protozoan
Introduction
 any member of the subkingdom Protozoa, a collection of single-celled eukaryotic (i.e., possessing a well-defined nucleus) organisms. As such, they are among the simplest of all living organisms. Although they comprise a subkingdom, protozoans are not necessarily related to one another. In biological terms, they are not a natural group but simply a collection of organisms. There are more than 65,000 described species, of which over half are fossil.
any member of the subkingdom Protozoa, a collection of single-celled eukaryotic (i.e., possessing a well-defined nucleus) organisms. As such, they are among the simplest of all living organisms. Although they comprise a subkingdom, protozoans are not necessarily related to one another. In biological terms, they are not a natural group but simply a collection of organisms. There are more than 65,000 described species, of which over half are fossil.Protozoa are ubiquitous in most soils and in aquatic habitats from the South to the North poles. Most are invisible to the naked eye. Many are symbionts of other organisms, and about one-third of the living species are parasites. The classification of protozoans requires regular revision because modern electron microscopy and new biochemical and genetic techniques provide an ever-increasing pool of knowledge about the relationships of various protistan species and groups, often showing previous assignments to be incorrect.
General features
 Free-living protozoan groups that inhabit soils and natural waters are extremely diverse, not only in their structure but also in the manner in which they feed, reproduce, and move. Among the mainly free-living groups are the flagellates (Mastigophora). Their name derives from the whiplike structures, or flagella, that are used for movement and feeding. Each flagellate cell bears one or more of these organelles.
Free-living protozoan groups that inhabit soils and natural waters are extremely diverse, not only in their structure but also in the manner in which they feed, reproduce, and move. Among the mainly free-living groups are the flagellates (Mastigophora). Their name derives from the whiplike structures, or flagella, that are used for movement and feeding. Each flagellate cell bears one or more of these organelles.The flagellates exhibit the greatest diversity of nutrition among the protozoa. Many contain pigments also shared by plants, such as chlorophyll, which capture light energy during photosynthesis for the manufacture of carbohydrates and other complex nutrient substances. Thus they possess a plantlike nutrition and are called autotrophs (self-feeders). Other flagellates are colourless (i.e., contain no photosynthetic pigments); they obtain their nutrients by feeding on algae, bacteria, and other protozoa. Such flagellates have an animal-like nutrition and are called heterotrophs. Some colourless flagellates have photosynthetic ecto- and endosymbionts—for example, Oikomonas syncyanotica, which carries cyanobacterial (blue-green algal) symbionts on its surface, and the dinoflagellate Amphisolenia, which contains endosymbiotic cyanobacteria. Some dinoflagellates, such as Noctiluca and Gyrodinium, may have other flagellates living within them as symbionts. Many autotrophic flagellates must also consume bacteria because photosynthesis alone is not sufficient. These flagellates and those with symbiotic algae exhibit a metabolism known as mixotrophy, in which heterotrophy and autotrophy are combined in a variety of ways and to different degrees. Thus, flagellates exhibit the complete nutritional spectrum, from totally plantlike nutrition to completely animal-like nutrition, with varying degrees of both.
In fact, nutrition is not taxonomically significant because many of the phytoflagellates (i.e., the plantlike groups, or Phytomastigophorea) do not contain photosynthetic pigments but feed in a heterotrophic manner. The dinoflagellates are a good example: about one-half do not contain plant pigments, but they are classified as dinoflagellates because in every other respect they are like their coloured relatives. Moreover, even among the coloured dinoflagellates, many are mixotrophs.
While the majority of flagellates are free-living, some have evolved a parasitic (parasitism) way of life. These include the hemoflagellates (class Zoomastigophorea, order Kinetoplastida), so called because at some stage in their life cycle they live in the blood of a vertebrate host. Several hemoflagellates cause disease, such as sleeping sickness and Chagas' disease.

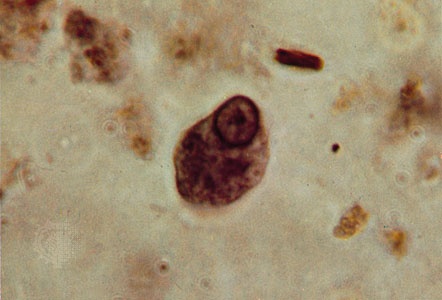 The amoebas (amoeba) (phylum Sarcomastigophora, subphylum Sarcodina) are a diverse group of free-living protozoa that probably evolved from a number of different primitive protozoan ancestors. While they are often regarded as the simplest of protozoans, because many resemble a blob of protoplasm with no apparent organized shape, some members of the Sarcodina (sarcodine) are actually extremely complex. The most sophisticated are the shell-bearing foraminiferans (foraminiferan) (superclass Rhizopoda, class Granuloreticulosea). There are two categories of amoebas: the naked amoebas, which lack skeletal structures, and those that possess a skeleton or shell of some type. Members of the Sarcodina move and feed by means of protoplasmic extrusions called pseudopodia (pseudopodium) (false feet). Pseudopodia vary in both structure and number among the different species. Some possess only one leading pseudopod, while others have a complex multibranched network of pseudopodia. Like the flagellates, the sarcodines also include some parasitic species among their number. A well-known example is Entamoeba histolytica, which causes amoebic dysentery in humans.
The amoebas (amoeba) (phylum Sarcomastigophora, subphylum Sarcodina) are a diverse group of free-living protozoa that probably evolved from a number of different primitive protozoan ancestors. While they are often regarded as the simplest of protozoans, because many resemble a blob of protoplasm with no apparent organized shape, some members of the Sarcodina (sarcodine) are actually extremely complex. The most sophisticated are the shell-bearing foraminiferans (foraminiferan) (superclass Rhizopoda, class Granuloreticulosea). There are two categories of amoebas: the naked amoebas, which lack skeletal structures, and those that possess a skeleton or shell of some type. Members of the Sarcodina move and feed by means of protoplasmic extrusions called pseudopodia (pseudopodium) (false feet). Pseudopodia vary in both structure and number among the different species. Some possess only one leading pseudopod, while others have a complex multibranched network of pseudopodia. Like the flagellates, the sarcodines also include some parasitic species among their number. A well-known example is Entamoeba histolytica, which causes amoebic dysentery in humans.The most evolved and complex protozoans are the ciliates (ciliate) (phylum Ciliophora). The cell surface is covered with hundreds of hairlike structures, or cilia, arranged in ordered rows called kineties. The cilia beat in synchronized waves and thereby propel the organism through the water. Most ciliates possess a cell mouth (cytostome) through which food enters the cell. (Some flagellates also have cytostomes.) In some ciliates, the cilia around the cytostome have become specially modified into sheets called membranelles, which create a feeding current and act as a sieve to trap food particles. Other important ciliate characteristics include the possession of two types of nuclei (a large nucleus, or macronucleus, and one or more small nuclei, or micronuclei, occurring in each cell); sexual reproduction by conjugation; and asexual reproduction by binary fission in an equatorial, or transverse, plane.
A number of the protozoan phyla are exclusively parasitic, either in higher animals or, as in the phylum Labyrinthomorpha, on algae (although some members of this phylum feed saprotrophically on the surface of marine grasses and algae by secreting extracellular enzymes). The entirely parasitic phylum Apicomplexa is particularly important to humans because among its members are those species responsible for causing such diseases as malaria and toxoplasmosis.
The major parasites causing pathological conditions in humans and other vertebrates are found in the apicomplexans and the two mainly free-living groups, the sarcomastigophorans and the ciliophorans. This fact, coupled with the importance of free-living protozoa in ecological processes, means that more is known about these three groups, and this article, therefore, concentrates on the functioning and biology of these protozoans.
Natural history
Size range and diversity of structure
Protozoa range in diameter from a few thousandths of a millimetre to several millimetres. Because the subkingdom contains many unrelated or loosely related groups, there is enormous diversity in structure and form. Even within a single phylum, the variation in form can be considerable.
 The flagellates (flagellum) range from a simple oval cell with one or more flagella to the structural sophistication of the collared flagellates (order Choanoflagellida). The collared flagellates lack photosynthetic pigments and are therefore colourless. They have a single flagellum surrounded by a delicate circular collar of fine pseudopodia on which they trap food particles. In some marine species, the whole cell is enclosed in an elaborate, open latticelike basket formed from strands of silica. The dinoflagellates (dinoflagellate), half of which contain plant pigments and rely to a greater or lesser degree on photosynthesis, may be surrounded by a cell wall armour with a complicated pattern. In some species (e.g., Ceratium), long spines arise from the cell surface and aid in flotation. Dinoflagellates possess two flagella; one beats in a transverse plane around the equator of the cell while the other beats in a longitudinal plane. Many of the flagellates live in colonies. In Volvox, for example, hundreds of individual organisms are embedded in a gelatinous sphere.
The flagellates (flagellum) range from a simple oval cell with one or more flagella to the structural sophistication of the collared flagellates (order Choanoflagellida). The collared flagellates lack photosynthetic pigments and are therefore colourless. They have a single flagellum surrounded by a delicate circular collar of fine pseudopodia on which they trap food particles. In some marine species, the whole cell is enclosed in an elaborate, open latticelike basket formed from strands of silica. The dinoflagellates (dinoflagellate), half of which contain plant pigments and rely to a greater or lesser degree on photosynthesis, may be surrounded by a cell wall armour with a complicated pattern. In some species (e.g., Ceratium), long spines arise from the cell surface and aid in flotation. Dinoflagellates possess two flagella; one beats in a transverse plane around the equator of the cell while the other beats in a longitudinal plane. Many of the flagellates live in colonies. In Volvox, for example, hundreds of individual organisms are embedded in a gelatinous sphere.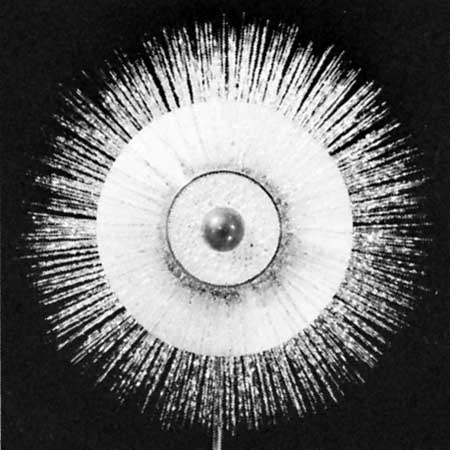 The sarcodines also are extremely diverse. They have four types of pseudopodia—lobopodia, filopodia, axopodia, and reticulopodia. The simplest (lobopodia) are blunt extensions of the protoplasm, and the most complex (reticulopodia) form a complicated branching network. The simplest of the sarcodines, the naked amoebas (Gymnamoebia), have no defined shape and extend one or many pseudopodia. At the opposite extreme are the complex foraminifera, which live inside multichambered calcareous shells up to several millimetres in diameter. The pseudopodia (reticulopodia) of foraminiferans extend from the aperture of the largest chamber of the shell and form a complicated, sticky branching network. Other sarcodines, known commonly as radiolarians (radiolarian) (class Polycystinea), form shells from silica; in some, the shell has so many holes that the structure resembles a sponge. Some of the most exquisite sarcodines are the sun protozoans, or heliozoans (heliozoan). Their radiating pseudopodia (axopodia), extend like spokes from the central body; microtubules support an outer layer of cytoplasm.
The sarcodines also are extremely diverse. They have four types of pseudopodia—lobopodia, filopodia, axopodia, and reticulopodia. The simplest (lobopodia) are blunt extensions of the protoplasm, and the most complex (reticulopodia) form a complicated branching network. The simplest of the sarcodines, the naked amoebas (Gymnamoebia), have no defined shape and extend one or many pseudopodia. At the opposite extreme are the complex foraminifera, which live inside multichambered calcareous shells up to several millimetres in diameter. The pseudopodia (reticulopodia) of foraminiferans extend from the aperture of the largest chamber of the shell and form a complicated, sticky branching network. Other sarcodines, known commonly as radiolarians (radiolarian) (class Polycystinea), form shells from silica; in some, the shell has so many holes that the structure resembles a sponge. Some of the most exquisite sarcodines are the sun protozoans, or heliozoans (heliozoan). Their radiating pseudopodia (axopodia), extend like spokes from the central body; microtubules support an outer layer of cytoplasm.The ciliates are the most structurally homogeneous group, although even they have evolved considerable variation on the cilia-covered cell. In some species (e.g., the hypotrich Euplotes) the cilia are combined to form thick conical structures, called cirri, which the ciliate uses to crawl along surfaces, rather like little legs. In others the cilia virtually disappear from the main body of the cell, but the circle of cilia around the mouth becomes well developed (as in the oligotrich Strombidium and the tintinnid ciliates). The peritrich ciliates have developed stalks and attach to plants and animals as a means of dispersal. Many peritrichs (e.g., Epistylis) form branching colonies.
A group of ciliates, the suctorians (suctorian), have completely lost their cilia in the adult phase. They have instead developed a stalk and many tentacles, which they use to capture passing prey, usually other ciliates. Because they cannot swim, they produce motile ciliated offspring, which settle elsewhere and then transform into the feeding stage, thus avoiding overcrowding.
Although the parasitic protozoa tend to be less structurally complex than free-living forms, considerable variation may occur during the course of their life cycles. Plasmodium, the malarial parasite that lives inside the liver and red blood cells of humans and the gut of its insect vector (the Anopheles mosquito), undergoes various changes in form through its asexual and sexual phases of development. Among the parasitic flagellates, the trypanosomes and their relatives (hemoflagellates), morphological variation occurs during the various stages of the life cycle in both the mammalian and insect host. Among species of leishmania, which cause visceral leishmaniasis (kala-azar), cutaneous leishmaniasis (Oriental sore), and mucocutaneous leishmaniasis (espundia (Kawasaki syndrome)), two distinctly different forms occur. In humans, rounded, nonflagellated forms called amastigotes feed and divide inside macrophage cells in different regions of the body, while in the gut of the insect vector there occurs a flagellated form called a promastigote. Members of the genus Trypanosoma, which cause sleeping sickness and other diseases, have flagellated forms with different morphologies. At some stage in the life cycle, all assume the trypomastigote form—i.e., slender with part of the flagellum running over the body and attached to it by a finlike extension to form an undulating membrane. They may also occur as amastigote (stumpy flagella) or promastigote forms.
Distribution and abundance
Protozoa have colonized a wide array of aquatic and terrestrial habitats from the Arctic and Antarctic to equatorial zones. In soils (soil organism) and bogs, ciliates, flagellates, and amoebas form part of a complex microbial community. They live in the moisture films surrounding soil particles, so that they are actually aquatic organisms, even though living in a terrestrial environment. Between 10,000 and 100,000 organisms per gram of soil may inhabit fertile land; the relative proportions of each group vary depending on soil type and latitude. In Antarctic soils, flagellates and testate (shell-dwelling) amoebas predominate, while in temperate woodland soils, ciliates are more numerous.
In the open waters of lakes, estuaries, and the sea, protozoa form an important component of the floating plant and animal community ( plankton). They are often present in densities of tens of thousands per litre of water. During photosynthesis, flagellates carrying plant pigments transform the energy from the Sun into organic matter, which, along with many algal species, forms the base of the aquatic food chain. Most planktonic protozoa, however, feed on bacteria, algae, other protozoa, and small animals. The most common planktonic protozoa are the zooflagellates, ciliates—especially members of the oligotrichs and the tintinnids (tintinnid) (which live inside small tubes, or loricae (lorica))—and the exclusively marine foraminiferans and radiolarians.
Although few data exist for oceanic deeps, foraminiferans have been found at depths of 4,000 metres, and some protozoans have been observed around hydrothermal vents on the ocean floor.
Importance
Protozoa play important roles in the fertility of soils. By grazing on soil bacteria, they regulate bacterial populations and maintain them in a state of physiological youth—i.e., in the active growing phase. This enhances the rates at which bacteria decompose dead organic matter. Protozoa also excrete nitrogen and phosphorus, in the form of ammonium and orthophosphate, as products of their metabolism, and studies have shown that the presence of protozoa in soils enhances plant growth.
Protozoa play important roles in wastewater treatment (water purification) processes, in both activated sludge and slow percolating filter plants. In both processes, after solid wastes are removed from the sewage, the remaining liquid is mixed with the final sludge product, aerated, and oxidized by aerobic microorganisms to consume the organic wastes suspended in the fluid. In the former process, aerobic ciliates consume aerobic bacteria, which have flocculated; in the latter process, substrates are steeped in microorganisms, such as fungi, algae, and bacteria, which provide food for oxidizing protozoa. In the final stages of both processes, solids settle out of the cleaned effluent in the settlement tank. Treatment plants with no ciliates and only small numbers of amoebas and flagellates produce turbid effluents containing high levels of bacteria and suspended solids. Good-quality, clean effluents are produced in the presence of large ciliated protozoan communities because they graze voraciously on dispersed bacteria and because they have the ability to flocculate suspended particulate matter and bacteria.
Protozoa probably play a similar role in polluted natural ecosystems. Indeed, there is evidence that, by feeding on oil-degrading bacteria, they increase bacterial growth in much the same way they enhance rates of decomposition in soils, thereby speeding up the breakdown of oil (oil spill) spillages.
Some radiolarians and foraminiferans harbour symbiotic (symbiosis) algae that provide their protozoan hosts with a portion of the products of photosynthesis. The protozoans reciprocate by providing shelter and carbon and essential plant nutrients. Many ciliates contain endosymbiotic algae, and one species, Mesodinium rubrum, has formed such a successful relationship with its red-pigmented algal symbiont that it has lost the ability to feed and relies entirely on symbiosis for its livelihood. Mesodinium often forms dense, nontoxic red blooms (or red tides) when it reaches high densities in plankton. Among the ciliates with endosymbionts, Mesodinium is the only completely photosynthetic species. Other ciliates achieve photosynthesis in another way. Although they do not have symbiotic algae, they consume plantlike flagellates, sequester the organelles that contain the plant pigments, and use them for photosynthesis. Because the isolated plastids eventually age and die, they must be replaced continuously.
The impact of protozoan grazing on phytoplankton can be considerable. It has been estimated that at least half of the phytoplankton production in marine waters is consumed by protozoa. Like the soil protozoa, these planktonic protozoans excrete nitrogen and phosphorus at high rates. The protozoans are a fundamental component in recycling essential nutrients (nitrogen and phosphorus) to the phytoplankton.
Parasitic protozoa have invaded and successfully established themselves in hosts from practically every animal phylum, although it is about parasitic species of medical and agricultural importance that most is known. The trypanosomes, for example, cause a number of important diseases in humans. African sleeping sickness is produced by two subspecies of Trypanosoma brucei, namely, T. brucei gambiense and T. brucei rhodesiense. The life cycle of T. brucei has two hosts, humans and other mammals and the blood-sucking tsetse fly, which transmits the parasite between humans.
 Trypanosomes live in the blood plasma and the central nervous system of humans and have evolved an ingenious way of fooling the immune system of the host. Upon contact with a parasite, the immune system generates antibodies that recognize the specific chemical and physical nature of the parasite and actively neutralize it. Just as the host's immune system is beginning to win the battle against the parasite and the bulk of the population is being recognized and destroyed by host antibodies, the parasite is able to shed its glycoprotein coat, which is attached to the cell surface, and replace it with a coat containing different amino acid sequences. Thus, the parasite essentially changes its makeup. These alternate forms are known as antigenic variants, and it has been estimated that each species may have as many as 100 to 1,000 such variants. The host must produce a new set of antibodies against each new variant; in the meantime, the parasite has time in which to replenish its numbers. Ultimately, unless the disease is treated, the parasite wins the battle and the host dies. Such antigenic variation makes the development of an effective vaccine against certain parasitic protozoan diseases virtually impossible.
Trypanosomes live in the blood plasma and the central nervous system of humans and have evolved an ingenious way of fooling the immune system of the host. Upon contact with a parasite, the immune system generates antibodies that recognize the specific chemical and physical nature of the parasite and actively neutralize it. Just as the host's immune system is beginning to win the battle against the parasite and the bulk of the population is being recognized and destroyed by host antibodies, the parasite is able to shed its glycoprotein coat, which is attached to the cell surface, and replace it with a coat containing different amino acid sequences. Thus, the parasite essentially changes its makeup. These alternate forms are known as antigenic variants, and it has been estimated that each species may have as many as 100 to 1,000 such variants. The host must produce a new set of antibodies against each new variant; in the meantime, the parasite has time in which to replenish its numbers. Ultimately, unless the disease is treated, the parasite wins the battle and the host dies. Such antigenic variation makes the development of an effective vaccine against certain parasitic protozoan diseases virtually impossible.A close relative of T. brucei, Trypanosoma cruzi, causes Chagas' disease, or American trypanosomiasis. The vector hosts are bugs (Rhodnius) and other arthropods, such as lice and bedbugs. In humans the nonflagellated (amastigote) form of the parasite lives inside macrophage cells, the cells of the central nervous system, and muscle tissue, including the heart, where it grows and divides. Short trypomastigote flagellated forms periodically appear in the blood, where they are readily taken up by the bloodsucking vector hosts. These flagellated forms do not divide in the blood, reproduction occurring only in the amastigote intracellular forms.
Relatives of the trypanosomes, species of the genus Leishmania, cause a variety of diseases worldwide known as leishmaniasis. Like T. cruzi, these are intracellular parasites of the macrophage cells. The intermediate, or vector, hosts are a variety of sand fly species (Phlebotominae). In cutaneous leishmaniasis (Oriental sore) the infected macrophages remain localized at the site of the infection, causing an unsightly lesion, but in visceral leishmaniasis the infected macrophages are carried by the blood to the visceral organs. This latter disease is characterised by enlargement of the spleen and liver, leading to the distended abdomen that is typical of kala-azar. In mucocutaneous leishmaniasis the initial skin infection spreads to the mucous membranes of the face (the nose, mouth, and throat), producing a lesion that can cause the destruction of part of the face.
malaria, which is caused by the protozoan Plasmodium, remains a serious disease despite both measures that can be taken to control and eradicate the mosquito vector host and the availability of an array of antimalarial drugs. The life cycle is fundamentally identical among the four species of Plasmodium, but the pathology of the disease varies in the frequency and severity of attacks and in the occurrence of relapses. Problems in controlling the disease include the development of resistance to insecticides by the mosquito and the evolution of drug resistance by the parasite. Prophylactic drugs taken before and during a visit to areas where malaria is endemic may prevent the disease from forming in persons who have no natural resistance. Since antigenic variation does not appear to occur in Plasmodium, modern genetic engineering techniques offer promise of producing a vaccine.
The apicomplexan Cryptosporidium (class Coccidea) is a protozoan parasite of humans and other mammals that has become particularly prominent since the 1970s. It has a one-host life cycle and lives inside the cells lining the intestines and sometimes the lungs. Cryptosporidium carries out all the asexual reproductive stages typical of an apicomplexan (see below) inside a single host and is passed from host to host in a resistant cyst stage called an oocyst. The disease caused by the parasite is typified by severe diarrhea and vomiting. Although there is no drug treatment, most healthy people recover quickly. In persons who have impaired immune systems, such as AIDS patients, however, Cryptosporidium can cause serious infections.
Form and function
The protozoan cell
The protozoan cell carries out all of the processes—including feeding, growth, reproduction, excretion, and movement—necessary to sustain and propagate life, although it does so at a somewhat simpler level than do multicellular organisms. The cell is enclosed in a unit membrane called the plasma membrane. Like all membranous structures in the eukaryotic cell, the plasma membrane is composed of protein and lipid molecules. The membrane is a barrier between the cytoplasm and the outside liquid environment. Some substances, such as oxygen, readily pass through the membrane by diffusion (passive transport), while others must be transported across at the expense of energy (active transport). Cilia and flagella arising from the cell are also sheathed in the cell membrane.
The cell also has internal membranes, which are not as thick as the plasma membrane. Among these are the endoplasmic reticulum, whose membranes separate out compartments of the cell, thereby allowing different conditions to be maintained in various parts—e.g., separation of different substances. Enzymes are arranged on the surface of the endoplasmic reticulum; one such enzyme system catalyzes the activity of the ribosomes during protein synthesis. The Golgi apparatus is a cluster of flattened vesicles, or cisternae, associated with the endoplasmic reticulum. The vesicles are concerned with membrane maturation and the formation and storage of the products of cell synthesis, as, for example, in the formation of scales on the surface coat of some flagellates. The scales are formed within the Golgi and are transported by the vesicles to the plasma membrane, where they are incorporated onto the surface of the cell. The Golgi apparatus is well developed in flagellates, poorly seen in most ciliates, and absent from some amoebas.
All protozoa possess at least one nucleus, and many species are multinucleate. The genetic material DNA (deoxyribonucleic acid) is contained within the chromosomes of the nucleus. Each nucleus is bounded by two unit membranes through which pores provide a channel permitting the passage of molecules between the cytoplasm and the nucleoplasm. Most ciliates have two types of nuclei, micronuclei and macronuclei. Almost all protozoan cells contain at least one micronucleus, and many contain more than one. The macronucleus can be quite variable in shape, resembling in some species a string of beads or a horseshoe. It directs the normal functioning of the cell and usually disintegrates during sexual reproduction, to be reformed from the products of micronuclear division after the sexual phase is completed.
Almost all protozoa contain double-membrane mitochondria (mitochondrion); the inner membrane forms fingerlike extensions (or cristae) into the mitochondrial interior, and the outer membrane forms the boundary of the organelle. Mitochondria are the sites of cellular respiration. Species that do not require oxygen (anaerobes), such as those that live in the intestinal tract of their hosts or those that occupy special anaerobic ecological niches, lack mitochondria. They have instead respiratory organelles called microbodies. These oblong or spherical membrane-bound organelles, about one to two micrometres in length, are believed to be the site of respiratory processes. They contain enzymes that oxidize pyruvate to acetate and carbon dioxide, resulting in the release of hydrogen sulphide under anaerobic conditions.
Photosynthetic pigments, when present, are housed in organelles called plastids. These also are bounded by at least two unit membranes. All plastids contain ribosomes and DNA of a type structurally similar to those of bacteria and other members of the kingdom Monera (prokaryotes). Ribosomes and DNA synthesize some of the protein and plastid RNAs of these protozoans independently from the synthesis carried out under the direction of the DNA of the macronucleus. Plastids are believed to have evolved from endosymbionts, and their structure and independent ability to produce proteins and RNA support this hypothesis.
Organisms that live in a liquid environment with a lower concentration of ions than is found in the interior of their cells—that is, a lower osmotic concentration—gradually gain water. If this remains unchecked, the cell swells and bursts. In protozoa the maintenance of the osmotic balance of the cell is achieved by the contractile vacuole. These membrane-bound organelles are situated close to the plasma membrane. They swell periodically and then suddenly contract and disappear, forcing their contents from the cell in repeated cycles. In the amoebas and the flagellates the contractile vacuole is formed when smaller vesicles combine with the main vacuole. In the ciliates the contractile vacuole is fed by a complex system of feeder canals, which are, in turn, fed by a complex of vesicles and fine tubules within the cytoplasm.
Heterotrophic protists have transitory food or digestive vacuoles. The number of these membrane-bound cell organelles depends on the feeding habits of the organism. Some species may have many, while others may contain only one or two at any one time. In ciliates the food vacuoles form at the base of the cytopharynx, while in species without a cell mouth the vacuoles form near the cell membrane at the site where food is ingested.
Within the cell, structural proteins of various types form the cytoskeleton (cell skeleton) and the locomotory appendages. They include microfilaments formed of a contractile protein also found in the muscles of animals (actin) and cylindrical microtubules (microtubule) formed from filaments of the protein tubulin. Microtubules are particularly important in the structural formation and functioning of cilia and flagella. Axopodia of certain flagellate species are supported by microtubules.
The protozoa exhibit diverse modes of locomotion across the various groups, but the modes of locomotion can be broadly divided into flagellar, ciliary, and amoeboid movement. Flagellar propulsion is employed by the flagellates and during some stages in the life cycles of certain sarcodines. The flagellum is a whiplike structure found not only in protozoans but in higher organisms as well (such as in sperm, the male reproductive cells of higher animals). The structure of all flagella is basically the same, consisting of a cylinder (axoneme) made up of a pair of central microtubules surrounded and joined by cross-bridges to a circle of nine pairs of microtubules. This “nine-plus-two” arrangement of the microtubules in the axoneme is surrounded by cytoplasm and ensheathed in cell membrane. The flagellum arises from the basal body, or kinetosome, within the cell.
The undulating motion of the flagellum is normally generated at its base. The waves move along the flagellum to produce a force on the water acting along the long axis of the organelle in the direction of the wave. The speed of movement is determined by the length of the flagellum and by the size of, and distance between, the waves it generates. Some species have hairs (mastigonemes) arising at right angles to the flagellum along its length, while other species have slender hairs called flimmer filaments. Either structure has the effect of altering the movement of water produced by undulations of the flagellum by reversing its flow toward the flagellar base.
Swimming speeds achieved by flagellates are relatively low. Ciliates have an increased number of beating flagella on the cell surface, thereby enabling greater power to be developed against viscous forces, for greater speeds. The structure of a cilium is identical to that of a flagellum, but the former is considerably shorter. Cilia are a type of flagellum arranged in closely aligned longitudinal rows called kineties. A complex system of fibres and microtubules arising from the basal bodies, or kinetosomes, of each cilium connects it to its neighbouring cilia in the kinety and to adjacent ciliary rows. In some species the body cilia may be reduced to specialized cirri, where the kinetosomes are not arranged in the usual rows but instead have a hexagonal pattern interlinked at several levels by fibres and microtubules.
The effective stroke of the cilium is usually planar, but in the recovery stroke the cilium sweeps out to the side, creating an overall beat with a three-dimensional pattern. The cilium performs work against the viscous force of the water during both the effective and the recovery strokes. To be effective, each cilium must beat in a coordinated manner with its neighbouring cilia. A synchronized beat is passed along a ciliary row by means of a hydrodynamic linkage between the cilia. During a beat, each cilium displaces a layer of surrounding water. Displaced water layers overlap between cilia and, as a consequence, interference occurs between the movements of adjacent cilia, creating a hydrodynamic linkage.
Amoeboid movement is characteristic of the sarcodines and some of the apicomplexans. It is achieved by pseudopodia and involves the flow of cytoplasm as extensions of the organism. The process is visible under the light microscope as a movement of granules within the organism. The basic locomotory organelle is the pseudopodium; the way in which movement is effected varies.
A variety of pseudopodial types are found among the naked and testate amoebas (Gymnamoebia and Testacealobosia (testacean), respectively). In some species, a single pseudopodium is extended at any one time; in others, numerous tubular pseudopodia are extended simultaneously. Some amoebas appear saclike throughout locomotion, and no pseudopodia are obvious. The numerous long, stiff protoplasmic extensions (axopodia) of heliozoans shorten and lengthen—the forward axopodia lengthen and become attached, while the posterior axopodia detach and retract—and the amoeba rolls slowly along. The foraminiferans move by extending slender pseudopodia (filapodia), which may be several millimetres long in some species. The extending filopodia branch and fuse with each other so that there is a complex, continuously changing network of pseudopodia pulling the organism along.
Various theories have been proposed to explain how pseudopodia effect movement. A widely accepted model suggests that the ectoplasm at the front of the pseudopod contracts; isometric tension is maintained on the endoplasm, while isotonic contraction in the rear ectoplasm increases pressure on the tail endoplasm to push it forward. In addition, the contractile protein actin and the force-generating enzyme myosin—which can release the energy carried by ATP (adenosine triphosphate) and is found in the muscle contraction system of higher animals—have been isolated from the cytoplasm of amoebas. During movement, the endoplasm moves toward the cell surface at the pseudopodial tip and then gels, while actin filaments polymerize to form a longitudinal network of endoplasm that interacts with myosin- and actin-binding proteins—some of which are attached to the plasma membrane—to create a contractile cytoskeleton. This contraction increases internal hydrostatic pressure, resulting in a flow of cytoplasm toward any area where the endoplasm can extend.
respiration
Aerobic microorganisms are so small that they are able to obtain the oxygen they require for metabolism from the surrounding liquid medium by simple diffusion. The special pigments or structures required for the acquisition and transport of oxygen found in higher organisms are not required in the protozoa. The respiratory pigment hemoglobin has been found in some ciliates (e.g., Tetrahymena), but it does not function as an oxygen-carrying pigment as in humans.
Most species of free-living protozoa appear to be obligate aerobes; that is, they cannot survive without oxygen. As in the cells of higher organisms, their respiration is based on the oxidation of the six-carbon glucose molecule to single-carbon carbon dioxide molecules and water via the Embden-Meyerhof pathway, tricarboxylic acid cycle (Krebs cycle), and cytochrome systems, the last two metabolic processes taking place in the mitochondria. Within a single species, the rate of oxygen consumption varies in relation to such factors as temperature, the stage in the life cycle, and the cell's nutritional status (i.e., whether or not it is well fed).
Obligate anaerobes, in which metabolism must take place in the absence of oxygen, are rarely found among eukaryotic organisms. Some parasitic anaerobic species, however, live in the gastrointestinal tract of humans and other vertebrates or, in one ecological group of ciliates (e.g., Metopus, Plagiopyla, and Caenomorpha), are associated with sulfide-containing sediments. The latter have been found to lack cytochrome activity, and both anaerobes contain microbodies rather than typical protozoan mitochondria. Along with the microbodies, the sulfur protozoa also harbour endosymbiotic and ectosymbiotic bacteria, which may take the metabolic end products released by the ciliates and reutilize them for growth and energy-yielding processes. These ciliates are believed to have reverted from an aerobic metabolism to an anaerobic metabolism in order to exploit a specialized ecological niche rich in bacteria as a food source.
The type of microbodies of the anaerobic intestine-dwelling species, which are called hydrogenosomes, function as respiratory organelles. They possess enzymes that oxidize pyruvate to acetate and carbon dioxide. Under anaerobic conditions this also results in the release of hydrogen; when oxygen is present, the hydrogen combines with the oxygen to form water.
Certain parasitic protozoa that live in the blood, such as Trypanosoma brucei, have evolved a system of aerobic respiration that does not involve the mitochondria. The initial stages of glycolysis in the Embden-Meyerhof pathway are the same, but glucose, rather than being broken down completely to carbon dioxide and water, is broken down only to the three-carbon molecule pyruvic acid, which is then excreted. The subsequent stages (the tricarboxylic acid cycle and the cytochrome system), which usually take place in the mitochondria, do not occur; instead, the terminal respiration is mediated by an L-α-glycerophosphate oxidase–L-α-glycerophosphate dehydrogenase system located in small membrane-bound vesicles throughout the cytoplasm.
Metabolism and nutrition

 The protozoa display a range of nutritional types, from the entirely plantlike photosynthetic (or autotrophic) nutrition to the totally animal-like (or heterotrophic) nutrition, in which bacteria, algae, other protozoa, and small animals like the crustacean copepods constitute the food source.
The protozoa display a range of nutritional types, from the entirely plantlike photosynthetic (or autotrophic) nutrition to the totally animal-like (or heterotrophic) nutrition, in which bacteria, algae, other protozoa, and small animals like the crustacean copepods constitute the food source.The coloured flagellates, or phytoflagellates (phytoflagellate) (Phytomastigophorea), contain a variety of pigments that trap the Sun's radiant energy and use it to synthesize complex carbohydrates from carbon dioxide and water in the process of photosynthesis. Many coloured flagellates combine autotrophy with heterotrophy and are, strictly speaking, mixotrophs. Some members of the Euglenida, Cryptomonadida, and Volvocida, for example, are commonly called the acetate flagellates because their preferred food sources are acetates, simple fatty acids, and alcohols. In the presence of the correct nutrients, these flagellates are able to switch from carbohydrate-producing photosynthesis when light is available to heterotrophy on acetate and other substrates when it is not. Many planktonic marine and freshwater phytoflagellates also feed voraciously on bacteria. Indeed, in some lakes they may be the main consumers of bacteria suspended in the plankton. It is believed that this ingestion of bacteria (phagotrophy) provides the flagellates not only with an additional source of carbon to supplement what is gained by photosynthesis but also with phosphorus and nitrogen, which are often scarce in planktonic waters, and possibly with vitamins, all of which are essential to photosynthesis. Bacteria are more efficient at taking up these nutrients because they have a higher surface-to-volume ratio than do the flagellates. Thus, one way for the flagellates to acquire essential nutrients is to consume the bacteria.
Heterotrophic protozoans (Zoomastigophorea, or zooflagellates (zooflagellate)) may take food into the cell at a specific point, such as the cytostome, at a particular region of the cell surface, or at any random point of entry. In the collared flagellates (choanoflagellate), for example, the collar and flagellum operate in feeding. The collar, composed of fine pseudopodia, surrounds the flagellum. The beating flagellum creates a water current, causing water to move through the collar (filter feeding). Particles of food in the current are trapped on the collar and are ingested by pseudopodia at its base. The ingested food is then enclosed in a membrane-bound digestive or food vacuole.
Many ciliates are also filter feeders, creating water currents with special ciliary structures associated with the cytostome. The synchronized beating of these ciliary structures pushes a stream of water against a membranelle composed of cilia; the membranelle acts as a collecting sieve, where the food particles become trapped in the free spaces between the cilia. Using this mode of feeding, ciliates can shift considerable volumes of water in relation to their size. Tetrahymena, for example, can filter 3,000 to 30,000 times its own volume in one hour.
Other ciliates lack complex oral cilia and gather their food by other means. Nassula has a complex cytostome and cytopharynx supported by a basketlike cytopharyngeal structure composed of microtubules. This species ingests filamentous algae by grasping the filament, bending it like a hairpin, and drawing it into the cytopharynx, where it is broken up into fragments and enclosed in digestive vacuoles. Predatory ciliates such as Didinium nasutum, Lacyrmaria olor, and Dileptus anser apprehend their prey with special structures called extrusomes. Among the various types of extrusomes are the toxicysts, which are found in the oral region and release toxins that paralyze the prey. The suctorians are ciliate predators that usually possess tentacles of two functional types, feeding tentacles and piercing tentacles; the latter trap and immobilize the prey, usually other ciliates that make chance contact with the outstretched tentacles of the suctorian. The cell contents of the prey are transported up through the feeding tentacles into the suctorian, where digestive vacuoles are formed. The transporting mechanism is mediated by a complex array of microtubules within the tentacle. A single suctorian can often feed on several prey at the same time, and frequently the prey are larger than the predator.
The sarcodines, all of which lack a cell mouth, or cytostome, also exhibit a diverse array of feeding mechanisms and diet. Some feed on filaments of cyanobacteria (blue-green algae)—which are composed of long chains of individual cells—by taking in the entire filament at any point on the cell surface and rolling it up into a coil inside a digestive vacuole. Others, such as the testate amoeba Pontigulasia, pierce single cells in algal filaments and remove the contents. The radiolarians and foraminiferans trap a wide range of prey, including protozoans, algae, and small crustaceans, in their complex pseudopodial networks and then convey the food items to the main body of the cell for ingestion.
 Parasitic protozoa feed in a variety of ways. Many live in the nutrient-rich medium of the body fluids—e.g., the blood or cells of their host. There they take in energy-rich fluids by pinocytosis, in which small amounts of the medium are pinched off into digestive vacuoles either at a specific site, such as the cytostome in ciliates or the flagellar pocket in trypanosomes, or along the surface of the cell in amoebas. Other parasitic protozoa engulf portions of the host tissue ( phagocytosis) in much the same way that free-living amoebas feed. Plasmodium, for example, engulfs portions of the red blood cells or liver cells in which they live. The hemoglobin in the cytoplasm of the red blood cell is only partially digested by the parasite; the protein portion of the hemoglobin molecule is degraded to its constituent amino acids, but the iron-containing portion is converted into insoluble iron-containing hemozoin, which remains within the parasite's endosomes until discarded at the next division. This process removes free hematin from the parasite cytoplasm, where it would otherwise prevent further metabolism within the parasite because it inhibits the actions of succinic dehydrogenase, an enzyme in the Krebs cycle.
Parasitic protozoa feed in a variety of ways. Many live in the nutrient-rich medium of the body fluids—e.g., the blood or cells of their host. There they take in energy-rich fluids by pinocytosis, in which small amounts of the medium are pinched off into digestive vacuoles either at a specific site, such as the cytostome in ciliates or the flagellar pocket in trypanosomes, or along the surface of the cell in amoebas. Other parasitic protozoa engulf portions of the host tissue ( phagocytosis) in much the same way that free-living amoebas feed. Plasmodium, for example, engulfs portions of the red blood cells or liver cells in which they live. The hemoglobin in the cytoplasm of the red blood cell is only partially digested by the parasite; the protein portion of the hemoglobin molecule is degraded to its constituent amino acids, but the iron-containing portion is converted into insoluble iron-containing hemozoin, which remains within the parasite's endosomes until discarded at the next division. This process removes free hematin from the parasite cytoplasm, where it would otherwise prevent further metabolism within the parasite because it inhibits the actions of succinic dehydrogenase, an enzyme in the Krebs cycle.Whatever the mode of heterotrophic nutrition or diet, the food material is enclosed in food vacuoles, which are bounded by cell membrane. Digestive enzymes are poured into the newly formed vacuole from the surrounding cytoplasm. In the ciliate Paramecium, where the process has been researched in detail, it is known that the digestive vacuoles initially decrease in size and the enclosed particles aggregate. As digestion proceeds, the vacuole increases in size and the contents become progressively acidic, before gradually becoming alkaline near the end of the process. The products of digestion are then absorbed into the surrounding cytoplasm, and the waste material is ejected from the cell anus, or cytoproct. The length of the digestive cycle varies and depends on the species and the diet.
Paramecium contains a reservoir of membrane-forming material in discoid vesicles for the purpose of producing food vacuoles. The food vacuoles form at the cytopharynx when the cytopharyngeal membrane and the discoid vesicles fuse. At the cytoproct, where the vacuoles are broken down and the waste material of digestion is ejected, the membrane material is retrieved and returned to the cytopharynx. Thus, the pool of digestive vacuole membrane is continuously recycled within the cell.
While they seem to lack a sensory system, protozoans are capable of food selection. Many of the filter feeders apparently discriminate solely on the basis of size, dictated by the dimensions of the spaces in the membranelle acting as a sieve. Some filter-feeding ciliates, such as the tintinnids, however, are known to be selective and appear to be able to capture or reject items that arrive at the feeding membranelles in the feeding current. The large ciliate Stentor, for example, takes ciliates in preference to flagellates and algae, and discrimination increases as the animal becomes less hungry. Carnivorous species exercise distinct selectivity. Most suctorians feed exclusively on particular ciliate taxa. They are selective feeders and usually do not capture flagellates, sarcodines, or their own ciliated swarmers. Evidence suggests that a reaction between chemical compounds on the surface of the prey and the tentacle tip of the suctorian is responsible for feeding selectivity. Sarcodines also display feeding selectivity. Amoeba proteus, for example, selects the flagellate Chilomonas paramecium in preference to Monas punctum, even when the number of Monas in the medium is high. In this case, selection may be based on the digestibility of the prey; the latter is digested in 3 1/2 hours, the former in 3 to 18 minutes.
Mixotrophy is a common phenomenon among free-living ciliates and sarcodines. Moreover, the degree of mixotrophy varies from complete reliance on the symbiotic alga or algae to transitory retention of the plastids of phytoflagellate prey with only a partial dependence on photosynthesis to supplement the cell's energy balance. Some phytoflagellates (e.g. Dinobryon and Ochromonas), which are primarily autotrophic, also feed on bacteria and are consequently mixotrophic, but this represents a different kind of mixotrophy from that practiced by the fundamentally heterotrophic ciliates and sarcodines.
Many of the foraminiferans and radiolarians possess symbiotic algae. In some foraminiferans and radiolarians several different symbiotic species of algae may live within the protozoan cytoplasm. During the day, the endosymbionts are distributed in the pseudopodial network, but at night they are withdrawn close to the main body of the cell or into the shell. Many thousands of these algae may exist within a single protozoan, and a significant amount of the products of photosynthesis (e.g., glucose, alanine, maltose) are transferred from the algae to the protozoan. Indeed, in some circumstances, the protozoan can survive on this source of energy if deprived of food, although its growth may be impaired.
In another form of mixotrophy, the sarcodines and ciliates sequester the plastids of their phytoflagellate prey and use them for photosynthesis. The plastids do not replicate inside the protozoan as they do in the symbiotic algae and must be replaced continuously. The large marine ciliate Tontonia appendiculariformis, for example, may contain thousands of plastids that have been derived from a variety of flagellates; moreover, the ciliate appears to be selective in its choice of prey from which to derive plastids.
Reproduction and life cycles
Asexual reproduction is the most common means of replication by protozoans. The ability to undergo a sexual phase is confined to the ciliates, the apicomplexans, and restricted taxa among the flagellates and sarcodines. Moreover, sexual reproduction does not always result in an immediate increase in numbers but may simply be a means of exchanging genetic material between individuals of the same species (conjugation). Free-living protozoans normally only resort to sexual reproduction when environmental conditions become adverse, because this mode of reproduction enhances the fitness of the population and increases the chance of mutation. When food and other conditions are favourable, asexual reproduction is practiced.
Asexual reproduction in free-living species usually involves nuclear division and the division of the cell into two identical daughter cells of equal size by binary fission. In parasitic protozoa and some free-living species, multiple fission, resulting in the production of many offspring that may not resemble the parent cell, is normal. During the cycle of growth and division, the protozoan undergoes a series of identifiable phases: a division phase, a growth phase during which the cell increases substantially in size, a phase of DNA synthesis, and a phase of preparation for division, which extends from the end of DNA synthesis until the initiation of division. The division of the cytoplasm is preceded by the division of the nucleus or nuclei.
The plane of division in protozoan cells varies among the different groups and is of taxonomic significance. The flagellates normally divide in a longitudinal plane. The usual process starts at the front end with the division of the flagella and the associated structures; simultaneously, the nucleus divides. The cytoplasm then splits from front to back into two identical daughter cells. The ciliates normally divide in an equatorial, or transverse, plane, thereby maintaining the correct number of ciliary rows, or kineties. The cell mouth and any specialized cilia around it are replicated in different ways among the various ciliate groups, depending on the complexity of the cytostome. The replication of the cytostome precedes the division of the cytoplasm. Some ciliates (e.g., Colpoda) divide within thin-walled reproductive cysts into two daughter ciliates, each of which then divides so that the cyst contains four progeny, which are released when the cyst wall ruptures.
The sedentary suctorians do not reproduce by binary fission because the production of an identical, nonswimming offspring would rapidly lead to overcrowding. They instead produce single ciliated offspring called swarmers by a process called budding. Budding can occur endogenously, in which the bud forms within the parent and is ejected when mature, or exogenously, in which the swarmer is formed outside the parent. The swarmers swim away from the parent, settle on a substrate, lose their cilia, and develop feeding tentacles and an attaching stalk.
Naked amoebas (rhizopods) have no fixed plane of division but simply round up and divide into two basically equal halves. The testate amoebas (testacean) (also rhizopods), which live in single-chambered shells, or tests (test), exude the daughter from the aperture of the shell. In species that have a shell formed from silica plates, the daughter contains the plates used to produce the shell but remains attached to the mother cell until the shell is fully formed, when the final severing of the cytoplasm between the individuals occurs. Some of the testate amoebas live inside proteinaceous shells. There, too, the new shell is secreted before binary fission is completed.
The foraminiferan and radiolarian sarcodines have evolved multiple fission. Both produce many flagellated swarmers, or zoospores. The common planktonic foraminiferan Globigerinoides sacculifer, for example, can produce 30,000 swarmers at one time. Each swarmer is about 5 micrometres (0.005 millimetre) long. In planktonic species the parent usually loses buoyancy and sinks by shedding spines and withdrawing the complicated pseudopodial network into the shell. The swarmers are produced in deep water and migrate upward as they mature. Each secretes a shell around itself, which is added to as the organism grows.
The foraminiferans are unusual among free-living protozoans in that a sexual phase is a regular part of the life cycle, alternating with an asexual phase. During the life cycle two types of swarmer are produced. One type, zoospores, have half the number of chromosomes of the parent (i.e., they are haploid); they grow until they become mature adults and can produce and release large numbers of gametic swarmers. These gametes are identical (isogamous) but are comparable to the eggs and sperm of higher organisms. The gametic swarmers fuse in pairs, thus restoring the full complement of chromosomes (i.e., they are diploid), and each individual grows, matures, and ultimately produces haploid zoospores.
Sexual reproduction among the flagellates is not widespread and can involve identical gametes (isogamy) or distinct male and female gametes (anisogamy). The female gametes are larger and are stationary, whereas the male gametes are smaller, produced in larger numbers, and motile.
Sexual reproduction among the ciliated protozoans takes the form of conjugation. The process does not result in an increase in numbers, but is a simple exchange of genetic material between two individual cells. Conjugation occurs only between compatible mating strains within a species, and each species may contain many mating strains. Before conjugation occurs, special chemical signals, called gamones, are released by some ciliates. The gamones cause compatible mating strains to undergo processes that facilitate conjugation. In other ciliates, such as Paramecium, gamones are bound to the cell surface and elicit their responses when the ciliates make physical contact.
During conjugation, two ciliates line up side by side. The macronucleus, which plays no part in the process, disintegrates. A series of nuclear divisions of the micronuclei in each ciliate then ensues, including a meiosis, during which a number of haploid micronuclei are produced in both cells. All but one of these haploid micronuclei disintegrate. The remaining haploid micronucleus in each cell then divides through mitosis into two haploid nuclei (gamete nuclei). A bridge of cytoplasm forms between the two ciliates, and one gametic nucleus from each cell passes into the other cell. The two gametic nuclei in each cell unite, thus restoring the diploid number of chromosomes. The micronucleus undergoes two mitotic divisions to produce four micronuclei; two of these will form the new micronuclei of the cell and two are destined to become the macronucleus. Following the process of conjugation, normal binary fission proceeds. The number of macronuclei and micronuclei formed is dependent on the species and remains the same as the original number.
When no suitable mating partner is available, ciliates may undergo a form of conjugation called autogamy, in which all of the nuclear processes described above occur. But, because only one individual is involved, there is no exchange of gametic nuclei; instead, the two gametic nuclei within the cell unite to form the restored micronucleus.
Specialized sedentary suctorian ciliates practice a modified form of conjugation. The conjugating individuals differ in appearance. The macroconjugants resemble the normal feeding individuals, and the microconjugants resemble the swarmers, although smaller. When a microconjugant locates a macroconjugant, it enters and fuses with it. This is quite different from the temporary association between two cells that occurs in most ciliates.
As is common with other parasitic organisms, parasitic protozoans face the problem of how to disperse from one host to another. In order to increase the probability of finding more hosts, most parasitic protozoa reproduce in high numbers. A representative life cycle of a parasitic protozoan can be found in members of the parasitic phylum Apicomplexa. These protozoans have a complex life cycle that involves a series of stages characterized by episodes of asexual multiple division called schizogony. In the parasite Plasmodium, for example, this phase of the life cycle occurs in the liver and red blood cells of humans. The parasite (sporozoite) enters the host's cells and grows while feeding on the cell contents. It then undergoes a multiple asexual division (schizogony) into many individuals (merozoites). The host's cell wall ruptures, permitting each individual to invade a new red blood cell and repeat the process.
In certain merozoites a sexual cycle is eventually initiated inside the red blood cell, and male and female gametes are produced. The male gametes (microgametocytes) are small, while the female gametes (macrogametocytes) are larger. The life cycle continues if the gametocytes are taken up by a feeding female mosquito of the genus Anopheles. Only the gametocytes can infect the mosquito. Inside the mosquito's gut the haploid gametes fuse to form a diploid zygote, which then undergoes sporogony, a process of multiple divisions in which many sporozoites are produced. The sporozoites migrate to the salivary glands of the insect and are injected into a new host when the mosquito next feeds. They are carried by the blood to the liver, where they undergo their first schizogony inside liver cells, thereafter invading the red blood cells for repeated cycles of schizogony.
The parasitic flagellates reproduce entirely by asexual means and do not appear to have a sexual phase in their life cycles. There is, however, evidence of genetic exchange between certain subspecies of Trypanosoma brucei, although the process by which this occurs is not known.
Adaptations
For the most part, parasitic protozoans live in a fairly constant environment. Temperature fluctuates very little, or not at all, inside the host, there is no risk of desiccation, and food is in constant supply. Free-living protists, on the other hand, face short- or long-term changes in temperature, acidity of the water, food supply, moisture, and light. Many protozoa respond to adverse environmental conditions by encysting. They secrete a thick, tough wall around themselves and effectively enter a quiescent state comparable to hibernation. The ability to form a resistant cyst is widespread among diverse protozoan groups and probably developed early in their evolutionary history. Resting cysts also are easily carried by the wind and form an important means of dispersal for species that live in the soil or are common in temporary ponds and pools. In climates with distinct cold seasons, the cyst may be an important phase in the annual life cycle.
The cyst wall is composed of a varying number of layers, the components of which are dependent on the species. During the encystment process, the protozoan cell undergoes a series of changes that considerably reduce the complexity of the organism. Flagellates and ciliates lose their flagella and cilia, the contractile vacuole and food vacuoles disappear, and the distribution of organelles within the cell may be reorganized. In some species, the cell volume reduces considerably. These changes are reversed during the process of excystment.
Certain of the tintinnid ciliates that live in the plankton of seas are programmed to break out of their cysts en masse at times of the year when the food supply is abundant. Helicostomella subulata, for example, excysts in June in temperate waters and becomes numerous from July through October. It encysts again in October, sinking to the sediments, where it remains until the following year. The cyst is a normal part of the annual life cycle, and even laboratory populations of this ciliate encyst at the same time as the natural population. This type of life strategy pattern has been demonstrated in several other ciliates and in sarcodines.
For soil-dwelling protozoa, the cyst is an important refuge when soil moisture disappears or when soil water becomes frozen. In soils that are subject to freezing and periodic short-term thawing, the protozoa rapidly excyst, feed, and reproduce and then encyst again when soil water becomes temporarily unavailable to them.
The cyst plays an important role in the life cycles of several parasitic protozoans that have a free-living dispersal stage, such as Entamoeba histolytica and Cryptosporidium. The cysts are excreted in the host's feces and survive in water or the soil. Humans are usually infected through drinking contaminated water or eating raw fruit and vegetables grown where human feces are used as fertilizer.
Some freshwater protozoans, especially the ciliates Spirostomum, Loxodes, and Plagiopyla, avoid unpleasant conditions, especially lack of oxygen, by abandoning their bottom-dwelling way of life and swimming upward to position themselves at a level where some oxygen is available but where they are not in direct competition with planktonic species. They remain there until oxygen again becomes available on the lake bottom, at which time they migrate downward.
The widespread occurrence of mixotrophy involving algal symbiosis and the retention and sequestration of the plastids of flagellate prey by planktonic ciliates and sarcodines is believed to be an adaptation to waters where food is limited. Ciliates that retain plastids appear to be far more common in waters where food is scarce than in productive waters. There appears to be an inverse relationship between this form of mixotrophy and the productivity of the waters.
Evolution and paleontology
Protists dominated life on Earth 1.5 billion years ago, giving rise eventually to multicellular organisms. While protozoa evolved early and have survived to the present day as unicellular organisms, they have undoubtedly undergone considerable evolutionary change. That many species must have become extinct as others appeared can be deduced from the limited fossil record. Extinct fossil foraminiferan species, for example, number around 34,000, while there are only about 4,000 described living species.
Only a small number of protozoans have left fossil remains. The calcareous shells of the sarcodine foraminiferans and calcium-secreting flagellate coccolithophores, for example, have produced substantial geologic strata in the chalk formed during the Cretaceous Period (144 to 66.4 million years ago) and the well-developed foram-limestones of the Upper Paleozoic Era (570 to 245 million years ago), Early Cretaceous Period (144 to 97.5 million years ago), and Cenozoic Era (66.4 million years ago to the present). Another fossil-forming group includes the radiolarians (radiolarian), which date to late Precambrian times.
The most abundant and important fossil protozoans are the foraminiferans. This entirely marine group is extremely important as stratigraphic markers in oil exploration. Because species have appeared and then become extinct frequently during geologic history and because they have fairly wide geographic distribution, particularly planktonic species, their value is in showing distinct phases in geologic history and, with specific species, in typifying particular beds of rock or strata.
The poor fossil record of protozoans has hampered attempts at unraveling the complexities of their evolution. Modern biochemical and electron microscopy techniques, however, are providing evidence for new affinities between groups and are elucidating possible evolutionary pathways. Comparisons of flagellar structures, mitochondria, and nuclear and plastid characteristics in conjunction with ribosomal RNA (ribonucleic acid) sequences are revealing the relationships of various taxa.
The main protozoan groups seen today—the ciliates, flagellates, sarcodines, and apicomplexans—almost certainly separated early in geologic history from amoeboid and flagellate ancestors and represent independent ancestral branches of the protozoan evolutionary tree. The ancestral eukaryote organism is thought to have been an amoeboid creature that relied on anaerobic metabolism. The evolution of mitochondria (the centres of aerobic respiration in the cell) as organelles from endosymbiotic bacteria and the establishment of oxidative pathways allowed a more efficient cellular energy balance, which led the way to the evolution of an enormously diverse array of eukaryotic organisms. Some of the early amoeboid eukaryotes developed flagella to enhance their food-gathering abilities and to provide a more efficient mode of propulsion. The flagellates gradually evolved different ways of life, and their structures became modified accordingly. As phagotrophs that ingested bacteria for food, they came in some cases to establish symbiotic associations with photosynthetic species, and ultimately the endosymbionts became plastids within the cell. Some of the flagellates came to depend entirely on photosynthesis and to abandon heterotrophy completely, though many still retain both heterotrophic and autotrophic nutrition as mixotrophs. (Some presentday mixotrophs, however, may be only secondarily mixotrophic, having reestablished heterotrophy in conjunction with photosynthesis.)
A considerable number of protozoans became parasitic, a mode of life that evolved independently among the protozoa many times. Ciliates and sarcodines became symbionts in the intestinal tracts of both vertebrates and invertebrates as a result of surviving the digestive enzymes of the predator. (Most present-day parasites among these protists are intestinal parasites.) Once inside the intestine of the host, they multiplied and gradually, through mutation and selection, came to rely on the resistant cyst as a means of survival and dispersal, losing the ability to survive in a free-living feeding form. Some species subsequently started to invade the tissues of their host (e.g., Entamoeba histolytica), resulting in disease.
The process of parasitism probably arose several times among the parasitic flagellates. The trypanosomes, for example, evolved from free-living zooflagellates, which adapted to living in the alimentary canal of primitive invertebrates in Precambrian times (3.8 billion to 570 million years ago). They evolved with their hosts, becoming symbionts in a wide variety of invertebrates, including annelids, nematodes, and mollusks. It was in the insects, however, that they underwent their most extensive evolutionary explosion into two stocks. At this stage they were transmitted from insect to insect by resistant cysts passed in the feces and ingested by subsequent hosts. When insects developed the habit of sucking vertebrate blood as an energy source, which is believed to have occurred about 40 million years ago, the protozoan symbionts that lived in the gut entered the blood of vertebrates, probably as feces left by the insect were rubbed into the wound. The blood provided a rich environment for the flagellates and thus evolved the two-host life cycles seen today in the Leishmania and Trypanosoma groups.
The apicomplexans (apicomplexan), which also inhabit the blood of vertebrates at some stage in their life cycle, probably evolved from a basal primitive stock seen today as the gregarines, which are parasites of invertebrates. They gave rise to a group of parasitic organisms of which the Coccidea, with a one-host life cycle, are primitive survivors. At first these protozoans lived in the guts of their vertebrate host, but they gradually began invading host tissues and eventually became adapted to spending part of their life cycle in the bloodstream. There they were taken up by blood-feeding insects, and an insect vector host became incorporated into the life cycle. Associated modifications in the reproductive pattern, as seen in Plasmodium, which belongs to the Haemosporina, also occurred. This series of events appears to have happened at least twice in the evolution of apicomplexan life cycles.
Classification
General principles
Until the 1970s the general view was that the protozoans were animals, and as such they were placed in the animal kingdom as the phylum Protozoa. Under this system of classification, zoologists placed the coloured flagellates in the phylum Protozoa despite their obvious plant affinities. Botanists, on the other hand, classified the same organisms as algae, which were regarded as plants. Protozoans are now regarded as a phylum or subkingdom of the kingdom Protista (sometimes called the Protoctista), which also includes the algae. The Society of Protozoologists, which periodically reviews the systematics of the group, favours the subkingdom level for the Protozoa.
Protozoan systematics remains a subject of debate and change. Protozoans comprise a large, unwieldy assemblage, and assignments of species to particular taxa change as new biochemical techniques and electron microscopy studies provide more details on the affinities of various species. The subkingdom Protozoa essentially represents a level of organization—that is, single-celled eukaryote organisms—and does not necessarily indicate single major branches of an evolutionary tree comparable with the kingdoms Plantae or Animalia. Thus, while affinities between some of the groups exist, this is not the case with all the phyla within the subkingdom. Moreover, certain groups (e.g., the Sarcodina (sarcodine) or amoebas) are considered by some researchers to have had many different ancestors and lineages (polyphyletic). The Labyrinthomorpha, Myxozoa, and Microspora have no clear affinities either with one another or with other protozoan groups; they are placed in Protozoa until more information is available.
Diagnostic features
The subkingdom Protozoa has been reviewed and divided by a committee of the Society of Protozoologists into six phyla. A major review was undertaken in 1980 by a committee headed by N.D. Levine, and a series of changes have been made subsequently as outlined in a major work on protozoan systematics published by the society in 1985. The six phyla include the Sarcomastigophora (flagellates and amoebas), the Ciliophora (ciliates), and the entirely parasitic Apicomplexa, all of which are discussed in this article. At least some of these groups show affinities in having common amoeboid or flagellate ancestors early in their evolutionary history. The remaining three phyla are totally unrelated to one another and to the phyla discussed here.
In this assemblage of organisms, the only common feature is a single-celled level of organization. Such a situation invariably means that within the whole group there are considerable differences in structure physiology, life mode, and life cycles.
In addition, the classification of protists generally continues to be debated, and a standard outline of the kingdom has not been established. The differences between the classification of protozoa given below and that given in the article protist reflect taxonomic variations that arise from individual interpretations.
Annotated classification.
Phylum Sarcomastigophora
All have one type of nucleus, though some may be multinucleate; Foraminiferida may show nuclear dimorphism similar to that seen in ciliates; sexual reproduction is not widespread; flagella or pseudopodia, and in some species both, occur at one or another stage in the life cycle.
Subphylum Mastigophora (flagellate)
At least 1, and often many, flagella as feeding and locomotory organelles; binary fission resulting in 2 identical daughter cells is the usual mode of asexual reproduction (symmetrogenic binary fission); sexual reproduction occurs in some groups.
Class Phytomastigophorea (phytoflagellate)
Typically possess chloroplasts, with a range of photosynthetic pigments; some members lack chloroplasts but have an evident relationship to the pigmented forms; all free-living, solitary or colonial; typical species include the green species Volvox (which belongs to the order Volvocida) and Euglena (of the order Euglenida) and the colourless species Peranema (Euglenida) and Noctiluca (a dinoflagellate from the order Dinoflagellida).
Class Zoomastigophorea (zooflagellate)
Colourless flagellates; heterotrophic nutrition; binary fission; some groups, such as the collared flagellates, produce flagellated swarmers; both free-living and parasitic species; includes many free-living groups, among which are the collared flagellates (Choanoflagellida) and the amoeboflagellates, having both amoeboid and flagellated forms in the life cycle (Cercomonadida), and various parasitic orders and families; includes Trypanosoma brucei gambiense and Trypanosoma cruzi (both of the order Kinetoplastida).
Subphylum Opalinata
Previously classified with the ciliates because of their outward appearance in being covered with rows of cilia; their kinetosomes differ from those of the ciliates; they have only 1 type of nucleus, whereas the ciliates have 2 nuclear types; they lack a cytostome; most divide by symmetrogenic binary fission; asexual and sexual phases of reproduction alternate in the life cycle; all members commensals in the alimentary tracts of amphibians.
Subphylum Sarcodina (sarcodine)
Not necessarily closely related; pseudopodia of varying types for locomotion and feeding; most are free-living and divide by binary fission; sexual reproduction is not widespread but does occur in some groups, such as the foraminiferans; some groups are naked; others have shells, or tests, formed from calcium, silica, or protein; others have a structured internal cytoskeleton.
Superclass Rhizopoda (rhizopod)
Includes all species that use different types of pseudopodia for movement (filopodia, lobopodia, and reticulopodia), such as naked amoebas (Gymnamoebia), testate forms (Testacealobosia), the foraminiferans (Granuloreticulosea), and some parasitic forms, but not forms that use axopodia.
Superclass Actinopoda
Radial stiffened axopods with a central rod composed of microtubules; many have mineral material incorporated into the cytoskeleton; mostly marine, many planktonic; includes Polycystinea (radiolarians, e.g., Hexacontium), Heliozoea (the heliozoans, or sun protozoans, e.g., Actinosphaerium eichhorni), Acantharea (e.g., Acanthochiasma rubescens), and Phaeodarea (e.g., Planktonella atlantica).
Phylum Labyrinthomorpha
An ectoplasmic network of spindle-shaped or spherical nonamoeboid cells; in some, amoeboid cells move within the network by gliding, using a unique cell-surface organelle associated with the ectoplasmic network; produce zoospores; saprozoic or parasitic on algae; mostly in marine and brackish waters.Phylum Apicomplexa
Previously called Sporozoa; parasitic in both vertebrates and invertebrates; vesicular nucleus; lack flagella or cilia, except in the flagellated microgamete stage; usually a sexual phase in the life cycle with male and female gametes; schizogony and sporogony are features of the life cycle, and cysts are often present at some stage in species with one-host cycles; typical species include Plasmodium falciparum, Toxoplasma gondii, Cryptosporidium, and Eimeria species.Phylum Microspora
Not analogous with any other protozoan group; include intracellular parasites of a wide variety of animals; produce spores at some stage in their life cycle, and all active stages develop in the host's tissues, specifically in the cytoplasm; invasive stage is the sporoplasm, a tiny uninucleate or binucleate structure only 1 micrometre in diameter; both schizogony and sporogony occur in the life cycle; a typical species is Pleistophora typicalis.Phylum Myxozoa
Parasites of the tissues and organ cavities of cold-blooded vertebrates, especially fish, and annelid worms; spore producers; typical species is Myxidium giardi.Phylum Ciliophora (ciliate)
Each cell usually has at least 1 macronucleus and 1 or more micronuclei; most have a cytostome; all cilia arranged in an ordered fashion at some time in the life cycle; divide by homothetogenic binary fission, but some sedentary forms produce swarmers during asexual reproduction; sexual reproduction is universal in ciliates and normally involves conjugation between 2 cells; more than 7,000 described species, of which the majority are free-living.
Subphylum Postciliodesmatophora
Class Karyorelictea
Long, wormlike ciliates that may be extremely contractile; in some, 1 of the surfaces may be devoid of cilia; 2 to many macronuclei; includes 4 orders; representative genera include Protocruzia, Trachelonema, Loxodes, and Geleia.
Class Spirotrichea
Conspicuous right and left oral or preoral ciliature; cytostome may be deep or shallow, and some species live inside loricae (tubes secreted by themselves or an agglomerate of material); lorica may be attached to a substrate; 3 subclasses, Heterotrichia, Choreotrichia (including the loricate tintinnid ciliates and the oligotrichs), and Stichotrichia.
Subphylum Rhabdophora
Class Prostomatea
Cytostome apical to subapical, with a shallow cytostomal cavity; oral kinetids may be tangential to the perimeter of the oral area; includes Coleps and Tiarina.
Class Litostomatea
Simple oral ciliature may have oral toxicysts; includes Dileptus and Lacrymaria.
Subphylum Cyrtophora
Class Phyllopharyngea
Where the feeding stage is sessile, the dispersal stage has simple ciliature; includes suctorians, which lack cilia in the trophic stage and feed via tentacles, and the chonotrichs, which are usually sessile ectosymbionts of crustaceans.
Class Nassophorea
Often well-developed oral/cytopharyngeal zones; typical genera are Nassula, Paramecium, Frontonia, and hypotrichs, such as Euplotes.
Class Oligohymenophorea
Complex oral cilia distinct from the body cilia; oral cavity may be deep, with ciliary structures extending into it; includes Tetrahymena and Vorticella.
Class Colpodea
Some inhabit gelatinous sheaths (Mycterothrix tuamotuensis); includes Woodruffia and Colpoda.
Additional Reading
John J. Lee, Seymour H. Hutner, and Eugene C. Bovee (eds.), An Illustrated Guide to the Protozoa (1985), includes sections on the different groups, with many excellent illustrations. Michael A. Sleigh, Protozoa and Other Protists (1989), covers the features and physiology of the major groups, with a section on symbiosis and ecology, and is well illustrated. Coverage of the ecology and physiology of ciliates, sarcodines, and flagellates may be found in Johanna Laybourn-Parry, A Functional Biology of Free-Living Protozoa (1984); and Tom Fenchel, The Ecology of Protozoa (1987). Richard R. Kudo, Protozoology, 5th ed. (1966, reissued 1971), although slightly out-of-date, is nonetheless a useful book covering the major groups in detail. Michael Levandowsky and Seymour H. Hutner (eds.), Biochemistry and Physiology of Protozoa, 2nd ed., 4 vol. (1979–81), contains articles on all aspects of protozoan physiology and biochemistry, including structure, locomotion, metabolism, mating behaviour, and immunology of parasitic protozoan diseases. J.P. Kreier and J.R. Baker, Parasitic Protozoa (1987), is an easy-to-read overview covering physiology, biochemistry, and life cycles. R.S. Phillips, Malaria (1983), a small, easy-to-read book, discusses pathology, immunology, and control methods. John R. Haynes, Foraminifera (1981), covers the fossil record and the importance of these sarcodines, with many illustrations. O. Roger Anderson, Radiolaria (1983), covers the biology of this marine group. F.J.R. Taylor (ed.), The Biology of Dinoflagellates (1987), contains articles on the physiology and other aspects of these flagellates. Lynda J. Goff (ed.), Algal Symbiosis (1983), contains articles on endosymbionts of some protozoans and their role. John O. Corliss, The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature, 2nd ed. (1979), is a specialized text providing further references. Lynn Margulis, Symbiosis and Cell Evolution: Life and Its Environment on the Early Earth (1981), is an interesting text on the evolution of cellular structures.
- sitar
- sitatunga
- sitella
- Sites of the modern Olympic Games
- Sithole, Ndabaningi
- sit-in
- Sitka
- Sitka National Historical Park
- Sitrah
- Sittang River
- Sittard
- Sitte, Camillo
- Sitter, Willem de
- Sitting Bull
- Sittwe
- situated approach artificial intelligence
- Sitwell, Dame Edith
- Sitwell, Sir Osbert, 5th Baronet
- Sitwell, Sir Sacheverell, 6th Baronet
- Siuri
- Siva Dayāl Sāheb
- Sivaji Ganesan
- Sivapithecus
- Sivas
- Siwalik Range