temperate forest
ecology
Introduction
 vegetation type with a more or less continuous canopy of broad-leaved trees. Such forests occur between approximately 25° and 50° latitude in both hemispheres (seeFigure 1-->). Toward the polar regions they grade into boreal forests, which are dominated by evergreen conifers, so that mixed forests containing both deciduous and coniferous trees occupy intermediate areas. Temperate forests usually are classified into two main groups: deciduous and evergreen.
vegetation type with a more or less continuous canopy of broad-leaved trees. Such forests occur between approximately 25° and 50° latitude in both hemispheres (seeFigure 1-->). Toward the polar regions they grade into boreal forests, which are dominated by evergreen conifers, so that mixed forests containing both deciduous and coniferous trees occupy intermediate areas. Temperate forests usually are classified into two main groups: deciduous and evergreen.Deciduous forests are found in regions of the Northern Hemisphere that have moist, warm summers and frosty winters—primarily eastern North America, eastern Asia, and western Europe. In contrast, evergreen forests—excepting boreal forests, which are covered in boreal forest—typically grow in areas with mild, nearly frost-free winters. They fall into two subcategories—broad-leaved forests and sclerophyllous forests. (Sclerophyllous vegetation has small, hard, thick leaves.) The former grow in regions that have reliably high, year-round rainfall; the latter occur in areas with lower, more erratic rainfall. Broad-leaved forests dominate the natural vegetation of New Zealand; they are significantly represented in South America, eastern Australia, southern China, Korea, and Japan; and they occur in less well-developed form in small areas of southeastern North America and southern Africa. Sclerophyllous forests occur particularly in Australia and in the Mediterranean region.
Origin
Temperate forests originated during the period of cooling of world climate that began at the start of the Cenozoic Era (66.4 million years ago to the present). As global climates cooled, climatic gradients steepened with increasing latitude, and areas with a hot, wet climate became restricted to equatorial regions. At temperate latitudes, climates became progressively cooler, drier, and more seasonal. Many plant lineages that were unable to adapt to new conditions became extinct, but others evolved in response to the climatic changes, eventually dominating the new temperate forests. In areas that differed least from the previously tropical environments—where temperate evergreen forests now grow—the greatest numbers of plant and animal species survived in forms most similar to those of their tropical ancestors. Where conditions remained relatively moist but temperatures dropped in winter, deciduous trees evolved from evergreen rainforest ancestors. In areas that became much more dry—though not to the extent that tree development was inhibited and only scrubland or desert environments were favoured—sclerophyllous trees evolved.
During the rapid climatic fluctuations of the past two million years in which conditions alternated between dry, cold glacial states—the ice ages of some northern temperate regions—and warmer, moister interglacial intervals, tree species of temperate forests had to migrate repeatedly to remain within climates suitable for their survival. Such migration was carried out by seed (seed and fruit) dispersal, and trees that were able to disperse their seeds the farthest had an advantage. In the North American (North America) and European (Europe) regions where ice-sheet development during glacial intervals was most extensive, the distances that had to be traversed were greatest, and many species simply died out. Extinctions occurred not only where migration distances were great but also where mountains or seas provided barriers to dispersal, as in southern Europe. Thus, many trees that were formerly part of the European temperate forests have become extinct in the floristically impoverished forest regions of western Europe and are restricted to small refuge areas such as the Balkans and the Caucasus. For example, buckeye (Aesculus) and sweet gum (Liquidambar) are two trees that no longer occur naturally in most parts of Europe, having disappeared during the climatic turmoil of the past two million years.
Human activities have had pronounced effects on the nature and extent of modern temperate forests. As long ago as 8,000 years, most sclerophyllous forests of the Mediterranean region had been cut over for timber or cleared (deforestation) to make space for agricultural pursuits. By 4,000 years ago in China the same process led to the removal of most broad-leaved and deciduous forests. In Europe of 500 years ago the original deciduous forests had disappeared, although they are remembered in nursery tales and other folklore as the deep, wild woods in which children and princesses became lost and in which dwarfs and wild animals lived.
The deciduous forests of North America had been cleared almost completely by the end of the 19th century. Australia and New Zealand experienced similar deforestation about the same time, although the earlier activities of pre-European peoples had had substantial impacts. The character of the Australian sclerophyllous forests changed in response to more than 38,000 years of burning by the Aboriginal people, and the range of these forests was expanded at the expense of broad-leaved forests. In New Zealand about half the forested area, which previously had covered almost the entire country, was destroyed by fire brought to the island by the Polynesian inhabitants who arrived 1,000 years before the Europeans.
Environment
winter in the temperate latitudes can present extremely stressful conditions that greatly affect the vegetation. The days are shorter and temperatures are low, so much so that in many places leaves (leaf) are unable to function for long periods and are susceptible to damage from freezing. These conditions reduce the photosynthetic (photosynthesis) activity of the trees. In regions where winter temperatures regularly fall well below the freezing point and where soil moisture and nutrients are not in short supply, many trees have evolved a type of leaf that is relatively delicate and thin with a life span of a single growing season. Because such deciduous leaves do not require a large input of chemical energy, it is not too wasteful for the plant to shed them after a single growing season.
The “throwaway” leaves of temperate deciduous trees are shed as the days shorten in autumn and are replaced by new leaves in the spring. The forests dominated by these trees, therefore, have extremely pronounced seasonal changes in appearance, function, and climate. Most trees in temperate deciduous forests follow this habit, although some evergreen species usually are scattered among them. In particular, several broad-leaved evergreen shrubs are found in the understory of temperate deciduous forests that have less delicate, longer-lasting leaves than their deciduous neighbours; these leaves have adaptations that allow them to survive the freezing winter temperatures, and they can carry out photosynthesis for more than a single summer.
In areas where milder conditions prevail, however, photosynthesis may be possible at any season without need for protective mechanisms against frost damage. In these relatively unstressful circumstances most trees may benefit from retaining their leaves throughout the year, and heavy use of resources through frequent leaf replacement is thereby avoided. In environments such as these that also have a sufficient supply of moisture, temperate broad-leaved forests are found.
In mild but drier temperate environments, moisture shortage necessitates that trees develop thickened leaves. These leaves often have a reduced surface area or they dangle pendulously from limbs, two strategies employed to slow the loss of water (transpiration). High levels of energy and nutrients are needed to produce these thick leaves, which, therefore, cannot be replaced easily at annual intervals, an added reason for sclerophyllous trees to retain their leaves throughout the year. With foliage perennially present, these trees can carry out photosynthesis whenever moisture becomes available, provided temperatures are warm enough; this characteristic is advantageous where rainfall is infrequent and unpredictable.
Soils (soil) in temperate sclerophyllous forests are frequently poor in mineral nutrients. This poverty of nutrients accounts in part for the nature of the vegetation, because the annual production of leaves or the development of a dense broad-leaved canopy requires a significant input of nutrients. In contrast, soils in regions of deciduous and broad-leaved evergreen forest are generally fertile. (The forests that occupied the best soils in most regions, however, have been cleared almost completely to make way for agriculture.) Typical temperate deciduous forest soils are mull soils, which have a high level of organic matter especially close to the surface that is well mixed with mineral matter. Variations in soil materials and fertility have a strong influence on the types of trees that will dominate the forest. For example, in northwestern Europe, the European beech (Fagus sylvatica) dominates deciduous forests on shallow soils that overlie chalk, while oak (Quercus) is dominant on deeper, clay soils. The richness of the ground flora beneath the trees generally increases with soil fertility.
Intimate associations, or mycorrhizae (mycorrhiza), between tree roots (root) and fungi (fungus) are important and occur in most tree species. Although important in all forest types, these interactions have been studied more thoroughly in temperate deciduous forests. The fungal component of this symbiotic partnership grows on or in the fine roots of trees and benefits by obtaining nutrition in the form of carbohydrates from the tree root; the tree, in turn, is better nourished because a mycorrhizal root is more efficient at absorbing dissolved mineral nutrients from the soil than is an uninfected root.
Waterlogging of soils in temperate deciduous woodlands commonly occurs in regions with higher rainfall and humidity in late winter and spring, such as the British Isles. This occurs not only because in winter rainfall is higher and evaporation lower but also because the trees, barren of foliage, transpire a minimal amount of moisture. Areas subject to waterlogging include clay-rich soils, places that have low slope angles, depressions, and spots along watercourses. These places tend to have a richer ground flora but a less luxuriant tree canopy, which consists of only a few species that are tolerant of wet soils.
The depth of tree roots in temperate deciduous forests varies, but in many instances roots are shallow, with few reaching 1 metre (3.28 feet) below the surface. In the European beech, for example, shallow lateral growth of roots predominates over the development of a deep taproot, leading to growth of a “root plate” just beneath the soil surface. This enables the tree to exploit nutrients released at the surface by litter decomposition efficiently, while avoiding deeper layers that may become waterlogged. However, trees with root plates are more prone to being blown over in gales, especially after heavy rain has made the soil more plastic.
Biota
The principal regions of deciduous forest all occur in the Northern Hemisphere and have historical connections between them. Thus, many similarities exist among their biota. The same plant and animal genera tend to occur in all regions, although different species are found in each region. However, the European (Europe) deciduous forest flora is poorer than that of eastern North America and East Asia. Many plants are common and widespread in the forests of North America and Asia, but in Europe they are present only as restricted relict populations or fossils. Examples include hickory (Carya), Magnolia, sour gum (Nyssa), and sweet gum.
Flora
Most of the areas of North American (North America) deciduous forest are dominated by oaks (oak) (several species of Quercus) and/or beech (Fagus grandifolia), with maples (maple) (Acer) and species of hickory and linden or basswood (Tilia) also widespread. Beech and basswood are rare in other North American vegetation types, but oaks, hickories, and maples are more widespread.
In addition to these widespread species, many other trees are important components of the North American deciduous forests on a local scale. The rich, mixed mesophytic (adapted to environments that are neither too dry nor too wet) forest type found north of the Appalachians includes buckeye and tulip tree (Liriodendron), while the southern floodplain forest of the Mississippi valley is made up of oaks mixed with sour gum and the evergreen conifer swamp cypress (Taxodium). Southward and eastward of the Appalachians, oaks and hickories mingle with another conifer, pine (Pinus), while east of the Great Lakes, beech mix with maples, birch (Betula), and hemlock (Tsuga). Elsewhere, ash (Fraxinus), hop hornbeam (Ostrya), poplar (Populus), elm (Ulmus), and, until its decimation by fungal infection, chestnut (Castanea), are also important. A wide range of understory shrubs and small trees includes dogwood (Cornus), holly (Ilex), Magnolia species, and serviceberry (Amelanchier).
Fragments of the North American deciduous forest also occur on mountains in Mexico and Guatemala, where many of the same trees—e.g., Fagus, Fraxinus, Juglans (walnut), Liquidambar, Quercus—occur as identical or closely related species. Commonly these trees are accompanied by an understory of evergreen shrubs of tropical affinity.
In Japan, Korea, and China, north of the evergreen broad-leaved forests there is a gradual transition to deciduous forests. In Japan deciduous forests are dominated by beech (Fagus crenata and F. japonica), oak (Quercus crispula), and maple (Acer carpinifolium and other species); other trees mingle with these, including cherries (several species of Prunus), ash, Magnolia, and, in the east, the evergreen conifer fir (Abies). The leaves of many deciduous trees in Japan, like those in North America but unlike most in Europe, turn to bright shades of red and yellow before they are shed in autumn, the maples being particularly spectacular. Below the trees a dense layer of dwarf bamboo (Sasa) commonly grows; it may be so thick that it prevents the canopy trees from regenerating from seedlings. Thus, rapid, dense regrowth by dwarf bamboo may seriously interfere with reforestation after logging. Many small flowering herbs such as Aconitum, Shortia, Mitchella, and Viola grow at ground level. Much of the area of Chinese deciduous forests is dominated by various oaks, frequently mixed with other trees including maples, alder (Alnus), ash, walnut, poplar, and many others. A varied understory includes many small trees or shrubs such as hornbeam (Carpinus), dogwood, service-tree (Sorbus), and the shrubs Acanthopanax and Aralia, which are relatives of ivy. Deciduous forests of birch fringe the oak forests at their northern and montane margins.
The dominant trees of the European deciduous forests are generally closely related to their equivalents in North America and Asia, consisting of different species of common genera. In well-drained areas, such as those on sloping ground or permeable soils, most deciduous forests are dominated by European beech or by one of a few species of oak. Beech is overwhelmingly dominant across large areas that have an oceanic climate; these regions are damper and milder than most other regions in which deciduous forests grow because of the influence of prevailing winds from the Atlantic Ocean. Eastward from Britain and western France through central Europe into Russia there is a progressive decrease in this oceanic influence. Along this gradient the forest flora changes. For example, beech, durmast oak (Quercus petraea), and European hornbeam (Carpinus betula) all reach their eastward limits in the area north of the Carpathian Mountains, while linden (Tilia cordata) is typical of a small number of deciduous forest trees that extend east beyond Moscow.
Places subject to seasonal waterlogging in European deciduous forests—and which are also nutrient-rich—often are dominated by alder (Alnus glutinosa), growing above a rich ground flora including ferns (Athyrium and Dryopteris), sedges (Carex), and forbs (Caltha and Filipendula).
The only significant temperate deciduous forests in the Southern Hemisphere occur in a small area of Chile around Valdivia, between about 36° and 41° S. Forests here are dominated by a deciduous species of beech, Nothofagus obliqua, which usually grows amid evergreen trees more typical of the broad-leaved forests bordering this area to the south.
The milder environments that support temperate evergreen forests generally lie closer to the Equator than do areas with temperate deciduous forest. They have richer biotas than the sclerophyllous or deciduous forests that grow in more stressful environments at similar latitudes, although they are less rich than the tropical rainforests where environmental stress is at a minimum throughout the year.
In the southernmost regions of Japan and Korea (Korea, South), which enjoy a warm, wet climate, the natural vegetation is evergreen broad-leaved forest dominated by oaks and their near relative Castanopsis cuspidata and by the laurel Machilus thunbergii. Camphor laurel (Cinnamomum camphora), figs (Ficus retusa), pandans (Pandanus boninensis), palms (Livistona subglobosa), and other plants that require year-round warmth also occur in the warmest places, whose vegetation is described as subtropical by some authorities and as warm temperate forest by others. Shrubs in these forests include species of Aucuba, Camellia, and Eurya, and orchids and ferns are commonly found at ground level.
Similar forests in southern China, now almost completely replaced by farmland or tree plantations, had an even richer flora, with oaks, members of the laurel family, and the tea relative Schima being prominent examples of a large diversity of trees. The remnants of these Chinese evergreen broad-leaved forests extend west to the foothills of the Himalayas, where similar forests at lower altitudes include many trees such as Alcimandra, Castanopsis, Machilus, Magnolia, and Mallotus. At higher altitudes there are more sclerophyllous forests that contain fewer species and are dominated by oaks.
In Australia a variety of temperate broad-leaved forests occur, usually as small patches in moist, sheltered, and fire-protected areas in mountainous and coastal terrain along the east coast. They are described as temperate rainforests owing to similarities with the flora, structure, and ecology of tropical rainforests that are found in similar environments to the immediate north. The warm temperate rainforests of milder, more northern areas display a high diversity of trees, including coachwood (Ceratopetalum apetalum), crab apple (Schizomeria ovata), and yellow carabeen (Sloanea woollsii). Palms are often present, as are various climbing plants and epiphytes (plants that grow on other plants but that derive moisture and nutrients from rain), although not to the extent that they occur in tropical rainforests. Ferns are typically abundant, and many large, graceful tree ferns grow there.
A similar warm temperate rainforest grows in northern parts of New Zealand; it contains a mixed broad-leaved canopy of trees such as Elaeocarpus, Metrosideros, and Weinmannia, which is frequently penetrated and overtopped by tall conifers, including the massive kauri (Agathis australis). Palms (Rhopalostylis sapida) and various lianas are often present.
Cool temperate evergreen broad-leaved forests in the southernmost areas of eastern Australia, particularly Tasmania, and in New Zealand and the southern portion of South America are usually dominated by evergreen species of beech (Nothofagus), with different species occurring in each region. Few other trees typically coexist with Nothofagus in these cool forests, which also lack climbers and vascular epiphytes, although they may have a great abundance of mosses on tree trunks, branches, and sometimes leaves. In Australia other trees that may be present include sassafras (Atherosperma moschatum), Elaeocarpus holopetalus, and leatherwood (Eucryphia lucida), while in New Zealand conifers in the plum pine family (Podocarpaceae) commonly emerge above the broad-leaved canopy, especially on sites with a history of natural disturbance such as landslides. In Chile other trees growing with Nothofagus, or in some cases forming temperate evergreen forests without it, include Eucryphia, Chilean laurel (Laurelia), and Persea, with bamboos (Chusquea) becoming abundant at some sites after forest disturbance.
South of the European deciduous forests lie areas that were occupied by temperate sclerophyllous forests before the effects of human manipulation of the environment were felt. These areas extend as a narrow ring around the coastline of the Mediterranean Sea. Typical evergreen trees are oaks (several species including the cork oak, Quercus suber) and the pistachio relative Pistacia lentiscus, commonly mixed with various deciduous species near their northern limits and with pines elsewhere. Forests similar to the Mediterranean sclerophyllous forests extend east, discontinuously, as far as the western slopes of the Himalayas, although throughout their range they have been extensively altered by human activities and nowhere are considered to be in their natural state.
Temperate sclerophyllous forests are most widespread in Australia. It is remarkable that across the major portion of a very large area, the most common trees are various species of one genus, Eucalyptus. About 500 species are known of this archetypal Australian tree, which occurs naturally in few other regions. However, gum trees, as they are often called, are well known in many regions to which they have been introduced. Two other diverse and widespread trees in these forests are Acacia and Casuarina. Many other trees, shrubs, and grasses occupy subordinate positions. A sharp boundary separates the temperate sclerophyllous forests from the tropical and temperate rainforests of moister sites in the east. This abrupt transition is maintained by fire, which prevents or reverses invasion of the sclerophyllous forest by rainforest plants. However, where fires are infrequent, especially in southern regions, rainforest vegetation can form an understory beneath the Eucalyptus canopy. To the north, temperate sclerophyllous forests grade into similar tropical savanna and sclerophyllous forests; inland they merge into shrublands and deserts.
Fauna
The fauna of temperate forests resembles the regional fauna. However, the structure of the vegetation provides similar ecological niches in all regions of the same vegetation type, so that, although different species inhabit different forests, they are of a similar type. Tree holes provide homes and nest sites for arboreal mammals and birds in most regions of temperate forest but with pronounced variations. For example, apart from bats no native mammals are found in the New Zealand forests. In Australia the arboreal mammals are all marsupials or bats, including gliders such as the greater glider (Petaurus volans) and opossums such as the common ringtail (Pseudocheirus peregrinus), which nests in holes, and the well-known koala (Phascolarctos cinerea), which is free-living and feeds mainly or entirely on young tree foliage.
In temperate forests of the Northern Hemisphere, squirrels are widespread. Local additional arboreal forms in Asian forests include monkeys, most of which are predominantly seedeaters. This feeding niche is particularly appropriate in Northern Hemisphere forests, which include more trees with large seeds, such as the acorn-producing oaks, than do their Southern Hemisphere equivalents.
Birds are less regionally distinct, with families such as those of the owl and pigeon being well-represented in almost all temperate forest regions. Nevertheless, there are still some pronounced regional variations. The tits (titmouse) (Paridae) dominate the foliage-gleaning insectivore guild in Europe, where warblers (warbler) (Sylviidae) are less varied; this situation is reversed in North America. More fundamental contrasts are apparent in Australia, where honeyeaters, which feed on nectar, and parrots, which feed on small, hard seeds, are diverse and common in the sclerophyllous forests. In the Northern Hemisphere few plants provide nectar for birds, and tree seeds are usually eaten by squirrels and pigeons.
Population and community development and structure
At mid-latitudes the sun (sunlight) never rises to the near-vertical position in the sky as it does in the tropics. In winter, when the sun appears particularly close to the horizon throughout the short period of daylight, its direct radiant energy impinges most on slopes facing the Equator—southward in the Northern Hemisphere, northward in the Southern Hemisphere. In deciduous forest regions, primarily in the Northern Hemisphere where the trees are without leaves in winter, direct sunlight bathes the forest floor. The ground surface and the vegetation on southern slopes are quickly warmed, paradoxically creating a more stressful environment for plants than exists on the shadier, cooler northern slopes. The stress is due to more extreme temperature fluctuations than the southern slope sustains, which affects the speed at which the plant tissues, frozen during the cold night, are thawed. On the southern slope the thawing is much more rapid, which is more damaging than the gradual thaw that the vegetation on the northern slopes experiences. As a result, herbaceous plants of the forest floor that retain green leaves throughout winter, such as Hepatica, Hydrophyllum, and Tiarella in eastern North America, are more common on northern slopes, while sunnier slopes become bare and brown during the winter months.
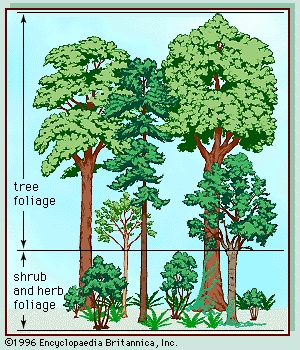 As days lengthen and temperatures warm in spring, new, green photosynthetic shoots develop rapidly from buds that formed and food reserves that were laid down in storage tissues during the previous growing season. Annuals growing from seed are rare. The most rapid development of all typically occurs in the herbaceous plants of the forest floor. These plants must take immediate advantage of the spring warmth and sunlight before the new tree foliage casts its heavy shade over the ground, drastically reducing available light energy and slowing the rate of photosynthesis. Many plants of the forest floor have underground energy-storing organs such as bulbs, corms, or fleshy rhizomes that allow them to grow rapidly and strongly in spring to gain maximum advantage from the short, warm, shade-free period. They frequently also produce strong, pointed shoots that are able to emerge above the thick layer of dead leaves that dropped from the trees of the canopy the previous autumn. Thus, the ground of temperate deciduous forests in spring typically is covered by a green carpet of foliage that often includes dramatic displays of colourful flowers before the buds on the trees above have opened. Later, after the leaves of the tree canopy have regrown, the ground cover declines. Typical low-growing plants in North American deciduous forests include species of Cypripedium, Erythronium, Hydrophyllum, Trillium, and Viola. In Europe bluebells (Hyacinthoides non-scripta), daffodils (Narcissus pseudonarcissus), and wood anemones (Anemone nemorosa), among many other herbaceous species, provide similarly spectacular spring flower displays. A vegetation profile of a typical temperate deciduous forest is shown in Figure 2-->.
As days lengthen and temperatures warm in spring, new, green photosynthetic shoots develop rapidly from buds that formed and food reserves that were laid down in storage tissues during the previous growing season. Annuals growing from seed are rare. The most rapid development of all typically occurs in the herbaceous plants of the forest floor. These plants must take immediate advantage of the spring warmth and sunlight before the new tree foliage casts its heavy shade over the ground, drastically reducing available light energy and slowing the rate of photosynthesis. Many plants of the forest floor have underground energy-storing organs such as bulbs, corms, or fleshy rhizomes that allow them to grow rapidly and strongly in spring to gain maximum advantage from the short, warm, shade-free period. They frequently also produce strong, pointed shoots that are able to emerge above the thick layer of dead leaves that dropped from the trees of the canopy the previous autumn. Thus, the ground of temperate deciduous forests in spring typically is covered by a green carpet of foliage that often includes dramatic displays of colourful flowers before the buds on the trees above have opened. Later, after the leaves of the tree canopy have regrown, the ground cover declines. Typical low-growing plants in North American deciduous forests include species of Cypripedium, Erythronium, Hydrophyllum, Trillium, and Viola. In Europe bluebells (Hyacinthoides non-scripta), daffodils (Narcissus pseudonarcissus), and wood anemones (Anemone nemorosa), among many other herbaceous species, provide similarly spectacular spring flower displays. A vegetation profile of a typical temperate deciduous forest is shown in Figure 2-->.In autumn the delicate leaves of the deciduous trees senesce and start to die. As this happens, they lose their green colour and turn various shades of brown, yellow, and red. Dramatic displays are created, becoming significant tourist attractions in the areas in which the colours are brightest—i.e., eastern North America and western Asia. A weak layer of tissue called the abscission layer develops at the base of each leaf stalk, and at this point the stalk breaks and the leaf is shed. The massive leaf drop that ensues during autumn has earned the season its alternate designation, fall.
As in other forests, the composition of the temperate deciduous forest is commonly determined by the influence of disturbances—natural as well as human—on tree regeneration. For instance, in eastern North America, the tulip tree (Liriodendron tulipifera) produces seeds that can remain dormant in the soil for up to seven years. When they germinate, subsequent establishment of the quick-growing saplings is most successful on bare mineral soil in full light. Stands of this species therefore tend to be of the same age and act as markers of the time at which a catastrophe destroyed the former tree cover, baring the ground. Beech that occur in the same region are shade-tolerant and occupy places not subject to catastrophe, regenerating beneath undamaged tree canopy.
There are still a few temperate forests that have been disturbed little by human activities, and the interrelationships between the vegetation and the large animals are of interest. A rare, relatively intact area of deciduous forest that contains some evergreen conifers is found in Poland. The most common trees include linden (species of Tilia), oak (Quercus robur (English oak)), hornbeam (Carpinus betulus), maple (Acer platanoides), and spruce (Picea abies). Until 1923 large areas were dominated by linden, but most tree regeneration at that time was by spruce, hornbeam, and maple. By 1973 these latter species had become common as canopy trees, and lindens began regenerating freely, as they had not at the earlier time. Such changes result in part from the varying regeneration requirements of the trees. Most successful regeneration takes place where gaps of light reach the forest floor. Competition between saplings of different species is intense and has different outcomes depending in part on soil and light intensities and to a large degree on the effects of mammals. Rooting by wild pigs, although it destroys many small plants, creates suitable conditions for seedling establishment. Saplings are subject to browsing by deer, which also feed on herbaceous plants that compete with tree seedlings. European bison similarly eat foliage and, in addition, can debark trees with their horns. The combined variable influences of these and other animals interact to produce a mosaic forest of different tree ages and composition, with longer-term changes resulting from population fluctuations due, in part, to human impact.
Most temperate forests, where they still survive at all, have been so exploited and disturbed by human influences that their natural condition is difficult to discern. However, some areas outside the old agricultural regions of Asia and Europe have sustained much less human disturbance, making it possible to define natural disturbances responsible for some variations. For example, temperate broad-leaved forests in Chile have been shown to vary in structure and composition according to their history of natural catastrophe in the form of earthquake-induced landslides; similar relationships between ecosystem alteration and natural disturbances have been demonstrated in New Zealand with volcanic eruption and in Tasmania with wildfire. In all these regions, sites undisturbed for many centuries have forests dominated by shady, highly competitive species of Nothofagus, often with few seedlings of any kind beneath the large, old trees. However, in the wake of natural catastrophe, other trees can invade the sites, and only gradually does Nothofagus reestablish itself and slowly resume dominance during subsequent tree generations. Therefore, in areas that have suffered many instances of disturbance, there exists today a variety of forest types. Comparable variations resulting from storm damage and wildfire occurrence have been recognized in North American deciduous forests.
Biological productivity
The total aboveground biomass (dry weight of organic matter in an area) for temperate deciduous forests is typically 150 to 300 metric tons per hectare; values for temperate broad-leaved forests are generally higher, and those for sclerophyllous forests are lower. The subterranean component is more difficult to measure, but it appears to approximate a value of about 25 percent of the aboveground component in deciduous forests and rather more in temperate evergreen forests. Total biomass in temperate deciduous forests is, therefore, about 190 to 380 metric tons per hectare. These values refer to mature, undamaged forests; when disturbances also are considered, the range would include lower values. (For a full discussion of productivity see biosphere: The organism and the environment: Resources of the biosphere (biosphere).)
For temperate forests gross primary productivity (the total biomass fixed by the vegetation in a unit area within a unit time) has been estimated at 16 to 50 metric tons per hectare per year. Net primary productivity, gross primary productivity less that used by plants in respiration, is approximately 10 metric tons per hectare per year; it is greatest in young forests where the trees are rapidly growing toward full size, and it declines in forests of old trees. While the gross primary productivity of temperate forests is considerably lower than that of tropical rainforests, the net primary productivity is not so different, reflecting the lower diversity and complexity of the consumer component of the temperate forest ecosystem.
Temperate forests have been useful to human populations in many diverse ways. Although in most places they have been replaced by simpler agricultural systems, large areas still remain, especially on poorer soils, and are important sources of timber. The total yield as well as the quality of timber is maximized by keeping forests in a condition of greatest net primary productivity—i.e., by harvesting trees before they reach their age of declining growth. Old growth forests have a high biomass and considerable conservational significance but are not efficient in terms of total sustained timber yield.
Additional Reading
Broad descriptions of temperate forests can be found in J.D. Ovington (ed.), Temperate Broad-Leaved Evergreen Forests (1983); and E. Röhrig and B. Ulrich (eds.), Temperate Deciduous Forests (1991). J.R. Packham et al., Functional Ecology of Woodlands and Forests (1992), provides a general account of forest ecology, with particular reference to European deciduous forests. M. Numata (ed.), The Flora and Vegetation of Japan (1974), describes the temperate broadleaf evergreen and deciduous forests of East Asia.
- Horkheimer, Max
- Horlivka
- Hormisdas, Saint
- Hormizd I
- Hormizd II
- Hormizd IV
- hormone
- Hormozgān
- Hormuz
- Hormuzd Rassam
- Hormuz, Strait of
- horn
- Horn, Arvid Bernhard, Greve
- hornbeam
- hornbill
- hornblende
- Hornblower, Jonathan
- hornbook
- Hornbostel, Erich Moritz von
- Horn, Cape
- horn coral
- horn dance
- horned owl
- horned poppy
- horned toad