornitholestes, and the ornithomimids. Theropod feet, despite the group's name, which means “beast (i.e., mammal) foot,” usually looked much like those of birds, which is not surprising, because birds inherited their foot structure from these dinosaurs. Three main toes were directed forward and splayed in a V-shaped arrangement; an additional inside toe was directed medially or backward. The whole foot was supported by the toes (digitigrade), with the “heel” elevated well above the ground. Toes usually bore sharp, somewhat curved claws. The forelimbs varied widely from the slender, elongated ones of Struthiomimus, for example, to shorter, more massively constructed grasping appendages like those of Allosaurus, to the greatly abbreviated arms and hands of Tyrannosaurus, to the abbreviated, stout limb and single finger of Mononykus, to the range of wings now seen in birds. The hands typically featured long, flexible fingers with pronounced, often strongly curved claws, which bore sharp piercing talons. Early theropods such as Coelophysis had four fingers, with the fifth reduced to a nubbin of the metacarpal and the fourth greatly reduced. Most theropods were three-fingered, having lost all remnants of the fourth and fifth fingers. Tyrannosaurids (including Albertosaurus, Daspletosaurus, Tarbosaurus, and Tyrannosaurus) were notable for their two-fingered hands and unusually short arms; they had lost the third finger. The odd Mononykus lost even its second finger, retaining only a bizarre thumb. This separation of function between fore and hind limbs was a feature of the first dinosaurs. Although the first theropods, sauropodomorphs, and ornithischians were all bipedal, only theropods remained exclusively so.
The forelimbs varied widely from the slender, elongated ones of Struthiomimus, for example, to shorter, more massively constructed grasping appendages like those of Allosaurus, to the greatly abbreviated arms and hands of Tyrannosaurus, to the abbreviated, stout limb and single finger of Mononykus, to the range of wings now seen in birds. The hands typically featured long, flexible fingers with pronounced, often strongly curved claws, which bore sharp piercing talons. Early theropods such as Coelophysis had four fingers, with the fifth reduced to a nubbin of the metacarpal and the fourth greatly reduced. Most theropods were three-fingered, having lost all remnants of the fourth and fifth fingers. Tyrannosaurids (including Albertosaurus, Daspletosaurus, Tarbosaurus, and Tyrannosaurus) were notable for their two-fingered hands and unusually short arms; they had lost the third finger. The odd Mononykus lost even its second finger, retaining only a bizarre thumb. This separation of function between fore and hind limbs was a feature of the first dinosaurs. Although the first theropods, sauropodomorphs, and ornithischians were all bipedal, only theropods remained exclusively so.The jaws of theropods are noted for their complement of sharp, bladelike teeth. In nearly all theropods these laterally compressed blades had serrations along the rear edge and often along the front edge as well. Tyrannosaur teeth differed in having a rounder, less-compressed cross section, better adapted to puncture flesh and tear it from bone. Troodontid teeth had recurved serrations slightly larger than those typical of theropods. Archaeopteryx and other basal birds had narrow-waisted teeth with greatly reduced serrations or none at all. Some theropods, such as most ornithomimids and oviraptorids, had lost most or all of their teeth.
In recent years a series of unusually well-preserved theropod dinosaurs have been discovered in deposits from the Early Cretaceous Period (144 million to 99 million years ago) in Liaoning province, China. These theropods have filamentous integumentary (integument) structures of several kinds that resemble feathers (feather). Such structures indicate that today's birds very likely evolved from theropod dinosaurs. See Dinosaur descendants (dinosaur).
Ceratosauria
Ceratosauria includes Ceratosaurus and all theropods more closely related to it than to birds. This group includes basal theropods such as Dilophosaurus and Coelophysis. It may also include the abelisaurids of South America and elsewhere, but this is not certain. Originally thought to be a natural group, Ceratosauria, as traditionally constituted, may represent a more general grouping of basal theropods, including the ancestral stock of most later theropods. The Late Triassic Coelophysis, about 1.5 meters long, is generally regarded as an archetypal primitive theropod. It has a long neck and a long, low head with numerous small, sharp, recurved teeth. The legs were long, the arms relatively short, and the tail very long. Dilophosaurus, from the Early Jurassic Period (206 million to 180 million years ago), is considerably larger (about 4 metres total length) and is distinguished by a pair of thin bony crests running along the top of the skull. Because no other theropod had such structures, these were apparently not necessary for any physiological function and so are thought to have been for display or species recognition. There is no evidence that Dilophosaurus spat venom.Tetanurae
These comprise birds and all the theropods closer to birds than to Ceratosaurus. They would include the true carnosaurs (carnosaur) and coelurosaurs described below as well as a few relatively large carnivorous basal forms (such as Torvosaurus, Spinosaurus, Baryonyx, Afrovenator, and Megalosaurus). The tetanuran theropods are distinguished by several features, including the complete loss of digits four and five of the hand, an upper tooth row extending backward only to the eye, and a fibula that is reduced and clasped by the tibia. The name Tetanurae, or “stiff tails,” refers to another unusual feature, a transition point in the tail sequence where the vertebrae change form in a distinctive way.
Carnosauria includes Allosaurus and all theropods more closely related to it than to birds, including forms such as Acrocanthosaurus, Sinraptor, and Giganotosaurus. The first known members appear in the Late Jurassic and persist into the Cretaceous. Originally, this group was designed to include all the big predatory dinosaurs, but it was recently recognized that only size, not their relationships, was the trait unifying this group. Some, such as Dilophosaurus and Carnotaurus, were probably more closely related to basal ceratosaurs. Others, such as Baryonyx and Spinosaurus, represented an unusual diversification of fish-eating forms that were almost crocodilian in some of their habits. Still others, such as Tyrannosaurus and its relatives, the albertosaurs and daspletosaurs, were probably just giant coelurosaurs, as had been hypothesized by German paleontologist Friedrich von Huene early in the 20th century. As these groups were removed from the original Carnosauria, only Allosaurus and its relatives of the great Late Jurassic and Early Cretaceous diversification were left. Along with Torvosaurus and the megalosaurs, they must have been among the most deadly and rapacious large predators of their time. They are distinguished by relatively few characteristics. It is commonly thought that carnosaurs had very short limbs, but this is not particularly true—they were proportionally much shorter in tyrannosaurs, which are no longer considered carnosaurs. True carnosaurs had limbs comparable in size to those of more basal theropods. Sauropod vertebrae have been found with carnosaur tooth marks in them, which attests to the predatory habits of these dinosaurs.
The coelurosaurs (“hollow-tailed reptiles”) include generally small to medium-size theropods, though the recent inclusion of tyrannosaurs would seem to discount this generalization. Coelurosauria is defined as birds and all tetanurans more closely related to birds than to the carnosaurs. The first known members, including birds, appear in the Late Jurassic; the great Cretaceous diversification of the other coelurosaurs ended with the Cretaceous extinctions.
In coelurosaurs the pelvis is modified so that the ischium is reduced to two-thirds or less the size of the pubis; the eyes are larger, and no more than 15 tail vertebrae bear transverse projections. Each of the various coelurosaurian groups has very distinct features that sets it apart from the others. The most basal known form, the Late Jurassic Compsognathus, was the size of a chicken and contemporaneous with the first known bird, Archaeopteryx. However, the two animals were not as closely related as some other coelurosaurs were to birds.Tyrannosaurs (tyrannosaur) and the related albertosaurs (Albertosaurus) were the largest of the Late Cretaceous theropods of the northern continents. They are distinguished by an exceptionally large, high skull and teeth with a much more rounded cross section than the typical daggerlike teeth of other theropods. Their forelimbs are very short, and the third finger is reduced to a splint or lost entirely. Tyrannosaurs are thought to have migrated to North America from Asia, because early relatives first appear on the latter continent. Although there has been some debate about whether tyrannosaurs were active predators or more passive scavengers, the distinction is not usually strong in living predatory animals, and frequently larger carnivores will chase smaller ones away from fresh kills. However, some skeletons of plant-eating dinosaurs evidently have healed wounds caused by tyrannosaur bites, so active predation appears to be sustained.
Ornithomimids were medium-size to large theropods. Almost all of them were toothless, and apparently their jaws were covered by a horny beak; they also had very long legs and arms. A well-known example is Struthiomimus. Most were ostrich-sized and were adapted for fast running, with particularly long foot bones, or metatarsals (metatarsal). The largest was Deinocheirus from Asia, known only from one specimen consisting of complete arms and hands almost 3 metres (10 feet) long—nearly four times longer than those of Struthiomimus. These animals' speed, toothlessness, and long hands with relatively symmetrical fingers leave their lifestyle and feeding habits unclear, but they may have been fairly omnivorous like ostriches, although they are not directly related.Oviraptorids, therizinosaurids, and caenagnathids appear to form a clade slightly more related to birds than to the coelurosaurs. Oviraptorids, known from the Late Cretaceous of Mongolia, had very strange skulls, often with high crests and a reduced dentition in an oddly curved jaw. The name oviraptor means “egg stealer,” and it was given because remains of this carnivorous dinosaur were found along with fossil eggs presumed to belong to a small ceratopsian, Protoceratops, which lay nearby. Recent discoveries in Mongolia of oviraptorids sitting in birdlike positions on nests of eggs formerly thought to belong to Protoceratops reveal that the parentage was misplaced and that oviraptorids, like their bird relatives, apparently tended their young. Therizinosaurids, or segnosaurs, were medium-size Asian theropods known only from a few examples. The mouth had bladelike teeth at the back but apparently no teeth at the front. The pelvis differed markedly from the normal saurischian design. They are very inadequately understood but seem to have been unlike all other theropods. Caenagnathids are not well known either but appear to have had rounded jaws that, lacking or bearing reduced teeth, are sometimes mistaken for the jaws of birds.
 The maniraptorans comprise birds, dromaeosaurs (dromaeosaur), and troodontids. Dromaeosaurs were medium-size predators with long, grasping arms and hands, moderately long legs, and a specialized stiffened tail that could be used for active balance control. Their feet bore large talons on one toe that were evidently used for raking and slicing prey. A famous discovery known as the “fighting dinosaurs of Mongolia” features a small dromaeosaur, Velociraptor, locked in petrified combat with a small protoceratopsian. The hands of the dromaeosaur are grasping the beaked dinosaur's frill, and the foot talons are apparently lodged in its throat. The best-known examples are Deinonychus of North America and Velociraptor of Asia.
The maniraptorans comprise birds, dromaeosaurs (dromaeosaur), and troodontids. Dromaeosaurs were medium-size predators with long, grasping arms and hands, moderately long legs, and a specialized stiffened tail that could be used for active balance control. Their feet bore large talons on one toe that were evidently used for raking and slicing prey. A famous discovery known as the “fighting dinosaurs of Mongolia” features a small dromaeosaur, Velociraptor, locked in petrified combat with a small protoceratopsian. The hands of the dromaeosaur are grasping the beaked dinosaur's frill, and the foot talons are apparently lodged in its throat. The best-known examples are Deinonychus of North America and Velociraptor of Asia. Ornithischia (ornithischian)
 The Ornithischia were all plant eaters, as far as is known. In addition to a common pelvic structure, they share a number of other unique features, including a bone that joined the two lower jaws and distinctive leaf-shaped teeth crenulated along the upper edges. They had at least one palpebral, or “eyelid,” bone, reduced skull openings near the eyes and in the lower jaw (antorbital and mandibular), five or more sacral (sacrum) vertebrae, and a pubis whose main shaft points backward and down, parallel to the ischium. The earliest and most basal form is the incompletely known Pisanosaurus, from the Late Triassic of Argentina. Some teeth and footprints and some fragmentary skeletal material of ornithischians are known from Late Triassic sediments, but it is only in the Early Jurassic that they become well known. Basal Jurassic forms include Lesothosaurus and other fabrosaurids, small animals that are the best-known basal ornithischians. They have the ornithischian features mentioned above but few specializations beyond these. Otherwise, the two main ornithischian lineages are the Cerapoda and Thyreophora.
The Ornithischia were all plant eaters, as far as is known. In addition to a common pelvic structure, they share a number of other unique features, including a bone that joined the two lower jaws and distinctive leaf-shaped teeth crenulated along the upper edges. They had at least one palpebral, or “eyelid,” bone, reduced skull openings near the eyes and in the lower jaw (antorbital and mandibular), five or more sacral (sacrum) vertebrae, and a pubis whose main shaft points backward and down, parallel to the ischium. The earliest and most basal form is the incompletely known Pisanosaurus, from the Late Triassic of Argentina. Some teeth and footprints and some fragmentary skeletal material of ornithischians are known from Late Triassic sediments, but it is only in the Early Jurassic that they become well known. Basal Jurassic forms include Lesothosaurus and other fabrosaurids, small animals that are the best-known basal ornithischians. They have the ornithischian features mentioned above but few specializations beyond these. Otherwise, the two main ornithischian lineages are the Cerapoda and Thyreophora.Cerapoda
Cerapoda is divided into three groups: Ornithopoda, Pachycephalosauria, and Ceratopsia. The latter two are sometimes grouped together as Marginocephalia because they share a few features, including a bony shelf on the back of the skull.
Ornithopoda (ornithopod)
Ornithopods include heterodontosaurs, known from southern Africa; the slightly larger hypsilophodontids, about three metres in length; the much larger iguanodontids, about nine metres long, mostly from North America and Europe; and the large duck-billed hadrosaurs of North America and Eurasia. In all these forms, the front teeth are set slightly lower than the cheek teeth; the jaw joint is set lower than where the teeth meet in the jaws (the occlusal plane); and the nasal bone is excluded by a separate bone (the premaxilla) from contacting the upper jaw (maxilla).
The postcranial anatomy of the ornithopods reflects the bipedal ancestry of the group, but the giant hadrosaurs and some iguanodontids may have been as comfortable on four legs as on two, especially while feeding on low vegetation. All members had hind legs that were much longer and sturdier than their forelegs. The thighbone (femur) was nearly always shorter than the shinbones (tibia and fibula), especially in all but the largest forms, and it usually bore a prominent process, called the fourth trochanter, just above mid-length for the attachment of the retractor, or walking, muscles. The pelvis was expanded, usually with an elongated and broad blade of the ilium for the attachment of the protractor, or recovery, leg muscles. The pubis, as in all ornithischians, had migrated backward to lie parallel to the ischium, as described above; but in all but the most basal forms, a new prepubic process began to grow forward from the pubis, eventually reaching far in front of the forward edge of the ilium and becoming expanded into a paddlelike shape in hadrosaurs. It is generally thought that this process supported abdominal muscles and connective tissues of internal organs, but little is demonstrably known. The tail was long and sometimes quite deep and flat-sided. The vertebral spines of the tail and trunk region were reinforced by a rhomboidal latticework of bony (ossified) tendons running in criss-cross fashion between adjacent spines. They suggest a certain degree of stiffening of the tail and backbone, which were balanced over the massive hips.
Ornithopod feet were modified from the primitive five-toed pattern in a way that resembled similar modifications in theropod feet. The three middle toes served as the functional foot; the inside toe was shortened and often held off the ground, and the outside toe was greatly reduced or absent altogether. The resemblance to theropod feet is so strong that the footprints of the two groups are easily confused, especially if poorly preserved. The toes of all but the most basal ornithopods terminated in broad, almost hooflike bones, especially in the duckbills, as opposed to the sharp claws of theropods, and this is one way to distinguish their footprints. The hand reflected the primitive five-digit design, and, as was generally true in archosaurs (archosaur), the fourth and fifth digits were shorter than the other three, with the third being longest. In iguanodontids and hadrosaurs, the fingers ended in broad, blunt bones rather than in claws, much like the toes. It is thought that these middle fingers and toes were covered by blunt, hooflike structures. In the duckbills the fingers apparently were encased in a mittenlike structure that could have broadened the hand for better support of the animal's weight on soft ground.
The Ornithopoda differ from one another mainly in the structure of their skulls, their jaws and teeth, their hands and feet, and their pelvises. Ornithopods constitute an excellent case study in evolution because, as the various lineages arise and die out from the latest Triassic to the latest Cretaceous, trends in size, complications and elaborations of teeth and chewing mechanisms, adaptations for quadrupedal posture in some forms, and other changes emerge clearly from their phylogenetic patterns.
In the fabrosaurids the teeth were simple leaf-shaped, laterally compressed elements arranged in a single front-to-back row in each jaw. They were not set in from the outer cheek surface as in most ornithopods. Small incisor-like teeth were borne on the premaxillary bones above, but (as always) no teeth were present on the predentary below. One pair of incisors had been lost. The lower jaw had no coronoid process for large muscle attachment, and the upper temporal opening (the jaw muscle site), like the mandibular opening, was relatively smaller than in theropods and other archosaurs. Upper and lower teeth alternated in position when the jaw was closed; they did not occlude directly.
In heterodontosaurs the cheek teeth were crowded together into long rows and set inward slightly from the outer cheek surface. The inset, which persisted through all later ornithopods, has been interpreted to suggest the presence of cheeks that may have held plant food in the mouth for further processing by the cheek teeth. They occluded directly to form distinct chisel-like cutting edges with a self-sharpening mechanism maintained by hard enamel on the outer side of the upper teeth and the inner side of the lower. There were prominent upper and lower tusklike teeth at the front of the mouth (the upper set in the premaxillary bones, the lower on the dentary bones). At least two pairs of incisors seem to have been retained. Certain features of the skull suggest much larger jaw muscles in heterodontosaurs than in the fabrosaurids.
 The hypsilophodonts had cheek teeth arranged in tightly packed rows set well inward from the outer cheek surfaces. The teeth occluded directly, and the opposing rows formed a long shearing edge similar to that of the heterodontosaurs. There was, however, no “tusk” either above or below. The premaxillaries had small simple incisor-like teeth above the beak-covered, toothless predentary. Strong projections of bone extended up from the lower jaw toward the moderate-size upper temporal fenestrae.
The hypsilophodonts had cheek teeth arranged in tightly packed rows set well inward from the outer cheek surfaces. The teeth occluded directly, and the opposing rows formed a long shearing edge similar to that of the heterodontosaurs. There was, however, no “tusk” either above or below. The premaxillaries had small simple incisor-like teeth above the beak-covered, toothless predentary. Strong projections of bone extended up from the lower jaw toward the moderate-size upper temporal fenestrae.The skulls of iguanodonts accommodated still larger jaw muscles, but the cheek teeth were less regular and compacted than in the primitive ornithopods and consequently did not occlude as uniformly. Both the premaxillaries and the predentary were toothless but probably were sheathed in horny beaks.
Specialization of the teeth and jaws reached a pinnacle in the hadrosaurs, or duck-billed ornithopods. In this group a very prominent, robust projection jutted from the back of the stout lower jaw. Large chambers housing muscles were present above this process and beneath certain openings in the skull (the lateral and upper temporal fenestrae). These chambers are clear evidence of powerful jaw muscles. The dentition consisted of numerous tightly compacted teeth crowded into large grinding batteries. The battery in each jaw was composed of as many as 200 functional and replacement teeth with distinct, well-defined wear, or grinding, surfaces that resulted from very exact occlusion. As teeth were lost from the front of the jaws in iguanodontids and hadrosaurs, the snouts expanded into a bulbous shape, especially in the “duck-billed” hadrosaur, and may have been covered by a horny beak that improved feeding. These bills apparently had edges sharp enough to shred and strip leaves or needles from low shrubs and branches. pine needles have been identified in duck-billed dinosaur remains and presumably represent stomach contents.
Other interesting specializations may have assisted iguanodontids and hadrosaurs in feeding. In both groups there was a marked increase in mobility (kinesis) among the joints of the bones of the facial region. As the jaws clamped down, some cheek bones were allowed to rotate outward slightly, perhaps to cushion the stress of chewing tough foods. The hands were also unusually modified in the two groups, though in different ways. In iguanodontids the wrist bones were coalesced into a single blocky structure that was less mobile than in more primitive wrist configurations. The joints of the thumb were similarly coalesced into a single conelike spike that had limited mobility on the wrist. The middle three digits flexed in the normal way and bore broad flat, spatulate claws. The fifth digit actually had two additional joints and became somewhat opposable to the rest of the hand. It is thought that the hands may have been adapted to grasp and strip vegetation, and the spikelike thumb has been suggested to have been an effective weapon against predators. These features were more or less continued in hadrosaurs, except in this group the blocky wrist was reduced and the thumb was lost completely.
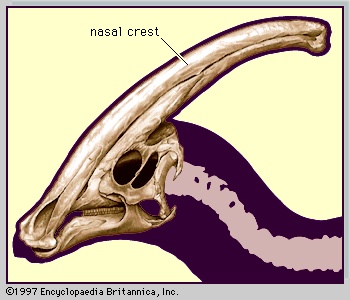 Some varieties of hadrosaurs are also noted for the peculiar crests and projections on the top of the head. These structures were expansions of the skull composed almost entirely of the nasal bones. In genera such as Corythosaurus, Lambeosaurus, Parasaurolophus (and a few others), the crests were hollow, containing a series of middle and outer chambers that formed a convoluted passage from the nostrils to the trachea. Except for passing air along to the lungs, the function of these crests is not widely agreed upon. Sound production (honking), an improved sense of smell, and a visually conspicuous ornament for species recognition are some suggestions. Because these animals are no longer considered to have been amphibious, ideas such as snorkeling and extra air storage space have generally been discarded. Besides, the crests had no opening at their ends and consequently would not have been able to work as snorkels; even the largest crests held only an estimated 2 percent of the volume of the lungs, hardly enough to justify the construction of such an elaborate structure.
Some varieties of hadrosaurs are also noted for the peculiar crests and projections on the top of the head. These structures were expansions of the skull composed almost entirely of the nasal bones. In genera such as Corythosaurus, Lambeosaurus, Parasaurolophus (and a few others), the crests were hollow, containing a series of middle and outer chambers that formed a convoluted passage from the nostrils to the trachea. Except for passing air along to the lungs, the function of these crests is not widely agreed upon. Sound production (honking), an improved sense of smell, and a visually conspicuous ornament for species recognition are some suggestions. Because these animals are no longer considered to have been amphibious, ideas such as snorkeling and extra air storage space have generally been discarded. Besides, the crests had no opening at their ends and consequently would not have been able to work as snorkels; even the largest crests held only an estimated 2 percent of the volume of the lungs, hardly enough to justify the construction of such an elaborate structure.Pachycephalosauria
In important respects the pachycephalosaurs conformed to the basic ornithopod body plan, and there is some evidence that pachycephalosaurs actually evolved from (and are therefore members of) ornithopods, perhaps similar to hypsilophodontids. All of them appear to have been bipedal. They bore the typical ornithopod ossified tendons along the back, and they had simple leaf-shaped teeth, although the teeth were enameled on both sides. The ornithischian type of pelvis was present, but a portion of the ischium was not.
 The pachycephalosaurs are known as domeheads because of their most distinctive feature—a marked thickening of the frontoparietal (forehead) bones of the skull. The thickness of bone was much greater than might be expected in animals of their size. The suggestion has been made that this forehead swelling served as protection against the impact of the type of head-butting activities seen today in animals such as bighorn sheep, but microscopic studies of the bone structure of these thick domes suggest that they are poorly designed to divert stresses away from the braincase. Also, the great variety of pachycephalosaur domes—from thin, flat skull tops to pointed ridges with large spikes and knobs facing down and back—suggests no single function in defense or combat.
The pachycephalosaurs are known as domeheads because of their most distinctive feature—a marked thickening of the frontoparietal (forehead) bones of the skull. The thickness of bone was much greater than might be expected in animals of their size. The suggestion has been made that this forehead swelling served as protection against the impact of the type of head-butting activities seen today in animals such as bighorn sheep, but microscopic studies of the bone structure of these thick domes suggest that they are poorly designed to divert stresses away from the braincase. Also, the great variety of pachycephalosaur domes—from thin, flat skull tops to pointed ridges with large spikes and knobs facing down and back—suggests no single function in defense or combat.Stegoceras and Pachycephalosaurus of the North American Cretaceous were, respectively, the smallest and largest members of the group, the former attaining a length of about 2.5 metres (8 feet) and the latter twice that. Pachycephalosaurs are known almost entirely from the Late Cretaceous (although Yaverlandia is from the Early Cretaceous) and have been found in North America and Asia. They are generally rare and still are relatively poorly known among dinosaur groups.
Ceratopsia
The first ceratopsian (“horn-faced”) dinosaur remains were found in the 1870s by the American paleontologist Edward D. Cope (Cope, Edward Drinker), who named the animal Agathaumus, but the material was so fragmentary that its unusual design was not at once recognized. The first inkling that there had been horned dinosaurs did not emerge until the late 1880s with the discovery of a large horn core, first mistaken for that of a bison. Shortly afterward, dozens of large skulls with horns were found—the first of many specimens of Triceratops.
Ceratopsians first appeared in the modest form of psittacosaurids, or parrot-reptiles, in the Early Cretaceous and survived to the “great extinction” at the end of the Cretaceous Period. Triceratops, together with Tyrannosaurus, was one of the very last of all known Mesozoic Era dinosaurs in North America, where the fossil record of the latest Cretaceous is best known. Ceratopsians had a peculiar geographic distribution: the earliest and most primitive kinds, such as Psittacosaurus, are known only from Asia—Mongolia and China, specifically. Protoceratops and its relatives are known from both Asia and North America. All the advanced ceratopsids (chasmosaurines and centrosaurines), with the exception of a few fragmentary and doubtful specimens, have been found only in North America.
 Ceratopsians ranged in size from relatively small animals the size of a dog to the nearly 9-metre- (30-foot-) long, four- to five-ton Triceratops. Although commonly compared to the modern rhinoceros, Triceratops grew to a weight and bulk several times that of the largest living rhinoceros, and its behaviour probably was correspondingly different. The most distinctive feature of nearly all members of the group was the horns on the head, hence the name ceratops. Correlated with the various arrays of head horns in the different taxa was the unusually large size of ceratopsian heads. Great bony growths extended from the back of the skull, reaching well over the neck and shoulders. This neck shield, or frill, resulted in the longest head that ever adorned any land animal; the length of the Torosaurus skull was almost 3 metres (10 feet), longer than a whole adult Protoceratops.
Ceratopsians ranged in size from relatively small animals the size of a dog to the nearly 9-metre- (30-foot-) long, four- to five-ton Triceratops. Although commonly compared to the modern rhinoceros, Triceratops grew to a weight and bulk several times that of the largest living rhinoceros, and its behaviour probably was correspondingly different. The most distinctive feature of nearly all members of the group was the horns on the head, hence the name ceratops. Correlated with the various arrays of head horns in the different taxa was the unusually large size of ceratopsian heads. Great bony growths extended from the back of the skull, reaching well over the neck and shoulders. This neck shield, or frill, resulted in the longest head that ever adorned any land animal; the length of the Torosaurus skull was almost 3 metres (10 feet), longer than a whole adult Protoceratops.Several hypotheses have been proposed to explain this frill structure: a protective shield to cover the neck region, an attachment site of greatly enlarged jaw muscles, an attachment site of powerful neck muscles for wielding the head horns, or a sort of ornament to present a huge, frightening head-on profile to potential attackers. The most unusual thought is that the structure was none of these, but rather acted as a giant heat-control apparatus, with its entire upper surface covered in a vast network of blood vessels pulsing with overheated blood or absorbing solar heat.
Most of these hypotheses are difficult to test. One important fact to keep in mind was that the frill was little more than a frame of bone, sometimes ornamented with knobs and spikes around large openings behind and above the skull. An exception to this pattern was Triceratops, which had a solid and relatively short frill, but Triceratops is so well known that its frill is often mistakenly considered typical of ceratopsians. The open frill of other ceratopsians would have provided only poor protection for the neck region and only a modest area of attachment for jaw or neck muscles. If skin and soft tissues spanned the area framed by the bony frill, it would have created a formidable presence when the head was lowered in threatening display. Such a large structure would naturally have absorbed and reflected sunlight that warmed the tissue and its internal blood vessels, but it is questionable whether this was an important or necessary function of the frill, since other dinosaurs do not have similar structures.
 The Ceratopsia are divided into groups that mirror their evolutionary trends through time: the primitive psittacosaurids, such as Psittacosaurus; the protoceratopsids, including Protoceratops of Asia and Leptoceratops of North America; and the ceratopsids, encompassing all the advanced and better-known kinds such as the chasmosaurines Triceratops and Torosaurus as well as the centrosaurines such as Centrosaurus (or Monoclonius)—all from North America.
The Ceratopsia are divided into groups that mirror their evolutionary trends through time: the primitive psittacosaurids, such as Psittacosaurus; the protoceratopsids, including Protoceratops of Asia and Leptoceratops of North America; and the ceratopsids, encompassing all the advanced and better-known kinds such as the chasmosaurines Triceratops and Torosaurus as well as the centrosaurines such as Centrosaurus (or Monoclonius)—all from North America.Like the pachycephalosaurs, the most basal ceratopsians, such as Psittacosaurus, look much like typical ornithopods, largely because of their relatively long hind limbs and short front limbs (probably resulting in bipedal stance and locomotion) and the persistence of upper front teeth and a fairly unspecialized pelvis. Resembling ornithopods in body form, Psittacosaurus had a shorter neck and tail and was much smaller (only 2 metres 【6.5 feet】 long) than the most advanced ornithopods such as the iguanodonts and hadrosaurs. Psittacosaurus, however, possessed a beak, the beginnings of a characteristic neck frill at the back of the skull, and teeth that prefigured those of the more advanced ceratopsians. It is also recognized diagnostically as a ceratopsian by the presence of a unique bone called the rostral, a toothless upper beak bone that opposed the lower predentary found in all ornithischians.
 The best-known of the protoceratopsids (Protoceratops) is the genus Protoceratops. Dozens of skeletal specimens, ranging from near hatchlings to full-size adults, have been found and studied. This rare treasure, the first to include very young individuals unmistakably associated with mature individuals, was the result of the series of American Museum of Natural History expeditions in the 1920s to the Gobi Desert (Gobi) of Mongolia. Their collection provided the first valid growth series of any dinosaur. Their discovery of several nests of eggs loosely associated with Protoceratops skeletons was the first finding of eggs that were unquestionably dinosaurian; originally attributed to Protoceratops, the eggs only recently were correctly attributed to the theropod Oviraptor (as noted in the section Tetanurae (dinosaur)).
The best-known of the protoceratopsids (Protoceratops) is the genus Protoceratops. Dozens of skeletal specimens, ranging from near hatchlings to full-size adults, have been found and studied. This rare treasure, the first to include very young individuals unmistakably associated with mature individuals, was the result of the series of American Museum of Natural History expeditions in the 1920s to the Gobi Desert (Gobi) of Mongolia. Their collection provided the first valid growth series of any dinosaur. Their discovery of several nests of eggs loosely associated with Protoceratops skeletons was the first finding of eggs that were unquestionably dinosaurian; originally attributed to Protoceratops, the eggs only recently were correctly attributed to the theropod Oviraptor (as noted in the section Tetanurae (dinosaur)).The skeletal anatomy of the protoceratopsids foreshadowed that of the more advanced ceratopsids. The ceratopsian skull was disproportionately large for the rest of the animal, constituting about one-fifth of the total body length in Protoceratops and at least one-third in Torosaurus. The head frill of Protoceratops was a modest backward extension of two cranial arches, but it became the enormous fan-shaped ornament of later forms. Protoceratops also displayed a short but stout horn on the snout due to development of the nasal bones; this too was a precursor of the prominent nasal horns of ceratopsids such as Centrosaurus, Chasmosaurus, Styracosaurus, Torosaurus, and Triceratops. The last two genera evolved two additional larger horns above the eyes. These horns undoubtedly were covered by horny sheaths or soft tissue, as is evidenced by impressions on them of superficial vascular channels for nourishing blood vessels. These advanced ceratopsids are sometimes divided into centrosaurines, which had a prominent nose horn but small or absent eye horns, and chasmosaurines, which had larger eye horns but reduced nose horns.
Ceratopsian jaws were highly specialized. The lower jaw was massive and solid to support a large battery of teeth similar to those of the duckbills. The lower jawbones were joined at the front and capped by a stout beak formed of the toothless predentary bone. This structure itself must have been covered by a sharp, horny, turtlelike beak. Continuous dental surfaces extended over the rear two-thirds of the jaw. The tooth batteries, however, differed from those of the hadrosaurs in forming long, vertical slicing surfaces as upper and lower batteries met, operating much like self-sharpening shears.
As in the hadrosaurs, each dental battery consisted of about two dozen or more tooth positions compressed together into a single large block. At each tooth position there was one functional, or occluding, tooth (the duckbills had two or three) along with several more unerupted replacement teeth beneath. (All toothed vertebrates, living and extinct, except mammals, have a lifelong supply of replacement teeth.) The suggestion is that they fed on something exceedingly tough and fibrous, such as the fronds of palms or cycads, both of which were plentiful during late Mesozoic times.
With the exception of the bipedal Psittacosaurus, and perhaps the facultatively bipedal protoceratopsids, all ceratopsians were obligate quadrupeds with a heavy, ponderous build. The leg bones were stout and the legs themselves muscular; the feet were semiplantigrade for graviportal stance and progression; and all the toes ended in “hooves” rather than claws. As in most other four-legged animals, the rear legs were significantly longer than the front legs (which again suggests their bipedal ancestry). The hind legs were positioned directly beneath the hip sockets and held almost straight and vertical. The front legs, on the other hand, projected out to each side from the shoulder sockets in a “push-up” position. Consequently, the head was carried low and close to the ground. This mixed posture was perhaps related to the large horned head and its role in combat, the bent forelegs providing a wide stance and stable base for directing the horns at an opponent and resisting attack.
The first four neck vertebrae of ceratopsians were fused (co-ossified), presumably to support the massive skull. The first joint of the neck was unusual in that the bone at the base of the skull formed a nearly perfect sphere that fit into a cuplike socket of the fused neck vertebrae. Such an arrangement would seem to have provided solid connections along with maximum freedom of the head to pivot in any direction without having to turn the body. Presumably ceratopsians used their horns (horn) in an aggressive manner, but whether they used them as defense against possible predators, in rutting combat with other male ceratopsians, or in both is not so clear. Evidence of puncture wounds in some specimens suggests rutting encounters, but the fact that both sexes apparently had horns seems to indicate defense or species recognition as their primary uses.
Thyreophora
The Thyreophora consist mainly of the well-known Stegosauria, the plated dinosaurs, and Ankylosauria, the armoured dinosaurs, as well as their more basal relatives, including Scutellosaurus and Scelidosaurus. Scutellosaurus was a small bipedal dinosaur, only about a metre (3.3 feet ) in length, known from the Early Jurassic Period of Arizona, U.S. It was first classified as a fabrosaurid because of its primitive skeletal structures. However, it differed from fabrosaurids in some important respects, including the possession of small bony plates, or scutes, of various shapes along the back and sides of its body. These scutes are also found in the slightly larger Scelidosaurus, which was up to three metres in length and quadrupedal; this dinosaur is known from the Early Jurassic of England and Arizona.
In the Middle and Late Jurassic, the first stegosaurs and ankylosaurs appeared. Like the previously described forms, they are distinguished by bony scutes. Scutes are maintained and elaborated all over the body in ankylosaurs but are reduced to a series of plates and spikes along the backbone in stegosaurs, though their basic structure remains the same in both groups. Thyreophorans also have low, flat skulls, simple S-shaped tooth rows with small leaf-shaped tooth crowns, and spout-shaped snouts.
Stegosauria
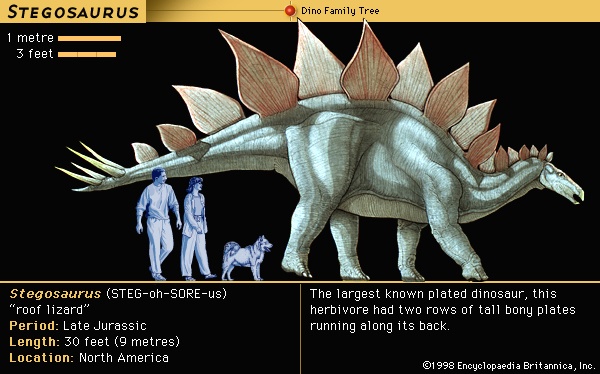 With their unique bony back plates, the stegosaurs are very distinctive. Relatively few specimens have been found, but they were widespread, with remains being found in North America, Africa, Europe, and Asia. Stegosaurian remains have appeared in Early Jurassic to Early Cretaceous strata. The most familiar genus is Stegosaurus, found in the Morrison Formation (Late Jurassic) of western North America. Stegosaurus was 3.7 metres (12 feet) in height and 9 metres in length, probably weighed two tons, and had a broad, deep body. Not all varieties of the Stegosauria were this large; for example, Kentrosaurus, from eastern Africa, was less than 2 metres high and 3.5 metres long.
With their unique bony back plates, the stegosaurs are very distinctive. Relatively few specimens have been found, but they were widespread, with remains being found in North America, Africa, Europe, and Asia. Stegosaurian remains have appeared in Early Jurassic to Early Cretaceous strata. The most familiar genus is Stegosaurus, found in the Morrison Formation (Late Jurassic) of western North America. Stegosaurus was 3.7 metres (12 feet) in height and 9 metres in length, probably weighed two tons, and had a broad, deep body. Not all varieties of the Stegosauria were this large; for example, Kentrosaurus, from eastern Africa, was less than 2 metres high and 3.5 metres long.All stegosaurs were graviportal and undoubtedly quadrupedal, although the massive legs were of greatly disparate lengths—the hind legs being more than twice the length of the forelegs. Whatever walking and running skills were possessed by the stegosaurs, their limb proportions must have made these movements extremely slow. The humerus of the upper arm was longer than the bones of the forearm, the femur much longer than the shinbones, and certain bones of the feet very short, which means that the stride must have been short. In addition, the feet were graviportal in design and showed no adaptations for running.
The stegosaurian skull was notably small, long, low, and narrow, with little space for sizable jaw muscles. The weakly developed dentition consisted of small, laterally compressed, leaf-shaped teeth arranged in short, straight rows. This combination of features seems odd in comparison with the large, bulky body. The weak dentition suggests that the food eaten must have required little preparation by the teeth and yet provided adequate nourishment. Perhaps the digestive tract contained fermenting bacteria capable of breaking down the cellulose-rich Jurassic plant tissues. Digestion may also have been assisted by a crop or gizzard full of pulverizing stomach stones (gastroliths), though none has yet been discovered in stegosaurian specimens. A collection of disklike bones is found in the throat region of Stegosaurus, but these are likely to have been embedded in the skin, not used in the gut. Even so, it is still difficult to understand how these animals, with such small and poorly equipped mouths, could have fed themselves adequately to sustain their great bulk. The same problem has been encountered in speculations about the feeding habits of sauropods.
The most distinctive stegosaurian feature was the double row of large diamond-shaped bony plates on the back. A controversy as to their purpose and how they were arranged has raged ever since the first Stegosaurus specimen was collected (1877, Colorado, U.S.). The evidence and a general consensus argue in favour of the traditional idea that the plates projected upward and were set in two staggered (alternating) rows on either side of the backbone. In other stegosaurs, such as Kentrosaurus, the plates are more symmetrical and may have been arranged side by side. The suggestion that the plates did not project above the back at all, but lay flat to form flank armour, has been rejected on the basis of studies of the microstructure of the bone of the plates, in which attachment fibres are embedded in a manner consistent with an upright position. In Stegosaurus itself, the end of the tail bore at least two pairs of long bony spikes, which suggests some sort of defensive role for the tail but not necessarily for the back plates. However, other stegosaurs, such as Kentrosaurus, had relatively small plates along the front half of the spine and spikes along the back half of the spine and the tail.
The discovery in 1976 that the bony plates of Stegosaurus were highly vascularized led to the suggestion that these “fins” functioned as cooling vanes to dissipate excess body heat in much the same way that the ears of elephants (elephant) do. The staggered arrangement in parallel rows might have maximized the area of cooling surface by minimizing any downwind “breeze shadow” that would have resulted from a paired configuration. Asymmetry is a bizarre anatomic condition, and, right or wrong, this certainly is an imaginative explanation of its presence in this animal. No other stegosaur, however, had such a peculiar feature. Rather, all other taxa had a variety of paired body spikes that seem best explained as passive defense or display adaptations rather than cooling mechanisms.
Ankylosauria
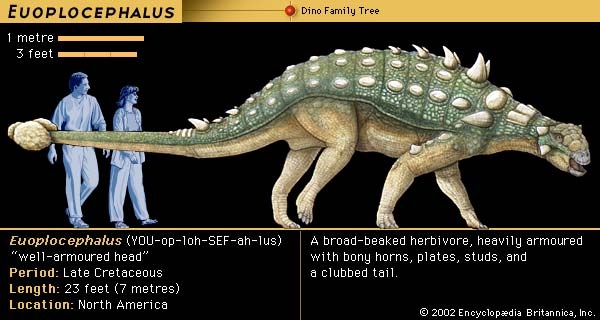 The ankylosaurs are known from the Late Jurassic and Cretaceous periods. They are called “armoured dinosaurs” for their extensive mosaic of small and large interlocking bony plates that completely encased the back and flanks. Most ankylosaurs, such as Euoplocephalus, Nodosaurus, and Palaeoscincus, were relatively low and broad in body form and walked close to the ground on short, stocky legs in a quadrupedal stance. As in stegosaurs, the hind legs were longer than the front legs, but they were not as disproportionate as those of Stegosaurus. Like the stegosaurs, however, their limbs were stout and columnar, the thighbone and upper arm were longer than the shin and forearm, and the metapodials were stubby. These features point to a slow, graviportal mode of locomotion. The feet were semiplantigrade and possibly supported from beneath by pads of cartilage. The bones at the ends of the digits (terminal phalanges) were broad and hooflike rather than clawlike.
The ankylosaurs are known from the Late Jurassic and Cretaceous periods. They are called “armoured dinosaurs” for their extensive mosaic of small and large interlocking bony plates that completely encased the back and flanks. Most ankylosaurs, such as Euoplocephalus, Nodosaurus, and Palaeoscincus, were relatively low and broad in body form and walked close to the ground on short, stocky legs in a quadrupedal stance. As in stegosaurs, the hind legs were longer than the front legs, but they were not as disproportionate as those of Stegosaurus. Like the stegosaurs, however, their limbs were stout and columnar, the thighbone and upper arm were longer than the shin and forearm, and the metapodials were stubby. These features point to a slow, graviportal mode of locomotion. The feet were semiplantigrade and possibly supported from beneath by pads of cartilage. The bones at the ends of the digits (terminal phalanges) were broad and hooflike rather than clawlike.The ankylosaur skull was low, broad, and boxlike, with dermal scutes (osteoderms) that were often fused to the underlying skull bones. In Euoplocephalus even the eyelid seems to have developed a protective bony covering. The jaws were weak, with a very small predentary and no significant projections of bone for jaw muscle attachment. The small jaw muscle chamber was largely covered by dermal bones rather than having openings. The teeth were small, loosely spaced, leaf-shaped structures reminiscent of the earliest primitive ornithischian teeth. All taxa had very few teeth in either jaw, in marked contrast to the highly specialized, numerous teeth of other ornithischians. These features of the jaws and teeth lead to the impression that the animals must have fed on some sort of soft, pulpy plant food.
Apparently neither very diverse nor abundant, the ankylosaurs are known only from North America, Europe, and Asia. They are divided into the more basal Nodosauridae and the more advanced Ankylosauridae, which may have evolved from nodosaurs. The most conspicuous difference between the two groups is the presence of a massive bony club at the end of the tail in the advanced ankylosaurs; no such tail structure is present in the nodosaurs. The patterns of the armour also generally differ between the two groups, and ankylosaurids tend to have even broader, more bone-encrusted skulls than did the nodosaurs.
Additional Reading
General works
Philip J. Currie and Kevin Padian (eds.), Encyclopedia of Dinosaurs (1997), comprises articles on topics related to dinosaur taxonomy, biology, and evolution as well as important paleontological sites and exhibits worldwide. James O. Farlow and M.K. Brett-Surman (eds.), The Complete Dinosaur (1997), emphasizes aspects of various groups of dinosaurs and their biology. David Norman and John Sibbick, The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom (1985, reissued 1998), provides a well-written and lavishly illustrated treatment that is excellent for the specialist and nonspecialist alike.Advanced textbooks on vertebrate evolution and paleontology include Michael J. Benton, Vertebrate Palaeontology, 2nd ed. (1997, reissued 2000); and Robert L. Carroll, Vertebrate Paleontology and Evolution (1988). David B. Weishampel, Peter Dodson, and Halszka Osmólska (eds.), The Dinosauria (1990), primarily contains extensive reviews of the major taxonomic groups, defining them via anatomic descriptions and drawings while also supplying fossil-site information.
The search for dinosaurs
Louie Psihoyos and John Knoebber, Hunting Dinosaurs (1994), assembles an impressive photographic record of the discoveries and the people responsible for them. Philippe Taquet, Dinosaur Impressions: Postcards from a Paleontologist (1998; originally published in French, 1994), vividly traces one paleontologist's travels throughout the world over a period of 30 years. Edwin H. Colbert, Dinosaurs: Their Discovery and Their World (1961), a landmark treatment of the subject by a world authority of the period, includes extensive photographic and line-drawing coverage, and Men and Dinosaurs: The Search in Field and Laboratory (1968, reissued 1971), provides a thorough illustrated history of the discovery, collection, and study of dinosaurs. John R. Horner and James Gorman, Digging Dinosaurs (1988, reprinted 1995), is a fascinating account of the search for and collecting of dinosaur eggs and nests as told by the discoverers. John H. Ostrom and John S. McIntosh, Marsh's Dinosaurs: The Collections from Como Bluff (1966, reissued 1999), is illustrated for technical professionals and contains a historical study of one of the most famous dinosaur localities.For specific information about the origin of birds from theropod dinosaurs, Lowell Dingus and Timothy Rowe, The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds (1998), is an excellent reference source. Kevin Padian and Luis M. Chiappe, “The Origin of Birds and Their Flight,” Scientific American, 278(2):38–47 (February 1998), provides a brief account of the evidence; and Jennifer Ackerman, “Dinosaurs Take Wing,” National Geographic, 194(1):74–99 (July 1998), furnishes additional information and graphics.
Natural history
John R. Horner and Edwin Dobb, Dinosaur Lives: Unearthing an Evolutionary Saga (1997), explores developments in the understanding of dinosaurian paleobiology. Peter Dodson, The Horned Dinosaurs: A Natural History (1996), presents a case history of the ceratopsians, an important group of dinosaurs. Kenneth Carpenter, Karl F. Hirsch, and John R. Horner (eds.), Dinosaur Eggs and Babies (1994, reissued 1996), a technical multiauthor work, reviews many aspects of dinosaur reproductive biology.
Extinction
The most authoritative account of the Late Cretaceous extinctions is J. David Archibald, Dinosaur Extinction and the End of an Era: What the Fossils Say (1996), a masterful book. Walter Alvarez, T. rex and the Crater of Doom (1998), offers a somewhat different point of view.
- Desbordes-Valmore, Marceline
- Desborough, John
- descamisado
- descant
- Descartes, René
- descent
- Deschamps, Eustache
- Deschamps, Émile
- Deschanel, Paul
- descloizite
- descort
- Dese
- Deseado River
- desensitization
- Deseret News, The
- desert
- Desert cultures
- desert dormouse
- Desert Fathers
- desertification
- desert palace
- desert pavement
- Desert Rats
- Deserts of the world
- desert varnish