angiosperm
plant
Introduction
any member of the more than 300,000 species of flowering plants (division Anthophyta), the largest and most diverse group within the kingdom Plantae. Angiosperms represent approximately 80 percent of all the known green plants now living. The angiosperms are vascular seed plants in which the ovule (egg) is fertilized and develops into a seed in an enclosed hollow ovary. The ovary itself is usually enclosed in a flower, that part of the angiospermous plant that contains the male or female reproductive organs or both. Fruits are derived from the maturing floral organs of the angiospermous plant and are therefore characteristic of angiosperms. By contrast, in gymnosperms (gymnosperm) (e.g., conifers), the other large group of vascular seed plants, the seeds do not develop enclosed within an ovary but are usually borne exposed on the surfaces of reproductive structures, such as cones, that originally produced the spores.
Unlike such nonvascular plants as the bryophytes (bryophyte), in which all cells in the plant body participate in every function necessary to support, nourish, and extend the plant body (e.g., nutrition, photosynthesis, and cell division), angiosperms have evolved (evolution) specialized cells and tissues that carry out these functions and have further evolved specialized vascular tissues that translocate the water and nutrients to all areas of the plant body. The specialization of the plant body, which has evolved as an adaptation to a principally terrestrial habitat, includes extensive root systems that anchor the plant and absorb water and minerals from the soil; a stem that supports the growing plant body; and leaves, which are the principal sites of photosynthesis for most angiospermous plants. Another significant evolutionary advancement over the nonvascular and the more primitive vascular plants is the presence of localized regions for plant growth, called meristems (meristem) and cambia (cambium), which extend the length and width of the plant body, respectively. Except under certain conditions, these regions are the only areas in which cell division takes place in the plant body, although cell differentiation continues to occur over the life of the plant.
The angiosperms dominate the Earth's surface and vegetation in more environments, particularly terrestrial habitats, than any other group of plants. As a result, angiosperms are the most important ultimate source of food for birds and mammals, including humans. In addition, the flowering plants are the most economically important group of green plants, serving as a source of pharmaceuticals, fibre products, timber, ornamentals, and other commercial products.
Although the taxonomy of the angiosperms is still incompletely known, the latest classification system incorporates a large body of comparative data derived from studies of DNA sequences. It is known as the Angiosperm Phylogeny Group II (APG II) botanical classification system. The angiosperms came to be considered a group at the division level (comparable to the phylum level in animal classification systems) called Anthophyta.
Throughout this article the orders or families are given, usually parenthetically, following the vernacular or scientific name of a plant. Following taxonomic conventions, genera and species are italicized. The higher taxa are readily identified by their suffixes: families end in -aceae and orders in -ales.
For a comparison of angiosperms with the other major groups of plants, see plant, bryophyte, fern, lower vascular plant, and gymnosperm.
General features



 The variety of forms found among angiosperms is greater than that of any other plant group. The size range alone is quite remarkable, from the smallest individual flowering plant, probably the watermeal (Wolffia; Araceae) at less than 2 millimetres (0.08 inch), to one of the tallest angiosperms, Australia's mountain ash tree (Eucalyptus regnans; Myrtaceae) at about 100 metres (330 feet). Between these two extremes lie angiosperms of almost every size and shape. Examples of this variability include the succulent cacti (Cactaceae), the fragile orchids (Orchidaceae; see photograph-->), the baobab (Adansonia digitata; Malvaceae), vines, rosette plants such as dandelion (see photograph-->), and carnivorous plants such as sundews (Drosera; Droseraceae 【see photograph-->】) and Venus's-flytrap (Dionaea muscipula; Droseraceae 【see photograph-->】). To understand this vast array of forms, it is necessary to consider the basic structural plan of the angiosperms.
The variety of forms found among angiosperms is greater than that of any other plant group. The size range alone is quite remarkable, from the smallest individual flowering plant, probably the watermeal (Wolffia; Araceae) at less than 2 millimetres (0.08 inch), to one of the tallest angiosperms, Australia's mountain ash tree (Eucalyptus regnans; Myrtaceae) at about 100 metres (330 feet). Between these two extremes lie angiosperms of almost every size and shape. Examples of this variability include the succulent cacti (Cactaceae), the fragile orchids (Orchidaceae; see photograph-->), the baobab (Adansonia digitata; Malvaceae), vines, rosette plants such as dandelion (see photograph-->), and carnivorous plants such as sundews (Drosera; Droseraceae 【see photograph-->】) and Venus's-flytrap (Dionaea muscipula; Droseraceae 【see photograph-->】). To understand this vast array of forms, it is necessary to consider the basic structural plan of the angiosperms.The basic angiosperm form is woody or herbaceous. Woody forms (generally trees and shrubs) are rich in secondary tissues, while herbaceous forms (herbs) rarely have any. Annuals (annual) are herbs that complete their growing cycle (growth, flowering, and death) within the same season. Examples of annuals can be found among cultivated garden plants, such as beans (Phaseolus; Fabaceae), corn (maize; Zea mays; Poaceae), and squashes (Cucurbita; Cucurbitaceae), as well as among the wildflowers, such as some buttercups (Ranunculus) and larkspurs (Delphinium). Biennials (biennial) are also herbs, but, unlike annuals, their growing cycle spans two years: the vegetative (nonreproductive) plant growth takes place from seed during the first year, and flowers and fruit develop during the second. The beet (Beta vulgaris; Amaranthaceae) and wild carrot (Daucus carota; Apiaceae) are well-known biennials.
A perennial grows for many years and often flowers annually. In temperate areas the aerial parts of a perennial die back to the ground at the end of each growing season and new shoots are produced the following season from such subterranean parts as bulbs, rhizomes, corms, tubers, and stolons.
Structure
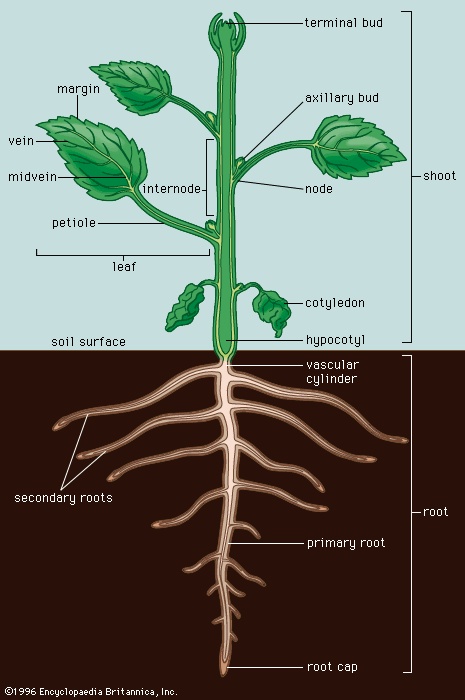 The basic angiosperm body has three parts: roots (root), stems, and leaves (Figure 1-->). These primary organs constitute the vegetative (nonreproductive) plant body. Together, the stem and its attached leaves constitute the shoot. Collectively, the roots of an individual plant make up the root system and the shoots the shoot system.
The basic angiosperm body has three parts: roots (root), stems, and leaves (Figure 1-->). These primary organs constitute the vegetative (nonreproductive) plant body. Together, the stem and its attached leaves constitute the shoot. Collectively, the roots of an individual plant make up the root system and the shoots the shoot system.Root systems
The roots anchor a plant, absorb water and minerals, and provide a storage area for food. The two basic types of root systems are a primary root system and an adventitious root system. The most common type, the primary system, consists of a taproot (primary root) that grows vertically downward (positive geotropism). From the taproot are produced smaller lateral roots (secondary roots) that grow horizontally or diagonally. These secondary roots further produce their own smaller lateral roots (tertiary roots). Thus, many orders of roots of descending size are produced from a single prominent root, the taproot. Most dicotyledons (dicotyledon) produce taproots, as, for example, the dandelion (Taraxacum officinale).
In some cases, the taproot system is modified into a fibrous, or diffuse, system, in which the initial secondary roots soon equal or exceed the primary root in size. The result is several large, positively geotropic roots that produce higher-order roots, which may also grow to the same size. Thus, in fibrous root systems there is no well-defined single taproot. In general, fibrous root systems are shallower than taproot systems.
The second type of root system, the adventitious root system, differs from the primary variety in that the primary root is short-lived and is replaced within a short time by many roots that form from the stem. Most monocotyledons have adventitious roots; examples include orchids, bromeliads, and many other epiphytic plants in the tropics. Grasses (grass) (family Poaceae) and many other monocotyledons produce fibrous root systems with the development of adventitious roots.
Many primary root and adventitious root systems have become modified for special functions, the most common being the formation of tuberous (fleshy) roots for food storage. For example, carrots (carrot) and beets (beet) are tuberous roots that are modified from taproots, and cassava (manioc) is a tuberous root that is modified from an adventitious root. (Tubers, on the other hand, are modified, fleshy, underground stems and will be discussed below.)
 Adventitious roots, when modified for aerial support, are called prop roots, as in corn or some figs (fig) ( Ficus; Moraceae). In many tropical rain forest trees, large woody prop roots develop from adventitious roots on horizontal branches and provide additional anchorage and support. Many bulbous (bulb) plants have contractile adventitious roots that pull the bulb deeper into the ground as it grows. Climbing plants often grip their supports with specialized adventitious roots. Some lateral roots of mangroves (mangrove) become specialized as pneumatophores in saline mud flats; pneumatophores are lateral roots that grow upward (negative geotropism) for varying distances and function as the site of oxygen intake for the submerged primary root system (see photograph-->). The plants mentioned above are only a few examples of root diversity in angiosperms, a condition that is unparalleled in any other vascular plant group.
Adventitious roots, when modified for aerial support, are called prop roots, as in corn or some figs (fig) ( Ficus; Moraceae). In many tropical rain forest trees, large woody prop roots develop from adventitious roots on horizontal branches and provide additional anchorage and support. Many bulbous (bulb) plants have contractile adventitious roots that pull the bulb deeper into the ground as it grows. Climbing plants often grip their supports with specialized adventitious roots. Some lateral roots of mangroves (mangrove) become specialized as pneumatophores in saline mud flats; pneumatophores are lateral roots that grow upward (negative geotropism) for varying distances and function as the site of oxygen intake for the submerged primary root system (see photograph-->). The plants mentioned above are only a few examples of root diversity in angiosperms, a condition that is unparalleled in any other vascular plant group.Stems (stem)
The stem is an aerial axis of the plant that bears leaves and flowers and conducts water and minerals from the roots and food from the site of synthesis to areas where it is to be used. The main stem of a plant is continuous with the root system through a transition region called the hypocotyl. In the developing embryo, the hypocotyl is the embryonic axis that bears the seedling leaves (cotyledons).
In a maturing stem, the area where a leaf attaches to the stem is called a node, and the region between successive nodes is called an internode. Stems bear leafy shoots (branches) at the nodes, which arise from buds (bud) (dormant shoots). Lateral branches develop either from axillary, or lateral, buds found in the angle between the leaf and the stem or from terminal buds at the end of the shoot. In temperate-climate plants these buds have extended periods of dormancy, whereas in tropical plants the period of dormancy is either very short or nonexistent.
The precise positional relationship of stem, leaf, and axillary bud is important to understanding the diversity of the shoot system in angiosperms. Understanding this relationship makes it possible to identify organs such as leaves that are so highly modified they no longer look like leaves, or stems that are so modified that they resemble leaves.
Branching in angiosperms may be dichotomous or axillary. In dichotomous branching, the branches form as a result of an equal division of a terminal bud (i.e., a bud formed at the apex of a stem) into two equal branches that are not derived from axillary buds, although axillary buds are present elsewhere on the plant body. The few examples of dichotomous branching among angiosperms are found only in some cacti, palms, and bird-of-paradise plants.
The two modes of axillary branching in angiosperms are monopodial and sympodial. Monopodial branching occurs when the terminal bud continues to grow as a central leader shoot and the lateral branches remain subordinate—e.g., beech trees (Fagus). Sympodial branching occurs when the terminal bud ceases to grow (usually because a terminal flower has formed) and an axillary bud or buds become new leader shoots, called renewal shoots—e.g., the Joshua tree (Yucca brevifolia). Plants with monopodial growth are usually pyramidal in overall shape, while those with sympodial growth often resemble a candelabra.
By combining monopodial and sympodial branching in one plant, many different tree architectures have evolved. A simple example is found in dogwoods (dogwood) (Cornus), where the main axis is monopodial and the lateral branches are sympodial.
Very different plant forms result from simply changing the lengths of the internodes. Extreme shortening of the internodes results in rosette plants, such as lettuce, Lactuca sativa (Asteraceae), in which the leaves develop but the internodes between them do not elongate. Extreme lengthening of the internodes often results in twining vines, as in the yam, Dioscorea esculenta (Dioscoreaceae).
Leaves (leaf)
The basic angiosperm leaf is composed of a leaf base, two stipules, a petiole, and a blade (lamina). The leaf base is the slightly expanded area where the leaf attaches to the stem. The paired stipules, when present, are located on each side of the leaf base and may resemble scales, spines, glands, or leaflike structures. The petiole is a stalk that connects the blade with the leaf base. The blade is the major photosynthetic surface of the plant and appears green and flattened in a plane perpendicular to the stem.
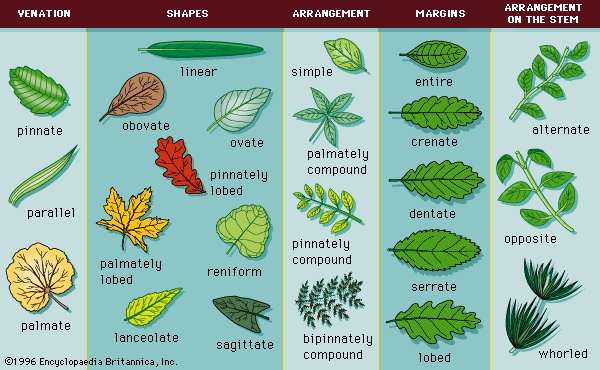 When only a single blade is inserted directly on the petiole, the leaf is called simple (Figure 2-->). Simple leaves may be variously lobed along their margins. The margins of simple leaves may be entire and smooth or they may be lobed in various ways. The coarse teeth of dentate margins project at right angles, while those of serrate margins point toward the leaf apex. Crenulate margins have rounded teeth or scalloped margins. Leaf margins of simple leaves may be lobed in one of two patterns, pinnate or palmate. In pinnately lobed margins the leaf blade (lamina) is indented equally deep along each side of the midrib (as in the white oak, Quercus alba), and in palmately lobed margins the lamina is indented along several major veins (as in the red maple, Acer rubrum). A great variety of base and apex shapes also are found.Many leaves contain only some of these leaf parts; for example, many leaves lack a petiole and so are attached directly to the stem (sessile), and others lack stipules (exstipulate). In compound leaves (Figure 2-->), a blade has two or more subunits called leaflets: in palmately compound leaves, the leaflets radiate from a single point at the distal end of the petiole; in pinnately compound leaves, a row of leaflets forms on either side of an extension of the petiole called the rachis. Some pinnately compound leaves branch again, developing a second set of pinnately compound leaflets (bipinnately compound). The many degrees of compoundness in highly elaborated leaves, such as bipinnately or tripinnately compound, cause these leaves to often appear to be shoot systems. It is always possible to distinguish them, however, because axillary buds are found in the angle between the stem and the petiole (axil) of pinnately or palmately compound leaves but not in the axils of leaflets.
When only a single blade is inserted directly on the petiole, the leaf is called simple (Figure 2-->). Simple leaves may be variously lobed along their margins. The margins of simple leaves may be entire and smooth or they may be lobed in various ways. The coarse teeth of dentate margins project at right angles, while those of serrate margins point toward the leaf apex. Crenulate margins have rounded teeth or scalloped margins. Leaf margins of simple leaves may be lobed in one of two patterns, pinnate or palmate. In pinnately lobed margins the leaf blade (lamina) is indented equally deep along each side of the midrib (as in the white oak, Quercus alba), and in palmately lobed margins the lamina is indented along several major veins (as in the red maple, Acer rubrum). A great variety of base and apex shapes also are found.Many leaves contain only some of these leaf parts; for example, many leaves lack a petiole and so are attached directly to the stem (sessile), and others lack stipules (exstipulate). In compound leaves (Figure 2-->), a blade has two or more subunits called leaflets: in palmately compound leaves, the leaflets radiate from a single point at the distal end of the petiole; in pinnately compound leaves, a row of leaflets forms on either side of an extension of the petiole called the rachis. Some pinnately compound leaves branch again, developing a second set of pinnately compound leaflets (bipinnately compound). The many degrees of compoundness in highly elaborated leaves, such as bipinnately or tripinnately compound, cause these leaves to often appear to be shoot systems. It is always possible to distinguish them, however, because axillary buds are found in the angle between the stem and the petiole (axil) of pinnately or palmately compound leaves but not in the axils of leaflets.The three patterns of leaf arrangement on stems in angiosperms are alternate, opposite (paired), and whorled. In alternate-leaved plants, the leaves are single at each node and borne along the stem alternately in an ascending spiral. In opposite-leaved plants, the leaves are paired at a node and borne opposite to each other. A plant has whorled leaves when there are three or more equally spaced leaves at a node.
Leaf modifications
Whole leaves or parts of leaves are often modified for special functions, such as for climbing and substrate attachment, storage, protection against predation or climatic conditions, or trapping and digesting insect prey. In temperate trees, leaves are simply protective bud scales; in the spring when shoot growth is resumed, they often exhibit a complete growth series from bud scales to fully developed leaves.
Stipules often develop before the rest of the leaf; they protect the young blade and then are often shed when the leaf matures. Spines are also modified leaves. In cacti (cactus), spines are wholly transformed leaves that protect the plant from herbivores, radiate heat from the stem during the day, and collect and drip condensed water vapour during the cooler night. In the many species of the spurge family ( Euphorbiaceae), the stipules are modified into paired stipular spines and the blade develops fully. In ocotillo (Fouquieria splendens; Fouquieriaceae), the blade falls off and the petiole remains as a spine.
Many desert plants, such as stoneplants (Lithops; Aizoaceae) and aloe (Aloe; Asphodelaceae), develop succulent leaves for water storage. The most common form of storage leaves are the succulent leaf bases of underground bulbs (e.g., tulip and Crocus) that serve as either water- or food-storage organs or both. Many nonparasitic plants that grow on the surfaces of other plants (epiphytes (epiphyte)), such as some of the bromeliads (bromeliad), absorb water through specialized hairs on the surfaces of their leaves. In the water hyacinth (Eichhornia crassipes), swollen petioles keep the plant afloat.
 Leaves or leaf parts may be modified to provide support. Tendrils (tendril) and hooks are the most common of these modifications. In the flame lily (Gloriosa superba; Colchicaceae), the leaf tip of the blade elongates into a tendril and twines around other plants for support. In the garden pea (Pisum sativum; Fabaceae), the terminal leaflet of the compound leaf develops as a tendril. In nasturtium (Tropaeolum majus; Tropaeolaceae) and Clematis (Ranunculaceae), the petioles coil around other plants for support. In catbrier (Smilax; Smilacaceae 【see photograph-->】), the stipules function as tendrils. In certain vining angiosperms with compound leaves, some of the leaflets have modified into grapnellike hooks—e.g., Tecoma radicans. Many monocotyledons have sheathing leaf bases that are concentrically arranged and form a pseudotrunk, as in banana (Musa). In many epiphytic bromeliads, the pseudotrunk also functions as a water reservoir.
Leaves or leaf parts may be modified to provide support. Tendrils (tendril) and hooks are the most common of these modifications. In the flame lily (Gloriosa superba; Colchicaceae), the leaf tip of the blade elongates into a tendril and twines around other plants for support. In the garden pea (Pisum sativum; Fabaceae), the terminal leaflet of the compound leaf develops as a tendril. In nasturtium (Tropaeolum majus; Tropaeolaceae) and Clematis (Ranunculaceae), the petioles coil around other plants for support. In catbrier (Smilax; Smilacaceae 【see photograph-->】), the stipules function as tendrils. In certain vining angiosperms with compound leaves, some of the leaflets have modified into grapnellike hooks—e.g., Tecoma radicans. Many monocotyledons have sheathing leaf bases that are concentrically arranged and form a pseudotrunk, as in banana (Musa). In many epiphytic bromeliads, the pseudotrunk also functions as a water reservoir.Insectivorous (carnivorous (carnivorous plant)) plants use their leaves to attract and trap insects. Glands in the leaves secrete enzymes that digest the captured insects, and the leaves then absorb the nitrogenous compounds (amino acids) and other products of digestion. Plants that use insects as a nitrogen source tend to grow in nitrogen-deficient soils.
Shoot system modifications
Entire shoot systems are often modified for such special functions as climbing, protection, adaptation to arid habitats, and water or food storage. The modifications generally involve structural and shape changes to the stem and the reduction of the leaves to small scales. Many of the modifications parallel those previously described for leaves. In the passion flower (Passiflora; Passifloraceae) and grape (Vitis vinifera; Vitaceae), axillary buds develop as tendrils with reduced leaves and suppressed axillary buds. In the grape these axillary tendrils are actually modified and reduced inflorescences. In the plant from which strychnine is obtained (Strychnos nux-vomica), the axillary buds develop into hooks for climbing. The tendrils of ivy (Hedera helix; Apiales) produce enlarged cuplike holdfasts.
Thorns represent the modification of an axillary shoot system in which the leaves are reduced and die quickly and the stems are heavily sclerified and grow for only a limited time (determinate growth). Thorns appear to protect the plant against herbivores. Examples are found in the Bougainvillea (Nyctaginaceae), where the thorn is a modified inflorescence, the honey locust (Gleditsia triacanthos; Fabaceae), the anchor plant (Colletia paradoxa; Rhamnaceae), and Citrus (Rutaceae).
 Cladodes (also called cladophylls or phylloclades) are shoot systems in which leaves do not develop; rather, the stems become flattened and assume the photosynthetic functions of the plant. In Asparagus (Asparagus officinalis; Asparagaceae), the scales found on the asparagus spears are the true leaves. If the thick, fleshy asparagus spears continue to grow, flat, green, leaflike structures called cladodes develop in the axils of the scale leaves. The presence of cladodes in unrelated desert angiosperm families is an excellent example of convergent evolution, or the independent development of the same characteristic in unrelated taxa.
Cladodes (also called cladophylls or phylloclades) are shoot systems in which leaves do not develop; rather, the stems become flattened and assume the photosynthetic functions of the plant. In Asparagus (Asparagus officinalis; Asparagaceae), the scales found on the asparagus spears are the true leaves. If the thick, fleshy asparagus spears continue to grow, flat, green, leaflike structures called cladodes develop in the axils of the scale leaves. The presence of cladodes in unrelated desert angiosperm families is an excellent example of convergent evolution, or the independent development of the same characteristic in unrelated taxa.All cacti (Cactaceae) have cladodes, and many desert members of the spurge (Euphorbiaceae) and milkweed ( Apocynaceae) families have similar vegetative morphologies that are derived by modifying different parts to look and function in the same way. Each of these plant groups has columnar, water-storing green stems, reduced leaves, and protective spines or thorns. They are often called stem succulents. In the cacti, the leaves on the main stems last for a very short time (they do not even develop as scale leaves) and the leaves of the axillary buds (the round cushion areas, or areoles, on the trunks) develop as spines. In the Euphorbiaceae, the leaves on the main stems are green but short-lived, and the stipules develop as spines. In the Apocynaceae the leaves are also small and ephemeral, and the axillary buds develop as thorns. The cacti are New World plants adapted to dry or arid habitats, and the Euphorbiaceae and Apocynaceae occur in similar habitats in Asia and Africa. The reduction of leaves is so extreme in the Cactaceae that the epiphytic cacti (e.g., Epiphyllum (leaf cactus)) of the Neotropics can no longer produce leaves; rather, they produce thin, flat cladodes that superficially resemble leaves.
Many shoot systems have been modified into organs of food storage, reproduction, or both, called rhizomes (rhizome), tubers, and corms. Rhizomes are distinguished from roots in having nodes with reduced leaves and internodes. Rhizomes are horizontal, usually subterranean shoots with scale leaves and adventitious roots on the underside. Their chief functions are vegetative reproduction and food storage; food stored in the rhizomes allows these plants to survive drought and extended winters. Most rhizomes are perennial, sending up new shoots from the nodes and spreading the colony. Often the terminal bud of a rhizome becomes upright and then flowers, with a rhizome axillary bud becoming a renewal shoot. Many economically important plants, such as banana, and almost all grasses, including bamboo, and sugarcane, have rhizomes. Such plants are propagated primarily by fragmentation of the rhizome. In some plants, the growing tips of rhizomes become much enlarged food storage organs called tubers (tuber). The common potato (Solanum tuberosum) forms such tubers. The much-reduced scale leaves and their associated axillary buds form the eyes of the potato. Tubers should not be confused with tuberous roots. Tubers are modified shoots, whereas tuberous roots are modified roots. The common feature, and hence the similar names, derives from the fleshy nature of both organs. Tubers and tuberous roots function in water and food storage, but only tubers are involved in vegetative (nonsexual) reproduction. Tuberous roots develop from taproots in carrots and from adventitious roots in dahlias (Dahlia; Asteraceae).
Another distinctive modification for food storage is the corm, a short, upright shoot system with a thick, hard stem covered with thin membranous scale leaves as in jack-in-the-pulpit (Arisaema triphyllum; Araceae) and gladiolus (Gladiolus; Iridaceae). Corms are usually hard and fibrous and function for overwintering and drought resistance.
Slender creeping stems that grow above the soil surface are called stolons, or runners. Stolons have scale leaves and can develop roots and, therefore, new plants, either terminally or at a node. In the strawberry (Fragaria), the stolons are used for propagation; buds appear at nodes along the stolons and develop into new strawberry plants.
Distribution and abundance
The diversity of form within the angiosperms has contributed to their successful colonization of more habitats than any other group of land plants. Gymnosperms (gymnosperm) (the nonflowering seed plants) are only woody plants with a few woody twining vines. There are few herbaceous or aquatic gymnosperms; most gymnosperms do not occur in mangrove (swampy) vegetation or marine habitats. With the exception of cycads, gymnosperms have simple leaves, and none are modified as spines, tendrils, or storage organs.
The absence of substantial diversity in the vegetative features of gymnosperms appears to have limited their ability to adapt to diverse or extreme habitats. The absence of vessels (vessel) in most gymnosperms, and hence the less efficient water transport system than that found in the angiosperms, is one example. In fact, the only gymnosperms with vessels, the Gnetales, is the only group that contains vines and the only group that deviates from the usually woody trunk growth form. The absence of vessels in angiosperms, however, is rare; the few groups without vessels are small trees or shrubs with limited distribution, as in the Winteraceae. Another factor contributing to the limited distribution of gymnosperms is that they do not produce reproductive structures until several years after the seed germinates; therefore, a woody habit is required to achieve sexual maturity. Finally, the gymnosperms also require a relatively stable environment for growth. Thus, restraints imposed by anatomy and life cycle have probably limited morphological diversity among the gymnosperms.
The wide variation in the angiosperm form is reflected in the range of habitats in which they grow and their almost complete worldwide distribution. The only area without angiosperms is the southern region of the Antarctic continent, although two angiosperm groups are found in the islands off that continent. Angiosperms dominate terrestrial vegetation, particularly in the tropics, although submerged and floating aquatic angiosperms do exist throughout the world. Angiosperms are the principal component of salt marshes, tidal marshes, and mangrove marshes. The only vascular marine plants are a few submerged marine angiosperms that occur in shallow waters of coastal areas throughout the world—for example, the sea grasses (Zostera and Cymodocea; Cymodoceaceae). The various terrestrial biomes (defined primarily based upon the type of vegetation and climate) are composed mainly of herbaceous and woody angiosperms, except for taiga (boreal forest), temperate rain forest, and juniper savanna, where conifers (a gymnospermous division) dominate the woody component and angiosperms dominate the herbaceous and shrub components.
Morphological and habitat diversity, together with cosmopolitan distribution, contributes to the wide ecological tolerance of the angiosperms—adapting to Alpine tundra regions and salt marshes, from the Arctic Circle to the lowland tropical rain forests. The importance of angiosperms in the terrestrial portion of the biosphere is rarely rivaled by any other group of organisms.
All but a few angiosperms are autotrophs: they are green plants (primary producers) that use solar radiation, carbon dioxide, water, and minerals to synthesize organic compounds; oxygen is a by-product of these metabolic reactions. The few exceptions are either saprophytes (e.g., the Indian pipe Monotropa uniflora; Ericaceae) that use connections with mycorrhizal (mycorrhiza) fungi (fungi that form an association with the roots of certain plants) to obtain carbohydrates from dead organic material or parasites (parasitism) that develop specialized roots (haustoria (haustorium)), which penetrate the host plant and absorb food and other materials (e.g., the dodder 【Cuscuta salina; Convolvulaceae】).
Importance
Contribution to food chain
Because angiosperms are the most numerous component of the terrestrial environment in terms of biomass and number of individuals, they provide an important source of food (food chain) for animals and other living organisms. Organic compounds (organic compound) (carbon-containing compounds, principally carbohydrates) not only are used by the plant itself for synthesizing cellular structures and for fueling their basic metabolisms but also serve as the only source of energy for most heterotrophic organisms. (Heterotrophs require an organic source of carbon that has originated as part of another living organism, in contrast to autotrophs, which require only an inorganic source of carbon—CO2.) solar energy is trapped by the photosynthetic pigments in the plant cells and converted into chemical energy, which is stored in the tissues of the plant. The trapped energy is transferred from one organism to the next as herbivores consume the plant, carnivores consume herbivores, and so on up the food chain. In a temperate forest, one angiosperm tree supports many thousands of animals (the majority being insects, birds, and mammals), a relationship that underscores the basic importance of the angiosperms to the food chain.
The angiosperm body contributes to the food chain in many ways. The vegetative parts (the nonreproductive organs, such as stems and leaves) are consumed by, and support, plant-eating animals. Vast numbers of insects and other invertebrates depend on shoots for food during all or part of their life histories. The reproductive organs (flowers (flower), fruits, and seeds) also provide an energy source for many animals. The pollen supports many pollinating insects, particularly bees.
The flowers provide food from floral nectaries that secrete sugars and amino acids. These flowers often produce fragrances that attract pollinators which feed on the nectar. Nectar-feeding animals include many insect groups (bees, butterflies, moths, flies, and even mosquitoes), many mammal groups (bats, small rodents, and small marsupials), and birds (honeyeaters, hummingbirds, and sunbirds). Nectaries also occur on the nonfloral, or vegetative, parts of some angiosperms, such as the leaves and the petioles of bull's-horn thorn (Acacia collinsii; Fabaceae). Ants live (mutualism) inside the hollow modified spinous structures of bull's-horn thorn and feed on the nectar. In return for this food source, they attack and destroy animals of all sizes as well as other plants that contact the acacia plant. In doing so, the ants protect the bull's-horn thorn from herbivores and other plants competing for the available space, light, and minerals.
Fruits produced by angiosperms are the principal food for many bats, birds, mammals, and even some fish. Seeds are also an important food source for many animals, particularly small rodents and birds. These animals often carry the fruits and seeds of the angiosperms they consume to new areas, where the angiosperms propagate.
Another aspect of angiosperm diversity is found in the production of secondary compounds, such as alkaloids, quinones, essential oils, and glycosides. Angiosperms have evolved a comprehensive array of unpalatable or toxic secondary plant compounds that protect the plants from foraging herbivores. Some insects, however, successfully store these secondary compounds in their tissues and use them as protection from predation.
As the principal component of the terrestrial biosphere, the angiosperm flora determines many features of the habitat, some of which are available food, aspects of the forest canopy, and grazing land. They supply nesting sites and materials for a wide range of birds and mammals, and they are the principal living spaces for many primates, reptiles, and amphibians. The tank bromeliad, which traps water in its crowns, provides a habitat for salamanders, frogs, and many aquatic insects and larvae. The animal inhabitants of the water-filled, insectivorous pitcher-plant leaves (Nepenthaceae) have adapted to the hostile environment of the leaves' digestive fluids.
Significance to humans
Angiosperms are as important to humans as they are to other animals. Angiosperms serve as the major source of food—either directly or indirectly through consumption by herbivores—and, as mentioned above, they are a primary source of consumer goods, such as building materials, textile fibres, spices, herbs, and pharmaceuticals.
Among the most important food plants on a global scale are cereals from the grass family (Poaceae); potatoes, tomatoes, eggplant, and red or chili peppers from the potato family (Solanaceae); legumes or beans (Fabaceae); pumpkins, melons, and gourds from the squash family (Cucurbitaceae); broccoli, cabbage, cauliflower, radish, and other vegetables from the mustard family (Brassicaceae, or Cruciferae); and almonds, apples, apricots, cherries, loquats, peaches, pears, raspberries, and strawberries from the rose family (Rosaceae). Members of many angiosperm families are used for food on a local level, such as ullucu (Ullucus tuberosus) in the Andes and cassava (Manihot esculenta) throughout the tropics. Tropical angiosperm trees are an important source of timber in the tropics and throughout the world.
The flowering plants have a number of uses as food, specifically as grains, sugars, vegetables, fruits, oils, nuts, and spices. In addition, plants and their products serve a number of other needs, such as dyes, fibres, timber, fuel, medicines, and ornamentals. Many plants serve more than one function. For example, the seeds of the kapok fruit (Ceiba pentandra; Malvaceae) yield a water-repellent fibre used in sound and thermal insulation and an edible oil used in cooking, lubricants, and soap; the oil cake is rich in protein and is fed to livestock; and the soft, light wood is used to make furniture and boats.
The angiospermous plant converts the energy of the sun into starch, the energy-rich storage form of sugar, and reserves it in the endosperm of the seed for the time when the seedling germinates and grows. Among the most economically important grains (cereal) throughout the world are corn, wheat (Triticum), rice (Oryza), barley (Hordeum), oats (Avena), sorghum (Sorghum), and rye (Secale), all members of the grass family, Poaceae.
Corn provides food for humans and domesticated animals, and its derivatives (e.g., cornstarch and corn oil) are used in making cosmetics, adhesives, varnishes, paints, soaps, and linoleum. Among the many cultivars of Zea mays, dent corn, variety indentat, is a widely used feed type in the United States. Wheat, barley, and rye are all members of the same tribe (Triticeae) within the family Poaceae. Wheat is among the oldest of the cultivated food crops. Barley is used for human consumption, livestock feed, and malting. Rye is usually used as a livestock feed, but can be used in baking and distilling liquor. Rice is a semiaquatic annual grass and is one of the major cereal crops of the world.
Vegetables constitute perhaps the greatest diversity of form and nutritional content and are grown for one or more of their parts—the flowers, shoots, or leaves; or the underground parts, such as tuberous roots, bulbs, rhizomes, corms, and tubers.
The globe, or French, artichoke (Cynara scolymus; Asteraceae, also known as Compositae) is an immature flower bud and receptacle overlaid by bracts. Asparagus (Asparagus officinalis; Asparagaceae) is a perennial plant cultivated for its succulent green shoots (spears) that arise from underground stems called crowns.
The mustard family ( Brassicaceae, also known as Cruciferae) contains a number of important vegetables—broccoli, brussel sprouts, cabbage, cauliflower, collards, kale, and kohlrabi—all members of Brassica oleraceae and comprising a group of vegetables called the cole crops, a term that probably reflects the fact that they are principally stem plants. The flower heads and stalks of broccoli and cauliflower are eaten, the two plants differing in that the white head of the cauliflower consists of malformed (hypertrophied) flowers that form in dense clusters. Brussel sprouts continually form many small heads in the axils of the leaves throughout the growing season. The cabbage head is a large terminal bud.
The edible portion of celery (Apium graveolens; Apiaceae) is the petiole (leaf stalk) that arises from a compact stem. Rhubarb (Rheum rhabarbarum; Polygonaceae) is a leafy plant also grown for its leaf petioles.
Parsley (Petroselinum crispum; Apiaceae), spinach (Spinacia oleracea; Amaranthaceae), and Swiss chard (Beta vulgaris; Amaranthaceae) are cultivated for their leaves, and the leek (Allium ampeloprasum; Alliaceae), a close relative of the onion, is cultivated for its leaf bases.
Root crops are grown for their fleshy subterranean storage bodies: tuberous roots, bulbs, rhizomes, corms, and tubers. The potato is a tuber found in Solanaceae, the potato family. Other important root crops include the beet (Beta vulgaris; Amaranthaceae), the sweet potato (Ipomoea batatas; Convolvulaceae), and (in the family Brassicaceae) the radish (Raphanus sativus), turnip (Brassica rapa), and rutabaga (B. napus).
Bulb crops are underground leafy scales attached to short compressed stems; food is stored in the leaves rather than the roots, causing them to enlarge into bulbs. Onions and garlic (Allium cepa and A. sativum, respectively; Alliaceae) are the most obvious examples of the bulb vegetable.
Many plants classified popularly as vegetables are in actuality fruits because they develop from the reproductive structures of the plant. The genus Cucurbita (Cucurbitaceae) includes the pumpkins, squashes, and gourds, of which C. moschata (winter squash, or crookneck pumpkin), C. pepo (summer squash, or marrow), and C. mixta (the pumpkin, or mixta squash) are some of the common types. Breadfruit (Artocarpus altilis; Moraceae), a plant native to the Pacific Islands, is a staple, providing a rich source of calcium and starch.
The common bean (Phaseolus vulgaris), including the French, or kidney, bean, the string bean, and the navy bean, is the edible fleshy pod containing the bean seeds. It provides a good source of protein. Lima beans (P. lunatus) probably originated in Central America and are now found in the United States, the lowland tropics, and Africa.
The cucumber (Cucumis sativus; Cucurbitaceae) produces a fruit that develops from a branching vine. Okra (Abelmoschus esculentus; Malvaceae) is a warm-weather crop that produces small fruit pods.
The garden, or English, pea (Pisum sativum; Fabaceae, also known as Leguminosae) is an annual, cool-weather plant cultivated for its edible green seed or pod. The pea is found throughout most temperate and tropical regions.
The family Solanaceae contains the important fruit vegetables—eggplants (aubergines (eggplant)), peppers (pepper), and tomatoes (tomato)—all herbaceous plants, which are perennial in the tropics and annual in temperate zones. (The family also contains the potato, which is a root crop.) The eggplant (Solanum melongena) remains an important food crop in Asia.
The pepper (Capsicum; Solanaceae) includes the sweet, or bell, pepper (which is green when immature, but red or yellow when ripe), and the red, or chili, pepper. A native of Central and South America, this herbaceous plant is a perennial in the tropics and an annual in temperate zones. Pepper plants are cultivated for their fruits, some of which are extremely pungent owing to the presence of capsaicin found in the septa, in the placenta, and, to a lesser extent, in the seeds, but not in the wall, of the fruit.
The tomato (Solanum lycopersicum, formerly Lycopersicum esculentum; Solanaceae), native to South America, was at one time wrongly reported to bear poisonous fruits. The fruit is a fleshy berry invested with many small seeds.
Plants cultivated for their fruits are found in temperate, tropical, or subtropical regions. Temperate plants are generally deciduous and either tolerate or require a cool period for growth. Apples (Malus) and pears (Pyrus) are important pome fruits of the family Rosaceae. Some well-known stone fruits of the family include the peaches and nectarines (Prunus persica), plums (P. domestica), and cherries (P. avium). Other temperate fruits grown on bushes, vines, or low plants include the grapes (Vitis; Vitaceae), strawberry (Fragaria; Rosaceae), blueberries (Vaccinium), and cranberries (V. macrocarpon), both from Ericaceae.
Tropical fruits tend to be grown on evergreen plants and can survive temperatures only above freezing. Subtropical plants are either deciduous or tropical and are not as susceptible to temperatures slightly below freezing. Citrus (Rutaceae), avocados (Persea americana; Lauraceae), olives (Olea; Oleaceae), dates (Phoenix dactylifera; Arecaceae), fig (Ficus; Moraceae), pineapple (Ananas comosus; Bromeliaceae), banana (Musa; Muscaceae), and papaya (Carica; Caricaceae) are tropical and subtropical plants.
Commercially important plants cultivated for the nuts they produce are almonds (Prunus dulcis; Rosaceae), walnuts (Juglans regia; Juglandaceae), pecans (Carya illinoinensis; Juglandaceae), macadamias (Macadamia; Proteaceae), and filberts (Corylus; Betulaceae).
Sugarcane (Saccharum officinurum; Poaceae) and sugar beet (Amaranthaceae) are rich sources of natural sugar.
Peanuts (Arachis) and soybeans (Glycine), both members of Fabaceae, the legume family, of the order Fabales, produce edible seeds that are important for their rich supply of protein or oil. Other plants rich in oil and important economically are the castor bean (Ricinus; Euphobiaceae), coconut (Cocos nucifera; Arecaceae), corn, cotton (Gossypium; Malvaceae), flax (Linum usitatissimum; Linaceae), olives, oil palm (Elaeis guineensis; Arecaceae), sesame (Sesamum; Pedaliaceae), and sunflowers (Helianthus; Asteraceae).
As noted earlier, some plants produce toxic secondary compounds for protection. Some of the secondary compounds produced by angiosperms are not toxic, however; in fact, many are found in herbs and spices (spice and herb)—for example, cloves, the dried flower buds of Syzygium aromaticum (Myrtaceae). The use of herbs and spices in cooking predates recorded history. Herbs are usually leaves or young shoots of nonwoody plants, although bay leaves and a few other leaves from woody plants are also considered herbs. Spices are the highly flavoured, aromatic parts of plants that are usually high in essential oil content. Spices are derived from roots, rhizomes, leaves, bark, seeds, fruits, and flower parts. The search for spices and alternative shipping routes for spices played a major role in world exploration in the 13th to 15th centuries. Many beverages are also derived from angiosperms; these include coffee (Coffea arabica; Rubiaceae), tea (Camellia sinensis; Theaceae), most soft drinks (e.g., root beer from the roots of Sassafras albidum; Lauraceae), and most alcoholic beverages (e.g., beer and whiskey from cereal grains and wine from grapes).
The angiosperms provide valuable pharmaceuticals (pharmaceutical). With the exception of antibiotics, almost all medicinals either are derived directly from compounds produced by angiosperms or, if synthesized, were originally discovered in angiosperms. This includes some vitamins (e.g., vitamin C, originally extracted from fruits); aspirin, originally from the bark of willows (Salix; Salicaceae); narcotics (e.g., opium and its derivatives from the opium poppy, Papaver somniferum; Papaveraceae); and quinine from Cinchona (Rubiaceae) bark. Some angiosperm compounds that are highly toxic to humans have proved to be effective in the treatment of certain forms of cancer, such as acute leukemia (vincristine from the Madagascar periwinkle, Catharanthus roseus; Apocynaceae), and of heart problems ( digitalis from foxglove, Digitalis purpurea; Plantaginaceae). Muscle relaxants derived from curare (Strychnos toxifera; Loganiaceae) are used during open-heart surgery.
The contribution of the angiosperms to biodiversity and habitat is so extremely important that human life is totally dependent on it. A significant loss of angiosperms would reduce the variety of food sources and oxygen supply in a habitat and drastically alter the amount and distribution of the world's precipitation. Many sources of food and medicine doubtless remain to be discovered in this group of vascular plants.
Structure and function
The wide diversity in the morphological features of the plant body has been discussed above. This section will outline the underlying structural (anatomic) diversity among angiosperms.
Vegetative structures
There are three levels of integrated organization in the vegetative plant body: organ, tissue system, and tissue. The organs of the plant—the roots, stems, and leaves—are composed of tissue systems (dermal tissue, ground tissue, and vascular tissue; see below Tissue systems (angiosperm)). The tissues of each of these systems are composed of cells of one or more types (parenchyma, collenchyma, and sclerenchyma; see below Tissue systems: Ground tissue (angiosperm)). Tissues composed of only one cell type and performing only one function are simple tissues, while those composed of more than one cell type and performing more than one function, such as support and conduction, are complex tissues. Xylem and phloem are examples of complex tissues.
The plant develops from a fertilized egg, called a zygote, which undergoes mitotic cell division to form an embryo—a simple multicellular structure of undifferentiated cells (i.e., those that have not developed into cells of a specific type)—and eventually a mature plant. The embryo consists of a bipolar axis that bears one or two cotyledons (cotyledon), or seed leaves; in most dicots (dicotyledon) the cotyledons contain stored food in the form of proteins, lipids, and starch, or they are photosynthetic and produce these products, whereas in most monocots and some dicots the endosperm stores the food and the cotyledons absorb the digested food. The embryos of dicotyledons have two seed leaves, while those of monocotyledons have only one.
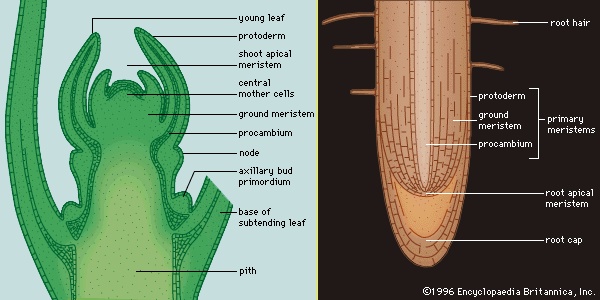 As the embryo continues to develop and new cells arise, the angiospermous plant develops specialized regions in which only cell division takes place and other areas in which nonreproductive (vegetative) activities, such as metabolism, respiration, and storage, occur. The areas of dividing cells, essentially permanently embryonic tissue, are called meristems, and their cells are termed initials. In the embryo they are found at either end of the bipolar axis and are called apical meristems. As the plant matures, apical meristems in the shoots produce new buds and leaves, and apical meristems in the roots are the points of active growth for roots (Figure 3-->). All growth produced by the apical meristems is primary growth and results in more primary tissues, which essentially extends the primary plant body.
As the embryo continues to develop and new cells arise, the angiospermous plant develops specialized regions in which only cell division takes place and other areas in which nonreproductive (vegetative) activities, such as metabolism, respiration, and storage, occur. The areas of dividing cells, essentially permanently embryonic tissue, are called meristems, and their cells are termed initials. In the embryo they are found at either end of the bipolar axis and are called apical meristems. As the plant matures, apical meristems in the shoots produce new buds and leaves, and apical meristems in the roots are the points of active growth for roots (Figure 3-->). All growth produced by the apical meristems is primary growth and results in more primary tissues, which essentially extends the primary plant body.After a cell in an apical meristem has divided mitotically, one of the two resulting daughter cells remains in the meristem as an initial cell, and the other cell is displaced into the plant body as a derivative cell. The displaced derivative cell may divide several times as it differentiates (changes in structure and physiology) from a meristemic cell into a mature cell, but only initial cells remain permanently in the apical meristem. However, although most permanently differentiated derivative cells are nondividing cells, and regions of division remain in the root and shoot apical meristems, there are regions of dividing derivative cells behind apical meristems that give rise to primary tissue systems and thus are also considered to be primary meristems.
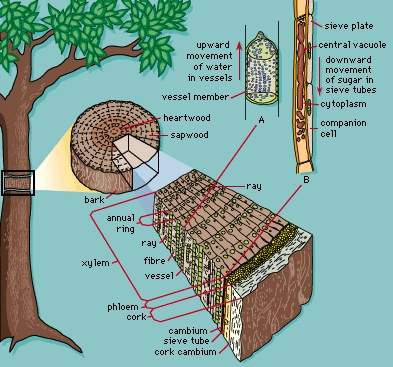
Three concentric regions of primary meristematic tissues develop immediately behind the apical meristem (Figure 3-->). These primary meristems produce the different tissues of the plant body: the outermost protoderm differentiates into the epidermis, a tissue that protects the plant; the adjacent ground meristem differentiates into the central ground tissues (the pith and cortex); and the procambium differentiates into the vascular tissues (the xylem, phloem, and vascular cambium). The xylem and phloem are conducting and supporting vascular tissues, and the vascular cambium is a lateral meristem that gives rise to the secondary vascular tissues, which constitute the secondary plant body.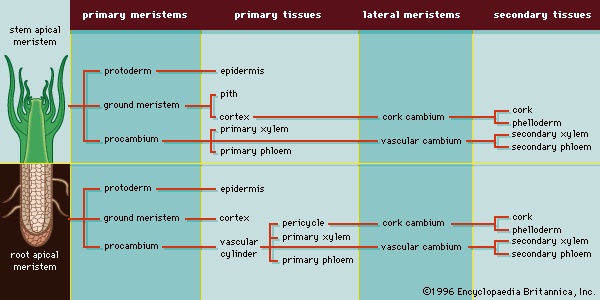 Lateral meristems, called cambia, run the length of the stems and roots of vascular plants and produce secondary tissues, which develop after a plant organ—or part of a plant organ—has ceased to elongate. Secondary growth is essentially an increase in girth. The vascular cambium produces secondary xylem and secondary phloem, and the cork cambium (phellogen) produces cork cells, from which the outer bark develops. Figure 4--> summarizes the patterns of primary and secondary growth from root and shoot apical meristems. (For a complete discussion of primary and secondary plant growth, see below Tissue systems: Vascular tissue (angiosperm).)
Lateral meristems, called cambia, run the length of the stems and roots of vascular plants and produce secondary tissues, which develop after a plant organ—or part of a plant organ—has ceased to elongate. Secondary growth is essentially an increase in girth. The vascular cambium produces secondary xylem and secondary phloem, and the cork cambium (phellogen) produces cork cells, from which the outer bark develops. Figure 4--> summarizes the patterns of primary and secondary growth from root and shoot apical meristems. (For a complete discussion of primary and secondary plant growth, see below Tissue systems: Vascular tissue (angiosperm).)Tissue systems
As mentioned above, three areas of meristematic tissue are derived directly from the apical meristem: the ground meristem, procambium, and protoderm. These meristematic tissues differentiate into the three primary tissues that constitute the primary plant body: ground tissue (pith and cortex), vascular tissue (xylem, phloem, and eventually the lateral, or secondary, meristem called the vascular cambium), and dermal tissue (epidermis), respectively.
Ground tissue
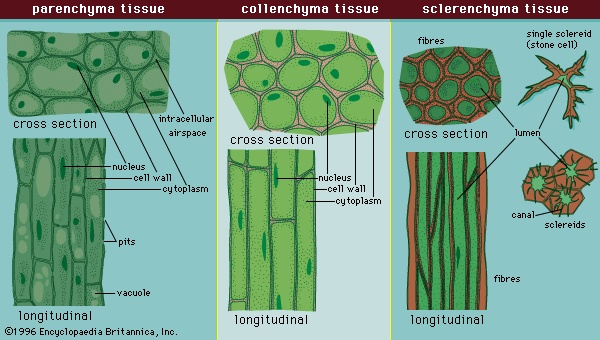 The ground tissue system arises from a ground tissue meristem and consists of three simple tissues: parenchyma, collenchyma, and sclerenchyma (Figure 5-->). The cells of each simple tissue bear the same name as their respective tissue.
The ground tissue system arises from a ground tissue meristem and consists of three simple tissues: parenchyma, collenchyma, and sclerenchyma (Figure 5-->). The cells of each simple tissue bear the same name as their respective tissue.Parenchyma, often the most common ground tissue, takes its name from the Greek para, meaning beside, and egchnma, meaning the contents of a pitcher (literally, something poured beside), indicating its ubiquitous nature throughout the plant body. It forms, for example, the cortex and pith of stems, the photosynthetic tissue layer within the epidermis of the leaves (mesophyll), the cortex of roots, the pulp of fruits, and the endosperm of seeds. Parenchyma is composed of relatively simple, undifferentiated parenchyma cells. In most plants, metabolic activity (such as respiration, digestion, and photosynthesis) occurs in these cells because they, unlike many of the other types of cells in the plant body, retain their protoplasts (the cytoplasm, nucleus, and cell organelles) that carry out these functions.
Parenchyma cells are capable of cell division, even after they have differentiated into the mature form. They can therefore give rise to adventitious buds and roots at some distance from the apical meristem at the tips of shoots and roots. Parenchyma cells are also capable of further differentiation into new cell types under appropriate conditions, such as after trauma. (For information concerning the development of bark during the secondary growth of tissues, see below Vascular tissue (angiosperm).) Parenchyma cells are active in secretion, photosynthesis, and water and food storage (especially in fleshy fruits). They have large fluid-filled vacuoles (vacuole) that maintain cell turgidity; when a plant wilts, for example, it is because the vacuoles in the parenchyma cells have lost water and have become flaccid. Thus, parenchyma also functions in plant support. However, parenchyma cells do not have a secondary cell wall at maturity and thus remain flexible and capable of elongation.
Prosenchyma cells are starch-containing parenchymal cells whose cell walls have become lined with lignin, as occurs in the stems of Bougainvillea (Nyctaginaceae). A specialized type of parenchyma cell, called a transfer cell, is involved in the short-distance movement of solutes by cell-to-cell transfer. Transfer cells occur in association with veins in leaves and stems and also in many reproductive parts.
Collenchyma tissue (Figure 5-->) consists of collenchyma cells that also have retained their protoplasts. They are closely related to parenchyma, although they have thick deposits of cellulose in their primary cell walls, and the two types often intergrade in areas of continuity.Collenchyma is found chiefly in the cortex of stems and in leaves. For many herbaceous plants it is the chief supporting tissue, especially during early stages of development. In plants in which secondary growth occurs, the collenchyma tissue is only temporarily functional and becomes crushed as woody tissue develops. Collenchyma is located along the periphery of stems beneath the epidermal tissue. It may form a complete cylinder or occur as discrete strands that constitute the ridges and angles of stems and other supporting structures of the plant.
Collenchyma cells, polygonal in cross section, are much longer than parenchyma cells. The strength of the tissue results from the thickened cell walls and the longitudinal overlapping and interlocking of the cells. The wall is not uniformly thick in all cells, and thickening may occur predominately in longitudinal strips at the corners of the cell, on the tangential (i.e., outer, toward the stem exterior) surface of the cell, or around the spaces between adjacent cells. Pits are present in the cell wall and provide a mechanism for intercellular communication. An important feature of collenchyma is that it is extremely plastic—the cells can extend and thus adjust to increase in growth of the organ. Because collenchyma cells are alive at maturity, these thickenings may be reduced when meristematic activity is resumed as in formation of a cork cambium or in response to wounding.
sclerenchyma tissue (Figure 5-->) is composed of sclerenchyma cells, which are usually dead at maturity (i.e., have lost their protoplasts). They characteristically contain very thick, hard secondary walls lined with lignin; consequently, sclerenchyma provides additional support and strength to the plant body.The two principal types of sclerenchyma cells are sclereids and fibres. Sclereids vary in shape and size and may be branched. They are common in seed coats and nutshells. Apart from providing some internal support for various plant organs, sclereids deter desiccation of hard seeds, such as beans, and discourage herbivory of certain leaves.
Fibres are slender cells, many times longer than they are wide. They are highly lignified cells with tapering (oblique) end walls. The side walls of fibres are often so thick that the centre of the cell (the lumen) is often occluded. Fibres have great tensile strength and yet are also elastic. These qualities are significant in the flexible support of the stems of large herbs and leaves of many monocotyledons, such as palms. Leaf fibres are the source of Manila hemp (Musa textilis; Musaceae), sisal (Agave sisalana; Agavaceae), and many other fibre products. Fibres are found in various parts of the plant and are particularly common in the vascular tissues (see below).
Vascular tissue
Evolution of the transport process
Water and nutrients flow through conductive tissues (xylem and phloem) in plants just as the bloodstream distributes nutrients throughout the bodies of animals. This internal circulation, usually called transport, is present in all vascular plants, even the most primitive ones.
The importance of transport processes in plants increased as multicellular plants evolved and became larger and their tissues acquired specialized functions. As land plants developed, long-distance transport assumed an important role; not only are carbohydrates transported from the organs in which they are formed (the leaves) to other parts—such as reproductive organs (flowers and fruits), stems, and roots—but water and minerals must be transported to leaves, which are not submerged in water (as are those of most primitive nonvascular plants) but are in a relatively dry air environment. Highly developed land plants have two types of tissues specialized for long-distance transport: the xylem and the phloem. Water and dissolved mineral nutrients ascend in the xylem (the wood of a tree, such as an oak or a pine), and products of photosynthesis, mostly sugars, move from leaves to other plant parts in the phloem (the inner bark of a tree).
Evolving land plants faced not only the problem of transport but also the problem of supporting their weight. Aquatic plants are supported by their buoyancy in water and do not need a rigid stem; flotation devices such as gas-filled stomata (stomate) and intercellular spaces hold them upright and enable them to grow toward the water surface and obtain sufficient sunlight for photosynthesis. On land, a rigid, self-supporting structure is necessary for plants; this structure, the xylem, consists of tiny rigid tubes through which water and dissolved mineral nutrients can move. The rigidity of the tubes within a stem is sufficient to make it self-supporting.
Land plants take up water from the soil through the roots; some exceptions, such as some plants of the Peruvian desert that grow in dry soil and epiphytes that grow in tree canopies, rely on adaptations that enable them to obtain water from the air. In most plants, then, water ascends through the xylem, the tiny capillaries of the woody stem tissue, into all plant parts but primarily into the leaves, from which it is transpired (evaporated) into the air. In this way, the mineral nutrients are transferred from the soil to all above-ground plant parts.
Plants living in humid habitats, such as the small and primitive mosses and liverworts, do not have a well-developed xylem, but rather have similar cells called hydroids that lack true lignin. Similarly, water plants that have returned from land to an aquatic habitat during evolution have a reduced xylem; such plants, which have readapted to an aquatic environment, are not woody, because they need neither water-conducting tissues nor a self-supporting structure. On the other hand, tall land plants such as trees and vines (lianas (liana)) have the most highly developed long-distance transport systems. Vines differ from trees in that their xylem serves primarily for water conduction; they depend, for the most part, on other plants for support. Certain vines are of great length (a few hundred metres) and have extremely highly developed tissues for transporting water and nutrients.
Most of the material that composes a plant's dry weight is a consequence of photosynthesis, in which light energy is converted into chemical energy used to synthesize organic substances. Carbon dioxide from the air and water, which the plant takes from the soil, are utilized during photosynthesis, which occurs mostly in green plant parts—especially the leaves. Since plants shed their leaves either continuously or periodically but still increase in size, it is clear that many photosynthetic products must be transported out of the leaves and carried to all other plant parts; this process takes place primarily in the phloem.
The discovery of the functions of xylem and phloem (wood and bark) was made following that of the circulation of blood in the 17th century. By the early 19th century, it had been established that water ascends from roots into leaves through xylem and that photosynthetic products descend through phloem. Experiments now called girdling experiments were performed, in which a ring of bark is removed from a woody plant. Girdling, or ringing, does not immediately interfere with upward movement of water in the xylem, but it does interrupt phloem movement. In some plants surgical removal of phloem is difficult; in this case phloem may be killed by using steam (steam girdling). Xylem conduction is normally not affected by such treatment, and movement in the two transport tissues can thus easily be distinguished. Girdling experiments, however, are not entirely foolproof. The question as to whether or not mineral nutrients can ascend in the phloem illustrates the kinds of difficulties that may be encountered. Much smaller amounts of mineral nutrients reach the leaves in girdled plants than in ungirdled ones. From this observation it might be concluded that some nutrients ascend in the phloem of ungirdled trees; girdling, however, interrupts the flow of sugars into roots. Roots are thereby starved and take up fewer mineral nutrients; the reduced flow of mineral nutrients to the leaves of girdled plants can thus be explained as a secondary effect.
It was once widely believed that flogging a fruit tree increases the crop. The rational explanation of this is simple: flogging a tree damages the phloem—i.e., it girdles the tree, perhaps partially. Girdling increases the sugar concentration of the leaves by restricting downward transport to the roots and may thus directly or indirectly stimulate flowering or, if done after fruit-set, may increase fruit size by reallocating sugars destined for the roots to the developing fruits.
Structural basis of transport
Two features of plant cells differ conspicuously from those of animal cells. In plant cells the protoplast, or living material of the cell, contains one or more vacuoles (vacuole), which are vesicles containing aqueous cell sap. Plant cells are also surrounded by a relatively tough but elastic wall. Water entering the vacuole by osmosis (i.e., movement of water across a membrane from regions of higher water concentration into regions of lower water concentration that normally contain dissolved substances, such as cell interiors) expands the protoplast and consequently the cell wall until the internal pressure is balanced by the elastic counterpressure of the wall. Spaces between and within cell walls are sufficiently large to permit water to flow around all cells. The space available for free water flow is called apoplast. Water in apoplast originates from the roots and contains nutrients taken up by them. Nutrients enter a cell by crossing the outer cytoplasmic membrane (the plasmalemma or plasma membrane).
Most of the metabolic activities of the cell—the chemical reactions of living systems—occur within protoplasts. Substances can enter a protoplast by their cytoplasmic connections between neighbouring cells (plasmodesmata (plasmodesma)) or by active transport mechanisms requiring energy and a group of enzymelike compounds called permeases. Plasmodesmata may penetrate neighbouring cell walls at areas called primary pit fields. Also, some substances pass out of cells into the apoplast and are transported by energy-requiring processes into the protoplast of another cell.
Cell-to-cell transport takes place in all plants, but it is a slow process; the higher plants evolved the specialized tissues, xylem and phloem, for rapid long-distance transport. The woody tissue, xylem, contains highly specialized cells for water conduction. The cells are long and reinforced by strong, woody (lignified) walls; their protoplast breaks down and dissolves after wall growth is completed, so that the entire inside of the cell becomes available for rapid water conduction. In other words, the water-conducting cells of xylem are dead when functional. In the more primitive conifers the xylem consists largely of spindle-shaped cells called tracheids (tracheid), which have a diameter around 0.04 millimetre (0.0016 inch) and a length of about 3 millimetres (0.12 inch). Flowering plants have a more highly specialized xylem, in which the mechanical function and the water-conduction function have been separated during evolution. Tracheids, the primitive conducting cells, have evolved into fibres for mechanical strength and vessels (vessel) for water conduction, particularly in angiosperms. Vessel elements are barrellike cells with widths of up to 0.5 millimetre (0.02 inch) in some plants. Vessel elements are arranged end to end; their end walls are partly or wholly dissolved, and rows of such cells thus form long capillaries (tubes) up to several metres in length. These tubes are the vessels.
Numerous vessels of limited length thus provide a certain protection against injury—that is, since water pressures in the xylem are often well below zero (i.e., the water is under tension), air will be sucked into any injured xylem vessel and spread immediately throughout it but cannot pass through the wet pit membranes into the uninjured units. Damage is thus confined to the units that are injured and cannot easily spread. In addition, the smaller the conducting unit, the more confined is the damage. Plants with large, highly efficient vessels are much more vulnerable to injury, as is evident, for example, from the vulnerability of the elm, which has large vessels, to Dutch elm disease, in which the water-conduction vessels are injured by beetle activity and fungal growth. In general, both the less efficient but safer coniferous wood and the more highly efficient but more vulnerable wood of flowering plants have been successful during evolution. Very tall trees occur in both groups—e.g., Sequoia among the conifers and Eucalyptus among the flowering plants.
The conducting elements of the phloem underwent evolutionary changes somewhat similar to those of the xylem. The conducting elements of conifers, called sieve (sieve tube) cells, are similar in shape and dimensions to tracheids. They do not have a woody wall, however, and they are alive at functional maturity even though their cytoplasm may be highly specialized and the cells have usually lost their nucleus during development. In flowering plants the conducting elements in the phloem are called sieve elements and consist of sieve cells and sieve-tube members, the latter differing in having some sieve areas specialized into sieve plates (generally on the end walls). Sieve-tube members are arranged end to end to form sieve tubes, a name derived from the sievelike end walls through which passage of food from one cell to the next occurs. Sieve elements are almost invariably accompanied by special companion cells believed to control, to a certain extent, the metabolism of the nucleus-free conducting cells.
Organization of the vascular tissue
Vascular tissue is organized into discrete strands called vascular bundles, each containing xylem and phloem. In stems, the vascular tissue is organized into many discrete vascular bundles. In the roots, the vascular tissue is organized within a single central vascular cylinder. The anatomy of roots and stems is discussed in their respective sections below.
The xylem conducts water and minerals within the primary plant body, and the phloem conducts food. The xylem cells are arranged end to end to form a longitudinal continuum throughout the plant. The phloem cells form a similar continuum. Thus, water enters the xylem cells in the roots and travels to the leaves via the stems, and photosynthates (products of photosynthesis) enter the phloem cells in the leaves and are translocated to the roots via the stems. Storage parenchyma and fibres are generally present, and sclereids rarely are.
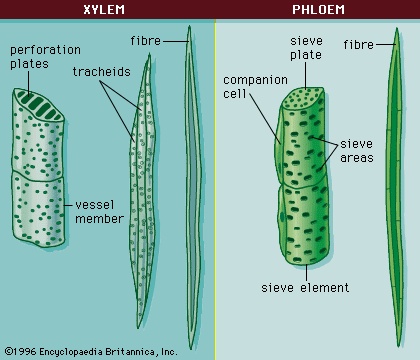 Primary xylem (Figure 6-->) consists of lignified tracheary elements (tracheids and vessel elements), which are dead at maturity (they have lost their protoplasts). Parenchyma cells also are interspersed throughout the tissue. Both tracheids and vessel elements are long hollow cells with tapered end walls. The end walls of adjacent tracheids contain paired small, rimmed, nonperforated pores, called bordered pits; water diffuses through a shared central membrane. The side walls have five patterns of thickening, which are believed to represent a developmental sequence from the initial xylem (protoxylem) to the final mature xylem (metaxylem): annular (a series of rings), helical (a long continuous spiral), reticular (a network), scalariform (a series of elongated bordered slits), and circular bordered pitting. Individual species may omit some of these patterns.
Primary xylem (Figure 6-->) consists of lignified tracheary elements (tracheids and vessel elements), which are dead at maturity (they have lost their protoplasts). Parenchyma cells also are interspersed throughout the tissue. Both tracheids and vessel elements are long hollow cells with tapered end walls. The end walls of adjacent tracheids contain paired small, rimmed, nonperforated pores, called bordered pits; water diffuses through a shared central membrane. The side walls have five patterns of thickening, which are believed to represent a developmental sequence from the initial xylem (protoxylem) to the final mature xylem (metaxylem): annular (a series of rings), helical (a long continuous spiral), reticular (a network), scalariform (a series of elongated bordered slits), and circular bordered pitting. Individual species may omit some of these patterns.Vessel elements differ from tracheids in that the end walls are modified into perforation plates, an area or areas in which there is no shared wall material or membrane. Vessel elements join to form continuous vessels. The perforations are much larger than those of the bordered pits of tracheids and are of four types: scalariform (slitlike), foraminate (circular), reticulate (a network), or simple (single). The bordered pitting of the side walls of vessel members is either scalariform or circular (generally scalariform bordered pitting is associated with scalariform, foraminate, or reticulate perforation plates). Vessel elements are found in the late metaxylem (the final, or most developed, form of the primary xylem).
The most common type of perforation plates in the angiosperms are scalariform and simple; the other types are rare. The putatively primitive angiosperms are without vessels and evolved from a condition in which only tracheids were present to one in which a series of long vessel elements had scalariform lateral walls and highly inclined end walls with many scalariform perforations, to short vessel elements with circular bordered pits in lateral walls and simple perforation plates in horizontal end walls.
This series of specializations has increased the efficiency with which water moves through the vessels: from the more generalized method of water diffusion through pit membranes of narrow tracheids to mass movement of water through the perforated end walls of relatively narrow scalariform vessels and then to relatively wide simple vessels with large single perforated end walls. This simple form is a rather streamlined system that facilitates the maximum movement of water in terms of amount and speed with the minimum amount of resistance, allowing for greater efficiency and effective water transport.
The primary phloem (Figure 6-->) is composed of sieve elements and fibres. Parenchyma cells are interspersed throughout. Sieve elements are longitudinal cells that transport food. They are composed of sieve cells and sieve-tube members. Sieve-tube members have clusters of pores in the cell walls known as sieve areas, which have either small pores or large pores; the latter are known as sieve plates. Sieve plates are mostly located on the overlapping adjacent end walls. As sieve-tube members differentiate, they lose their nucleus, ribosomes, vacuoles, and dictyosomes (the equivalent of the Golgi apparatus in animals); they are not dead, however, and remain metabolically active. Each sieve-tube member has an associated specialized parenchyma cell called a companion cell. They are derived by mitosis from the same parent cell and remain connected with each other. Photosynthates are actively secreted into, and actively removed from, sieve-tube members by their companion cells. Other unspecialized parenchyma cells also are present in primary phloem and provide storage.Finally, the primary vascular tissue system usually has fibres, particularly in herbaceous plants. The fibres occur in groups either around vascular bundles or as a cap over the phloem (phloem fibres).
 The primary vascular system (Figure 7-->) serves three functions. First, the sieve tubes conduct photosynthates via companion cells from green stems and leaves to nongreen areas (usually roots, lateral meristems, and shoot apical meristems) to promote growth and development. Second, tracheary elements provide a water-conducting system and a support system as a result of their rigid lignified cell walls. Third, fibres provide additional support.
The primary vascular system (Figure 7-->) serves three functions. First, the sieve tubes conduct photosynthates via companion cells from green stems and leaves to nongreen areas (usually roots, lateral meristems, and shoot apical meristems) to promote growth and development. Second, tracheary elements provide a water-conducting system and a support system as a result of their rigid lignified cell walls. Third, fibres provide additional support.Secondary vascular system
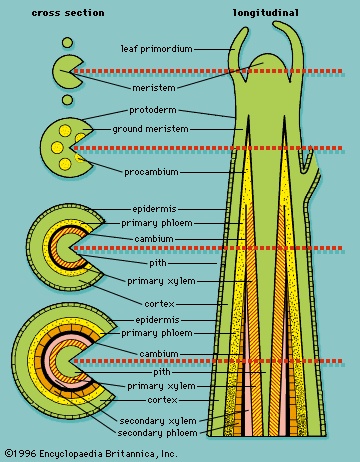 In woody plants, a vascular system of secondary vascular tissue develops from a lateral meristem called the vascular cambium (Figure 8-->). The vascular cambium, which produces xylem and phloem cells, originates from procambium that has not completely differentiated during the formation of primary xylem and primary phloem. The cambium is thought to be a single row of cells arranged as a cylinder that produces new cells: externally the secondary phloem and internally the secondary xylem. Because it is not possible to distinguish the cambium from its immediate cellular derivatives, which also divide and contribute to the formation of secondary tissues, the cambium and its immediate derivatives are usually referred to as the cambial zone.
In woody plants, a vascular system of secondary vascular tissue develops from a lateral meristem called the vascular cambium (Figure 8-->). The vascular cambium, which produces xylem and phloem cells, originates from procambium that has not completely differentiated during the formation of primary xylem and primary phloem. The cambium is thought to be a single row of cells arranged as a cylinder that produces new cells: externally the secondary phloem and internally the secondary xylem. Because it is not possible to distinguish the cambium from its immediate cellular derivatives, which also divide and contribute to the formation of secondary tissues, the cambium and its immediate derivatives are usually referred to as the cambial zone.Unlike the apical meristems, which consist of a population of similar cells, the cambium consists of two different cell types; the fusiform initials and the ray initials. The fusiform initials are elongated tapering cells that give rise to all cells of the vertical system of the secondary phloem and xylem (secondary tracheary elements, fibres, and sieve cells and the associated companion cells). The ray initials are isodiametric cells—about equal in all dimensions—and they produce the vascular rays, which constitute the horizontal system of secondary tissues; this horizontal system acts in the translocation and storage of food and water.
The fusiform and ray initials of the cambium divide in a plane tangential to the surface of the stem, with the long axes of the fusiform and ray initials parallel to the long axis of the plant organ. The cambium generates xylem mother cells toward the inside and phloem mother cells toward the outside. These cells in turn continue to divide tangentially, producing new cells that add to the xylem and to the phloem. Divisions of the cambium cells and xylem and phloem mother cells do not result in the production of equal amounts of secondary xylem and secondary phloem; because the cambium produces more cells internally than externally, more secondary xylem is produced than secondary phloem. Because divisions in the fusiform and ray initials are primarily tangential, new cells are regularly arranged in well-defined radial rows, a characteristic pattern for secondary vascular tissues.
Divisions in the cambium not only produce secondary vascular tissues but also increase the circumference of the cambium. As new cells are continuously added to the inside of the cambium, the cambium increases laterally (in circumference) to keep pace with the circumferential growth of the stem. In some plants, this is accomplished simply by radial division of the fusiform and ray initials. In other plants, the mechanism for increasing cambial diameter or increasing the number of cambial cells is more complex. If cambial activity is extensive, the primary tissues lying outside the cambium, such as primary phloem, cortex, and epidermis, are crushed by the pressure of new secondary tissue growth or become torn and obliterated because they cannot accommodate the rapidly increasing diameter of the plant.
As growth proceeds, the cork cambium forms in living cells of the epidermis, cortex, or, in some plants, phloem and produces a secondary protective tissue, the periderm. The cork cambium is, like the vascular cambium, a lateral meristem that produces cells internally and externally by tangential divisions. Unlike the cambium, the cork cambium consists of one cell type.
Another type of meristem active in certain plants, especially grasses, is the intercalary meristem. These cells possess the ability to divide and produce new cells, as do apical and lateral meristems. They differ, however, in being situated between regions of mature tissue, such as at the base of grass leaves, which are themselves located on mature stem tissue. In many instances intercalary meristems function for only a short time and eventually completely differentiate into mature tissues. Intercalary meristems are usually located at positions on the stem where leaves have emerged (nodes) and are largely responsible for elongation in grass shoots and leaves. Intercalary meristems are the internode regions where cell division of the ground meristem persists for a longer time than in other areas of the internode. In rosette plants, intercalary meristems are lacking.
Secondary xylem is composed of tracheary elements, rays, fibres, and interspersed axial parenchyma cells. The tracheary elements consist of only tracheids, as in the few vessel-less angiosperms (e.g., Winteraceae), or of both tracheids and vessel elements, as in the vast majority of angiosperms. Axial parenchyma may surround the vessel elements (paratracheal) or be randomly dispersed among the vessel elements (apotracheal).
Tyloses are balloonlike outgrowths of parenchyma cells that bulge through the circular bordered pits of vessel members and block water movement. The presence of tyloses in white oaks (white oak) makes their wood watertight, which is why it is preferred in casks and shipbuilding to red oak, which lacks tyloses and does not hold water. In trunks and branches that lean, there is eccentric growth of tension wood on the upper surface; tension wood is a type of reaction wood found in angiosperms that contains gelatinous fibres which shrink and pull.
Growth rings (growth ring) in the secondary xylem of temperate woody angiosperms are usually annual, but under environmental fluctuations, such as drought, more than one can form, or none at all. Growth rings result from the difference in density between the early wood (spring wood) and the late wood (summer wood); early wood is less dense because the cells are larger and their walls are thinner. Although the transition of early wood to late wood within a growth ring may be obscure, that demarcation between the adjacent late wood of one ring and the early wood of the next ring is clear. Diffuse-porous wood occurs when the size of the vessels (pores) in a growth ring are fairly uniform and evenly distributed (e.g., red maple, Acer rubrum; Sapindaceae). Ring-porous wood occurs when the pores of the early wood are distinctly larger than those of the late wood (e.g., black walnut, Juglans nigra; Juglandaceae).
Both xylem and phloem have limited longevity. The oldest phloem layers are the outermost—the dead bark of the stem surface. The yearly amounts of xylem visible as distinct rings in cross sections of stems are known as annual rings. The oldest xylem layers (i.e., the oldest annual rings) are in the dead central core, or heartwood, of the woody stem, which can often be recognized by its darker coloration. The lighter-coloured sapwood is living and functions as storage tissue and, especially in the outermost sapwood, as conducting tissue; the younger annual rings make up the sapwood. In some highly specialized tree species with large vessels (such as some oaks, ashes, and others), only the very outermost growth ring functions in water conduction.
Conducting tissues seldom run straight along a tree stem; usually they are arranged in a helical or spiral pattern, sometimes called the spiral grain of a tree. The angle of the spiral arrangement usually changes from year to year; the path of water up a tree stem may therefore be very complicated if more than one growth layer acts as a conducting tissue. Functionally, the effect of the variable spiral grain is to distribute water to all parts of the tree from any root.
The secondary phloem of angiosperms consists of sieve-tube members, companion cells, scattered parenchyma, ray parenchyma, and fibres. The fibres usually occur in clusters or as bands alternating with bands of sieve tubes and parenchyma cells. As the vascular cambium continues to produce more secondary xylem to the inside, the older (most exterior) portions of the secondary phloem are crushed, die, and are sloughed off as part of the bark. Successive cork cambiums (see below Dermal tissue (angiosperm)), essentially lateral meristems from which the bark arises, originate in the parenchyma of the phloem and produce additional cork.
Uptake of water and mineral nutrients from the soil
Water uptake from the soil by root cells is passive, in that water may be pulled into the root by low xylem pressure and also follows osmotic gradients caused by the mineral nutrients, which are taken up actively (i.e., with the expenditure of metabolic energy) across root cell membranes. As the mineral nutrients—the ions (charged components) of inorganic salts—are taken up, they are largely incorporated into organic molecules. Thus, the solutes in xylem sap are mostly complex organic substances, sometimes of a specific nature; for example, nicotine synthesis takes place in the roots of tobacco plants, where nitrogen is incorporated into compounds that have moved to the roots through the phloem as sugars. If a tomato shoot is grafted onto a tobacco rootstock, nicotine-containing tomato leaves are formed. On the other hand, a tobacco shoot grafted onto a tomato rootstock results in a plant with nicotine-free tobacco leaves. Many other specific nitrogen-containing substances originate in the roots; in most plants, however, nitrogen is transported to the leaves from the roots in the form of compounds known as amino acids (amino acid) and amides (amide).
The major chemical elements (chemical element) needed by a plant are carbon, hydrogen, oxygen, phosphorus, potassium, nitrogen, calcium, iron, and magnesium; in addition, many other elements are required in very small amounts. A lack of any element may result in deficiency diseases. A few elements taken up by plants are of no nutritive value and usually are eliminated or crystallized (e.g., silica), sometimes by deposition in special cells.
The plant is able to control to some extent the substances that enter. If equal amounts of sodium and potassium are available to roots of plants, and the amount of the two elements inside the plant is analyzed, less sodium is likely to be found than potassium. The structural basis for the control of uptake of substances into roots is the so-called Casparian strip, a conspicuously thickened wall area one cell layer deep surrounding primary roots; it prevents excess soil solution from being pulled directly into the central part of the root where the xylem is located. As a result, the soil solution has to pass through a cell barrier in which uptake can be metabolically controlled. After nutrients are inside living root cells and have been converted to appropriate compounds, the latter are released into the xylem and move to above-ground parts.
Process of xylem transport
The total amount of conducting tissue remains about the same from roots to leaves. In terms of water movement, the velocity of movement might be expected to be uniform throughout the entire axial system of stem, branches, and twigs. Because some trees (e.g., oaks (oak)) have thick twigs, however, the velocity of water movement is greater in the stem than in the twigs at any time. Similarly, in tree species with slender branches (such as birches (birch)), the reverse is true. Normally the proportion of xylem to leaves supplied by that xylem is greater in plants growing in dry habitats than in plants found in wet ones and may be as much as 700 times greater in certain desert plants than in aquatic plants and herbs of relatively humid forest floors. The leaves of dry-habitat plants thus are more richly supplied with water-conducting xylem tissue than are those of moist habitats.
The velocity of sap movement in trees varies throughout a 24-hour period. During the night, especially a rainy night, sap flow may stop; velocity increases with daylight, peak rates being found in the early afternoon. Peak velocities correlate with vessel size; the rate of sap flow in trees with small vessels is about 2 metres (7 feet) per hour; that in trees with large vessels, about 50 metres (160 feet) per hour. The energy required to lift water in both cases is comparable; in trees with large pores, water simply moves faster through fewer and larger vessels.
It was demonstrated about 1900 that living cells of the stem are not responsible for water movement. It is now generally recognized that water in the xylem moves passively along a gradient of decreasing pressures. Under certain special conditions, water is pushed up the stem by root pressure. This may be the case with herbaceous (nonwoody) plants in the greenhouse under conditions of ample water supply and little transpiration. In nature, these conditions may be met in early spring before the leaves emerge, when the soil is wet and transpiration is low. Under such conditions, water movement is caused by active uptake of ions (charged particles) and by the entry of water from the soil into the roots. Most of the time, however, water is pulled into the leaves by transpiration. A gradient of decreasing pressures from the base to the top of a tree can be measured, even though pressures are low.
A vacuum pump cannot pull water to a height of more than 10 metres (about 33 feet). Since many trees are far taller than 10 metres, the mechanism by which they move water to their crowns has been investigated. Is it possible for trees to pull water into their crowns along a decreasing pressure gradient or do they employ some other mechanism? If trees pull water, that in the xylem would have to be held on the tracheid and vessel walls by adhesion, and water molecules would have to hold together by cohesion. The hypothesis that water is pulled upward along a pressure gradient during transpiration has been called the cohesion theory (cohesion hypothesis). Two critical requirements of the cohesion mechanism of water ascent are (1) sufficient cohesive strength of water and (2) existence of tensions (i.e., pressures below zero) and tension gradients in stems of transpiring trees.
Although the tensile strength of water is very high, an excessive pull exerted on a water column will break it. The tallest trees are about 100 metres (330 feet) high. A nonmoving water column at an atmospheric pressure of 1 atmosphere at the base of the tree is exposed to a pressure of −9 atmospheres (i.e., a tension of 9 atmospheres) at the top. Under conditions of peak flow at midday, this gradient increases by about 50 percent; in other words, a transpiring sequoia would have a pressure in the xylem of at least −14 atmospheres at the top if the basal pressure is 1 atmosphere. If the pressure at the base is −10 atmospheres because of dry soil, however, the pressure at the top drops to −25 atmospheres. It has been demonstrated that water columns in the xylem can withstand this tension, or pull, without breaking.
Negative pressures and gradients of negative pressures have been shown to exist in trees with an ingeniously simple device called the pressure bomb. A small twig is inserted in a container (the pressure bomb), its cut stump emerging from a tightly sealed hole. As pressure is applied to the container and gradually increased, water from the xylem emerges from the cut end as soon as the pressure being applied is equal to the xylem tension that existed when the twig was cut. This method has been used to measure gradients of negative pressures in trees. Movement in the xylem is by mass flow of the whole solution, and the force is either the tension pull of transpiration or root pressure, or both. In general, however, water movement in the xylem is by transpiration pull. The mechanism of phloem transport remains unclear (see below).
Process of phloem transport
Products of photosynthesis (primarily sugars) move through phloem from leaves to growing tissues and storage organs. The areas of growth may be newly formed leaves above the photosynthesizing leaves, growing fruits, or pollinated flowers. Storage organs are found in roots, bulbs, tubers, and stems. Thus the movement in the phloem is variable and under metabolic control (whereas movement in xylem is always upward from the roots).
The rate at which these substances are transported in the phloem can be measured in various ways—e.g., as velocities in distance traveled per unit time or as mass transfer in (dry) weight transported per unit time. Velocities appear to be graded—i.e., some molecules move faster than others within the same channel. Peak velocities of molecules usually are of the order of 100 to 300 centimetres (40 to 120 inches) per hour. Average velocities, more difficult to measure but significant in mass-transfer considerations, are lower.
Mass transfer can be measured by weighing a storage organ, such as a potato tuber or a fruit, at given time intervals during its growth. Mass transfer per cross-sectional area of conducting tissue is referred to as specific mass transfer and is expressed as grams per hour per square centimetre of phloem or sieve tubes. With a given specific mass transfer, the velocity with which a liquid of a certain concentration flows can be calculated; in dicotyledonous stems, for example, specific mass transfer is between 10 and 25 grams per hour per square centimetre of sieve tube tissue at times of peak performance. In certain tree species the sieve tubes can be tapped to obtain an exudate. The concentration of this exudate, multiplied by the measured average velocity, is of the same order of magnitude as specific mass transfer, indicating that liquid movement through sieve tubes could account for transport.
Much of the experimental work on phloem transport now is done with the aid of radioactive substances; for example, when radioactive carbon dioxide administered to an illuminated leaf is incorporated into sugar during photosynthesis and carried from the leaf, the velocity of this movement can be measured by determining the arrival of radioactivity at given points along the stem. Whole plants, as long as they are reasonably small, can be pressed against photographic film after the conclusion of a similar experiment, and the photographic image will indicate the areas to which radioactive sugar has moved.
The mechanism of phloem transport has been studied for many years. A number of hypotheses have been put forth over the past years, but none is entirely satisfactory. One fundamental question is whether sugars and other solutes move en masse as a flowing solution or whether the solvents diffuse independently of the solvent water. The phenomenon of exudation from injured sieve tubes (sieve tube) supports the first possibility, which has been further supported by a discovery involving aphids (aphid) (phloem-feeding insects): when aphids are removed from plants while feeding, their mouthparts remain embedded in the phloem. Exudate continues to flow through the mouthparts; the magnitude of the rate of this exudation indicates that transport within the sieve tube to the mouthparts occurs as a flow of solution.
Evidence against solution flow is the movement of substances in opposite directions through a section of phloem at any one given time. This, however, has never been convincingly demonstrated in just one sieve tube. On the other hand, attempts to find simultaneous movement of sugars and water along a phloem path, in order to demonstrate solution flow, have been only partially successful.
Mass-flow hypotheses include the pressure-flow hypothesis, which states that flow into sieve tubes at source regions (places of photosynthesis or mobilization and exportation of storage products) raises the osmotic pressure in the sieve tube; removal of sugars from sieve tubes in sink regions—i.e., those in which sugars are removed or imported for growth and storage—lowers it. Thus a pressure gradient from the area of photosynthesis (source) to the region of growth or storage (sink) is established in sieve tubes that would allow solution flow. The electroosmotic hypothesis postulates that solution is moved across all sieve plates (areas at which individual sieve elements end) by an electric potential that is maintained by a circulation of cations (positively charged chemical ions), such as potassium. Transport hypotheses postulating solute movement independent of solvent water include the spreading of solute molecules between two liquid phases and the active transport of molecules by a type of cytoplasmic movement that is often referred to as cytoplasmic streaming.
During the life of a leaf, its role as a sink or a source changes. A young developing leaf before it is photosynthetic is a sink for sugars produced by older leaves. After the leaf begins to expand and turn green, it is both a sink (importer of sugar) and a source (exporter of sugar) as a result of its own photosynthetic capacity. When mature and fully expanded, the leaf then becomes a source of sugar production.
Transport and plant growth
It is important to realize that the plant, with its two transport systems, xylem and phloem, is able to move any substance to virtually any part of its body; the direction of transport is usually opposite in the two systems, and transfer from one system to the other takes place easily. An exception is transport into flowers and certain fruits, in which flow in each system is unidirectional.
Numerous substances move from roots to mature leaves through xylem and are transferred from the leaves, together with sugars, through the phloem to other plant parts. In the autumn months in temperate regions, plants store most of the products resulting from photosynthesis during the summer months in structures such as stems, bulbs, and tubers and mobilize it in the spring when new growth begins. A few plants, such as some tropical monocotyledons (certain palms (palm), for example), store food for many years for use at the time of flowering and fruit-set at the end of their lives.
Plant hormones (hormone), or growth regulators, are effective in very small amounts; they induce or enhance specific growth phenomena. Because the site of hormone synthesis is different from its place of action, hormones must be transported before they can exert their effects. There are five major types of plant hormones, including auxin, gibberellin, cytokinin, ethylene, and abscisic acid. Each type plays a different role in plant growth and development, from influencing cell division, fruit ripening, and seed dormancy to directing stem elongation and food mobilization. The best-characterized of these hormones are the auxins (auxin), the most common of which is called indoleacetic acid. Auxins are formed in young, growing organs, such as opening buds, and are transported away from tips of shoots toward the base of the plant, where they stimulate the cells to elongate and sometimes to divide. Responses to gravity and light are also under auxin control. Auxins move to the lower side of a leaning stem; cells on the lower side then elongate and cause the stem to bend back to a vertical position. Response to gravity in many roots is the opposite of that in shoots; the same mechanism of auxin distribution is responsible, but roots react to different quantities of the hormone than do shoots. Similar auxin distributions are responsible for phototropic responses—i.e., the growth of plant parts such as shoot tips and leaves toward light. In certain cases auxin may be destroyed on the illuminated side, and the unilluminated side with more auxin elongates, causing the shoot to bend toward the light.
Auxins are not normally transported through vascular tissue; moreover, transport is polar—i.e., it takes place along the stem from tip to base, regardless of the stem's position. Velocities of transport are of the order of 5 to 10 millimetres (0.2 to 0.4 inch) per hour, and transport requires the expenditure of metabolic energy. There is evidence that most growth hormones can be transported through xylem or phloem, but, at least in the case of auxin, the transport mechanism is specific directionally from morphological top to bottom.
Hormone transport is also involved in the stimulation of flowering (flower). In some plants, flowering is triggered by short or long days. The receptor of this stimulus is in the leaves. A chemical substance, probably a flowering hormone of an as-yet-unknown nature, then moves to the shoot apex and causes a transformation of the vegetative growing point into a flowering shoot.
Many growth-correlating phenomena are effected by transported hormonal stimuli. A vigorously growing terminal (topmost) shoot may inhibit lateral buds lower down from growing out and may force later branches to bend down. If the terminal shoot is removed, laterals grow out and topmost lateral branches bend upward. In leaning trees with secondary tissue (wood), the cambium produces compression wood on the lower side (in conifers (conifer)) or tension wood on the upper side (in dicotyledons (dicotyledon)) in response to a hormone; the stem responds by pushing (in conifers) or pulling (in dicotyledons) itself upright. Transport of growth-regulating substances is thus largely responsible for the characteristic shape of each plant species.
Dermal tissue
The dermal tissue system—the epidermis—is the outer protective layer of the primary plant body (the roots, stems, leaves, flowers, fruits, and seeds). The epidermis is usually one cell layer thick, and its cells lack chloroplasts.
As an adaptation to a terrestrial habitat, the epidermis has evolved certain features that regulate the loss of water, carbon dioxide, and oxygen. Cutin and waxes are fatty substances deposited in the walls of epidermal cells, forming a waterproof outer layer called the cuticle. Often, epicuticular waxes, in the form of sheets, rods, or filaments, are exuded over the cuticle, giving some leaves their whitish, greenish, or bluish “bloom.” The cuticle and epicuticular waxes minimize transpiration from the plant. The waxy deposits can be thin or thick, depending on the requirements of the plant; for example, desert plants usually have heavy wax coatings.
The plant, however, must have some means of exchanging water vapour, carbon dioxide, and oxygen through this cuticle barrier. Dispersed throughout the epidermis are paired, chloroplast-containing guard cells, and between each pair is formed a small opening, or pore, called a stoma (stomate) (plural: stomata). When the two guard cells are turgid (swollen with water), the stoma is open, and, when the two guard cells are flaccid, it is closed. This controls the movement of gases, including water vapour in transpiration, into the atmosphere. Guard cells and stomata are found on aerial plant parts, most frequently on leaves, but are not known to occur on aerial roots.
The trichomes (pubescences) that often cover the plant body are the result of divisions of epidermal cells. Trichomes may be either unicellular or multicellular and are either glandular, consisting of a stalk terminating in a glandular head, or nonglandular, consisting of elongated tapering structures. Leaf and stem trichomes increase the reflection of solar radiation, thereby reducing internal temperatures, and thus reduce water loss in plants growing under arid conditions.
Epiphytic bromeliads (air plants (epiphyte) such as Spanish moss, Tillandsia usneoides; Bromeliaceae) absorb water and minerals via foliar trichomes. The glandular trichomes produce and secrete substances such as oils, mucilages, resins, and, in the case of carnivorous plants, digestive juices. Plants growing in soils with high salt content produce salt-secreting trichomes (e.g., saltbush, Atriplex vesicaria; Amaranthaceae) that prevent a toxic internal accumulation of salt. In other cases, trichomes help prevent predation by insects, and many plants produce secretory (glandular) or stinging hairs (e.g., stinging nettle, Urtica dioica; Urticaceae) for chemical defense against herbivores. In insectivorous plants, trichomes have a part in trapping and digesting insects. Prickles, such as those found in roses, are an outgrowth of the epidermis and are an effective deterrent against herbivores.
As defined above, the epidermis is the outermost protective layer of the primary plant body. At a certain stage in their life cycle, woody plants cease to grow in length and begin to add to their girth, or width. This is accomplished not by the addition of more primary tissue but by the growth of secondary vascular tissue around the entire circumference of the primary plant body. The secondary vascular tissue arises from the vascular cambium, a layer of meristematic tissue insinuated between the primary xylem and primary phloem (see above Vascular tissue (angiosperm)). Secondary xylem develops on the inner side of the vascular cambium, and secondary phloem develops on the outermost side. A second lateral cambium, called the phellogen or cork cambium, is the source of the periderm, a protective tissue that replaces the epidermis when the secondary growth displaces, and ultimately destroys, the epidermis of the primary plant body.
In woody plants, the phellogen, or cork cambium, arises in any of the three tissue systems near the surface of the plant body. The cork cambium produces cork cells toward the outside and parenchyma cells toward the inside. As a unit, the cork cambium, cork cells, and parenchyma (phelloderm) form the periderm. Like the epidermis, the periderm is a protective tissue on the periphery of the plant body; however, because the periderm is produced by a lateral meristem, it is considered to be of secondary origin (in contrast to the primary origin of the epidermis from the protoderm). At maturity the cork cells are nonliving, and their inner walls are lined with suberin, a fatty substance that is highly impermeable to gases and water (which is why cork is used to stop wine bottles). The walls of cork cells may also contain lignin.
In stems, the first cork cambium usually arises immediately inside the epidermis or in the epidermis itself. In roots, the first cork cambium appears in the outermost layer of the vascular tissue system, called the pericycle (see below Plant organs: Roots).
The meristematic tissue of the cork cambium produces more and more derivatives of cork cells and parenchyma and displaces them into the outer margins of the plant body. Because the epidermal cells do not divide, they cannot accommodate an increase in stem diameter. Thus, the epidermal cells soon become crushed by the growing number of cork cells derived from the cork cambium, eventually die, and are sloughed off.
The epidermis is then replaced by cork cells until eventually the original cork cambium ceases to produce derivative cork and is itself destroyed. A new cork cambium eventually arises in the secondary phloem situated just behind the old cork cambium. That portion of the secondary phloem that forms between the new cork cambium and the old one becomes crushed and displaced externally as well. This process is repeated often each growing season.
The term cork is used to denote the outer derivatives of the cork cambium specifically. bark, on the other hand, is an inclusive term for all tissues outside of the vascular cambium. The two regions of the bark are the outer bark, composed of dead tissues, and the inner bark, composed of living tissues of the secondary phloem. Outer bark is shed continually from a tree, often in a distinctive pattern, as the circumference increases because its dead cells cannot accommodate the increased diameter. Bark contributes to the support of the tree and protects the living tissue of the active secondary phloem and vascular cambium from desiccation and from such environmental disturbances as fire.
Plant organs
Roots (root)
The root apical meristem, or root apex, is a small region at the tip of a root in which all cells are capable of repeated division and from which all primary root tissues are derived. The root apex is protected as it passes through the soil by an outer region of living parenchyma cells called the root cap. As the cells of the root cap are destroyed and sloughed off, new parenchyma cells are added by a special internal layer of meristematic cells called the calyptrogen. Root hairs also begin to develop as simple extensions of protodermal cells near the root apex. They greatly increase the surface area of the root and facilitate the absorption of water and minerals from the soil.
Along the longitudinal axis of a root, beginning with the root cap and leading away from the root tip, there are five distinct zones in which certain specific growth patterns dominate: cell division, cell elongation, primary tissue maturation, mature primary tissues, and secondary tissue growth (the latter is found in woody roots—i.e., those of perennial dicotyledons). There is a gradual transition between these regions.
The region of cell division includes the apical meristem and the primary meristems—the protoderm, ground meristem, and procambium—derived from the apical meristem. As is generally true of nonmeristematic regions elsewhere in the plant body, root length in the second region is increased as a result of cell elongation rather than by cell division. The region of maturation that follows is where the cells differentiate (i.e., change in structure and physiology into cells of a specific type) and where the first primary phloem and xylem, as well as mature root hairs, are clearly seen. The region of mature primary tissues is where the anatomy of the primary body of the root is most obvious and where all the elements of the vascular cylinder, cortex, and epidermis are evident. Finally, in the region of secondary growth, the secondary xylem and phloem as well as the periderm add girth to the plant.
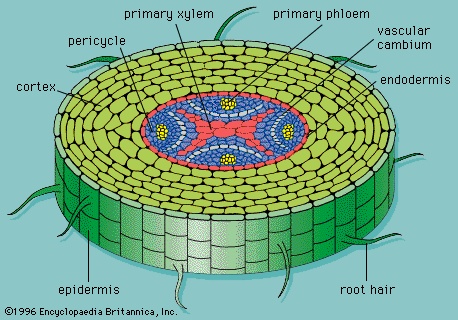 There are many individual vascular strands (or vascular bundles) in the primary body of the stem (see below Stems), and they all converge into a single central vascular cylinder in the root, forming a continuous system of vascular tissue from the root tips to the leaves. At the centre of the vascular cylinder of most roots is a solid, fluted (or ridged) core of primary xylem (Figure 9-->). The primary phloem lies between these flutes or ridges. Parenchyma cells are dispersed throughout the vascular cylinder.
There are many individual vascular strands (or vascular bundles) in the primary body of the stem (see below Stems), and they all converge into a single central vascular cylinder in the root, forming a continuous system of vascular tissue from the root tips to the leaves. At the centre of the vascular cylinder of most roots is a solid, fluted (or ridged) core of primary xylem (Figure 9-->). The primary phloem lies between these flutes or ridges. Parenchyma cells are dispersed throughout the vascular cylinder.The vascular cylinder of the root is surrounded by a layer of parenchymatous pericycle cells. As the root ages, many of these cells become fibres, particularly in monocotyledons and many herbaceous dicotyledons. As defined above (see Tissue systems: Ground tissue (angiosperm)), a characteristic feature of parenchyma cells is their ability to differentiate into cells of a different type under appropriate conditions. The parenchyma cells of the pericycle, then, can be considered meristematic in that they give rise to new lateral meristems and lateral roots. In woody roots the vascular cambium (the lateral meristem that gives rise to secondary phloem and secondary xylem) originates in the pericycle as well as in the procambium; the procambium is the primary meristematic tissue between the primary phloem and xylem. The first cork cambium is a lateral meristem that arises in the pericycle; the successive cork cambia arise in the secondary phloem. Because lateral roots are initiated in the pericycle and grow out through the cortex and epidermis, they are said to have an internal, or endogenous, origin, in contrast to the external, or exogenous, origin of leaves and the apical meristem of stems (see below Stems (angiosperm)).
Ground tissue called the cortex surrounds the vascular cylinder and pericycle. The cortex of roots generally consists of parenchyma cells with large intercellular air spaces. The endodermis (the innermost layer of the cortex adjacent to the pericycle) is composed of closely packed cells that have within their walls Casparian strips, water-impermeable deposits of suberin that regulate water and mineral uptake by the roots. The cortex is surrounded by the dermal system consisting of a single layer of epidermal cells.
The few variations that occur in root anatomy are mainly found among the monocotyledons. The roots of monocotyledons lack secondary growth. Monocotyledons also generally have a parenchymatous pith in the centre of the vascular cylinder and fibres or sclereids, or both, in the cortex; and extensive well-developed pericyclic fibres. Orchids (orchid) have a multiple-layered epidermis called a velamen, which consists of nonliving compact cells with lignified strips of secondary walls. These cells provide support, prevent water loss, and assist the plant in absorbing water. When dry the orchid root appears white, and when wet the root appears green because the cells of the velamen absorb water, become translucent, and reveal the green cortical cells.
Stems (stem)
The shoot apical meristem and the primary meristems lie at the apex of the shoot and give rise to the primary tissues of the stem. The shoot apical meristem produces leaves and axillary buds exogenously; as a result, the epidermis of stems and leaves is continuous. (In contrast, as mentioned above, the lateral roots are produced endogenously, and the dermal system of the lateral roots is discontinuous with that of the parent root.)
The stem has growth periods similar to those of the root, but longitudinal regions are not as obvious as in the root until the nodes become differentiated and internode lengths increase. Elongation of internodes involves many cell divisions and is followed by cell elongation. At this point, growth in thickness involves some radial cell division and cell enlargement.
The primary tissue systems appear after internode elongation. The procambium differentiates as a basically continuous hollow cylinder or discrete procambial strands, which differentiate into primary xylem and phloem. The ground tissue that lies outside the procambial cylinder is the cortex, and that within is the pith. Ground tissue called the interfascicular parenchyma lies between the procambial strands and remains continuous with the cortex and pith. As the vascular tissue grows, xylem and phloem develop, the vascular bundles mature, the single-layered epidermis differentiates as epidermal cells, trichomes, and a few stomata, and the parenchymatous pith may develop as collenchyma or contain sclereids or fibres or both; unequal pith proliferation and expansion produces the flattened stems (pads) of prickly-pear (prickly pear) cacti (Opuntia; Cactaceae). The parenchymatous cortex also may develop some collenchyma, sclereids, or fibres; unequal growth and expansion of the cortex produces the cladodes of epiphytic cacti (e.g., night-blooming cereus, Selenicereus; Cactaceae). In most aquatic angiosperms, the parenchymatous cortex contains large intercellular spaces. As a rule, angiosperm stems have no endodermis or definable pericycle.
The most common arrangement of the primary xylem and phloem is called a collateral bundle; the outer portion of the procambium (adjacent to the cortex) becomes phloem, and the inner portion (adjacent to the pith) becomes xylem. In a bicollateral bundle, the phloem is both outside and inside the xylem, as in Solanaceae (the potato family) and Cucurbitaceae (the cucumber family). In the monocots, the phloem may surround the xylem, or the xylem may surround the phloem.
The vascular bundles of the stem are continuous not only with the primary vascular system of the root but also with the vascular bundles of the leaves. At each node, one or more longitudinal stem bundles enter the base of the leaf as leaf traces, connecting the vascular system of the stem with that of the leaf. The point at which the stem bundle diverges from the vascular cylinder toward the leaf is a leaf gap, called a lacuna. The number of lacunae varies among angiosperm groups and remains a characteristic for classifying the various species.
Several leaves in a line along the stem have common stem bundles. In some species all stem bundles and their associated leaf traces are interconnected, while in others each stem bundle and the associated leaf trace remains laterally independent of the others. An arrangement of two trace leaves and a single lacuna is found among several primitive angiosperm families and throughout the gymnosperms and is the organization from which other nodal patterns are derived.
In woody dicots (dicotyledon), the vascular cambium is formed in parts that grow toward each other and unite. Each vascular bundle develops a meristematic area of growth from an undifferentiated (parenchymatous) layer of cells between the primary xylem and primary phloem, called a fascicular cambium. This meristematic area spreads laterally from each bundle and eventually becomes continuous, forming a complete vascular cambium.
The arborescent (treelike) stems of monocotyledons have a different growth pattern and anatomy from dicotyledons. Scattered throughout the ground tissue are vascular bundles with no fascicular cambia and no definable pith or cortex. Secondary growth, when it occurs, is different because a secondary thickening meristem forms under the epidermis. This secondary thickening meristem produces secondary parenchyma (conjunctive tissue) to the inside, and then secondary vascular bundles develop within this conjunctive tissue. Thus, there are no rings of secondary xylem or secondary phloem as in woody dicotyledons.
Many arborescent monocots have only massive primary growth without secondary growth. This primary growth is derived from a primary-thickening meristem under the leaf bases that is a lateral continuation of the apical meristem. This primary-thickening meristem produces vast amounts of parenchyma to the inside, through which the leaf traces differentiate.
Leaves
Leaves initially arise from cell divisions in the shoot apical meristem. A slight bulge (a leaf buttress) is produced, which in dicots continues to grow and elongate to form a leaf primordium. (Stipules, if present, appear as two small protuberances.) Marginal and submarginal meristems on opposite flanks of the primordium initiate leaf-blade formation. Differences in the local activity of marginal meristems cause the lobed shapes of simple leaves and the leaflets in compound leaves. An increase in width and in the number of cell layers is brought about by marginal meristems. Subsequent expansion and increase in length is achieved by cell division and the general enlargement of cells throughout the blade.
Leaf growth is determinate; the tip matures first, and maturation then progresses toward the base, after which the leaf cells cease to grow and divide. (Stem growth is generally indeterminate since the meristems are active indefinitely.) The petiole, when present, and the leaf base become thickened, and often the latter expands laterally and fully or partially encloses the stem. Soon after the cells of the marginal meristems begin to divide, procambial strands differentiate into the leaf from the stem bundles to form the midvein, or midrib. The smaller lateral veins of the leaf are initiated near the leaf tip; subsequent major lateral veins are initiated sequentially toward the base, following the overall pattern of leaf development. A major lateral vein may have one or more orders of smaller veins, which also are initiated in size from larger to smaller. This results in the netlike venation patterns characteristic of dicotyledonous leaves.
The anatomy of a mature dicot leaf generally reflects the habitat, especially the availability of water. Mesomorphic leaves are adapted to conditions of abundant water and relatively humid conditions; xeromorphic (xerophyte) leaves are adapted to dry conditions with relatively low humidity; and hydromorphic leaves are adapted to aquatic situations, either submerged or in standing water. Mesomorphic leaves (the most common type) are characteristic of crop plants, such as tomatoes and soybeans. Their veins (vascular bundles) permeate the ground tissue of the dermal system—a single layer of epidermal cells with interspersed guard cells. The ground tissue system, the mesophyll, is divided into two regions: the palisade parenchyma, located beneath the upper epidermis and composed of columnar cells oriented perpendicular to the leaf surface, and spongy parenchyma, located in the lower part of the leaf and composed of irregularly shaped cells. The veins contain primary xylem and phloem and are enclosed by a layer of parenchyma called the bundle sheath. Only the midvein and some large lateral veins have any secondary growth.
The anatomy of mesomorphic leaves is designed to function optimally for water uptake and gas exchange in photosynthesis under mesic (moist) conditions. The spongy mesophyll with irregularly shaped cells provides increased surface area internally, while the elongate palisade cells provide optimal exposure of chloroplasts to light.
The anatomy of hydromorphic leaves is simplified: the cuticle is thin or lost; the guard cells are raised and are found only on the upper surface in floating leaves (they are lost in most submerged leaves); the mesophyll contains aerenchyma (an adaptation to promote water loss) and little or no collenchyma or sclerenchyma; and the vascular system (particularly the water-conducting element of the vascular system, the xylem) is only weakly developed since the water provides much of the mechanical support to the plant normally provided by the xylem. The abundance of water means that there is no need for mechanisms that prevent water loss and little need for additional supports. The leaves generally become large and thin, and the reduction or loss of cuticle, vascular tissue, and ground tissue (mesophyll) permits the rapid loss of water vapour (transpiration). The guard cells on the upper surface of floating leaves also monitor the rate of water loss through the central stomata. Such plants may wilt if the turgor (water) pressure is reduced. Water lilies (water lily) (Nymphaeaceae) and rice crops contain hydromorphic leaves.
Xeromorphic adaptations to arid conditions are quite varied and tend to prevent water loss during periods when water is limited and must be conserved by the plant. There are many modifications limiting transpiration: two examples are a multilayered epidermis covered by thick layers of epicuticular wax or mucilages secreted into stomates; another is dense mats of trichomes on both surfaces of the leaf and guard cells and stomata sunken into the lower surface and often lined with numerous trichomes, which trap moisture, thereby inhibiting total water loss. Mesophyll modifications provide a means of storing water. Most xeromorphic leaves have a high volume-to-surface ratio—i.e., they are small and compact. In addition, many are fleshy and often oval to round in shape.
 The development of monocotyledonous leaves after initiation of cell division on the shoot apical meristem is different from that of dicotyledons and results in leaves with different morphologies from those of dicotyledons. Leaves in monocotyledons have either a radial leaf tip or are expanded in the same plane as the stems instead of at a right angle to the stem, as in dicotyledons. The leaf buttress begins as a ring that encloses the stem. The upper portion of the buttress develops a meristem on the side facing the stem (adaxial meristem). Growth at this adaxial meristem forms the flattened leaf with the radial (cylindrical) leaf tip typical of the monocotyledons. If the adaxial meristem is long-lived (Figure 10-->), long flat leaves in the same plane as the stem are formed ( Iris; Iridaceae); if short-lived (Figure 10-->), flat leaves with short cylindrical tips develop (snake plant, Sansevieria trifasciata; Agavaceae). When the radial (topmost aspect of the leaf) is short, the base becomes flattened because the marginal meristems (those on either side of the midvein) continue to expand outward. A monocot leaf grows either radially or along the margins, but not both in the same region. The monocot leaf grows in length from a meristem at its base, which is why it is possible to mow grass and have the leaf blades continue to grow.
The development of monocotyledonous leaves after initiation of cell division on the shoot apical meristem is different from that of dicotyledons and results in leaves with different morphologies from those of dicotyledons. Leaves in monocotyledons have either a radial leaf tip or are expanded in the same plane as the stems instead of at a right angle to the stem, as in dicotyledons. The leaf buttress begins as a ring that encloses the stem. The upper portion of the buttress develops a meristem on the side facing the stem (adaxial meristem). Growth at this adaxial meristem forms the flattened leaf with the radial (cylindrical) leaf tip typical of the monocotyledons. If the adaxial meristem is long-lived (Figure 10-->), long flat leaves in the same plane as the stem are formed ( Iris; Iridaceae); if short-lived (Figure 10-->), flat leaves with short cylindrical tips develop (snake plant, Sansevieria trifasciata; Agavaceae). When the radial (topmost aspect of the leaf) is short, the base becomes flattened because the marginal meristems (those on either side of the midvein) continue to expand outward. A monocot leaf grows either radially or along the margins, but not both in the same region. The monocot leaf grows in length from a meristem at its base, which is why it is possible to mow grass and have the leaf blades continue to grow.The developmental pattern from a basal intercalary meristem has placed constraints on the anatomy of monocot leaves, particularly with respect to venation and the position of stomates. This has produced a leaf anatomy characteristic of the monocots. There is no midvein, and veins are longitudinally parallel. The stomates are in rows between the veins, and the mesophyll is often poorly developed and mostly parenchymatous with scattered bundles of fibres. Thus, most monocot leaves are uniform in appearance and texture. Most of the hydromorphic and xeromorphic modifications found among dicot leaves, however, also occur in monocot leaves in similar environments.
Reproductive (reproductive system, plant) structures
General features
The broad range of variation in the morphology and structure of nonreproductive (vegetative) organs within the angiosperms has been outlined above. There is a similarly broad range in the morphology and structure of the reproductive organs of the plant.
Many vegetative buds (bud) sooner or later become flower buds. Flower buds are modified leaves borne on a short axis with very short internodes and no axillary buds. The floral axis has determinate growth, in that at some point it ceases to grow.
Flowers (flower), the reproductive tissues of the plant, contain the male and/or female organs. They may terminate short lateral branches or the main axis or both. Flowers may be borne singly (as in the daffodil and Magnolia) or in clusters called inflorescences (inflorescence) (e.g., bromeliads, snapdragons, and sunflowers). Fruits are derived from the floral parts of the angiospermous plant.
 A complete flower is composed of four organs attached to the floral stalk by a receptacle (Figure 11-->). From the base of the receptacle upward these four organs are the sepals, petals, stamens, and carpels (carpel). In dicots the organs are generally grouped in multiples of four or five (rarely in threes), and in monocots they are grouped in multiples of three.
A complete flower is composed of four organs attached to the floral stalk by a receptacle (Figure 11-->). From the base of the receptacle upward these four organs are the sepals, petals, stamens, and carpels (carpel). In dicots the organs are generally grouped in multiples of four or five (rarely in threes), and in monocots they are grouped in multiples of three.The sepals, the outermost layer, are usually green, enclose the flower bud, and collectively are called the calyx. Petals are the next layer of floral appendages internal to the calyx; they are generally brightly coloured and collectively are called the corolla. The calyx and corolla together compose the perianth. The sepals and petals are accessory parts or sterile appendages; though they protect the flower buds and attract pollinators, they are not directly involved with sexual reproduction. When the colour and appearance of sepals and petals are similar, as in the tulip tree (Liriodendron tulipifera) and Easter lily (Lilium longiflorum), the perianth is said to be composed of tepals.
Internal to the corolla are the stamens, spore-producing structures (microsporophylls) that are collectively called the androecium. In most angiosperms, the stamens consist of a slender stalk (the filament) that bears the anther (and pollen sacs), within which the pollen is formed. Small secretory structures called nectaries are often found at the base of the stamens and provide food rewards for pollinators. In some cases the nectaries coalesce into a nectary or staminal disc. In many cases the staminal disc forms when a whorl of stamens is reduced into a nectiferous disc, and in other cases the staminal disc is actually derived from nectary-producing tissue of the receptacle.
At the centre of the flower are the carpels, collectively called the gynoecium. Carpels are megasporophylls that enclose one or more ovules, each with an egg. After fertilization, the ovule matures into a seed, and the carpel matures into a fruit. Carpels, and thus fruit, are unique to angiosperms.
A complete flower contains all four organs, while an incomplete flower is missing at least one. A bisexual (or “perfect”) flower has both stamens and carpels, and a unisexual (or “imperfect”) flower either lacks stamens (and is called carpellate) or lacks carpels (and is called staminate). Species with both staminate flowers and carpellate flowers on the same plant (e.g., corn) are monoecious (hermaphroditism), from the Greek for “one house.” Species in which the staminate flowers are on one plant and the carpellate flowers are on another are dioecious, from the Greek for “two houses.”
 Floral organs are often united or fused: connation is the fusion of similar organs—e.g., the fused petals in the morning glory; adnation is the fusion of different organs—for example, the stamens fused to petals in the mint family ( Lamiaceae). The basic floral pattern consists of alternating whorls of organs positioned concentrically: from outside inward, sepals, petals, stamens, and carpels (Figure 12-->). It is possible in most cases to interpret the flower with respect to missing parts and/or the modification of parts to function as missing parts simply by positional relationships. In a complete five-merous flower (starting from the outside) there would be a whorl of five sepals, followed by an alternating whorl of five petals, followed by an alternating set of five stamens. In the floral diagram (Figure 12-->), the midline of each petal is midway between the midlines of two adjacent sepals. Because the whorls alternate, the midline of each stamen of the stamen whorl is between the midlines of two adjacent petals and on the midline of each sepal. When the petals are missing and bracts appear coloured and petaloid as in the Bougainvillea, one of the three whorls is missing: there are only two whorls of five organs instead of the three whorls of five organs described above. Because one whorl of the flower is obviously composed of stamens that bear functional pollen and the other whorl is composed of a brightly coloured set of organs that resemble petals one might conclude that the sepals are missing. But examination of positional relationships between the whorls reveals that the midline of each stamen is on the same line as the midline of the organs of the brightly coloured set. Thus, position tells us that the brightly coloured whorl represents a sepal whorl and that the sepals have assumed the function of the missing petals.
Floral organs are often united or fused: connation is the fusion of similar organs—e.g., the fused petals in the morning glory; adnation is the fusion of different organs—for example, the stamens fused to petals in the mint family ( Lamiaceae). The basic floral pattern consists of alternating whorls of organs positioned concentrically: from outside inward, sepals, petals, stamens, and carpels (Figure 12-->). It is possible in most cases to interpret the flower with respect to missing parts and/or the modification of parts to function as missing parts simply by positional relationships. In a complete five-merous flower (starting from the outside) there would be a whorl of five sepals, followed by an alternating whorl of five petals, followed by an alternating set of five stamens. In the floral diagram (Figure 12-->), the midline of each petal is midway between the midlines of two adjacent sepals. Because the whorls alternate, the midline of each stamen of the stamen whorl is between the midlines of two adjacent petals and on the midline of each sepal. When the petals are missing and bracts appear coloured and petaloid as in the Bougainvillea, one of the three whorls is missing: there are only two whorls of five organs instead of the three whorls of five organs described above. Because one whorl of the flower is obviously composed of stamens that bear functional pollen and the other whorl is composed of a brightly coloured set of organs that resemble petals one might conclude that the sepals are missing. But examination of positional relationships between the whorls reveals that the midline of each stamen is on the same line as the midline of the organs of the brightly coloured set. Thus, position tells us that the brightly coloured whorl represents a sepal whorl and that the sepals have assumed the function of the missing petals.The receptacle
The receptacle is the axis (stem) to which the floral organs are attached. Floral organs are attached either in a low continuous spiral, as is common among primitive angiosperms, or in alternating successive whorls, as is found among most angiosperms.
The peduncle is the stalk of a flower or an inflorescence. When a flower is borne singly, the internode between the receptacle and the bract (the last leaf, often modified and usually smaller than the other leaves) is the peduncle. When the flowers are borne in an inflorescence, the peduncle is the internode between the bract and the inflorescence; the internode between the receptacle of each flower and its underlying bracteole is called a pedicel. Thus, in inflorescences, bracteole is the equivalent of bract, and pedicel is the equivalent of peduncle.
 Often the bract subtending an inflorescence is brightly coloured, as in the poinsettia (Euphorbia pulcherrima; Euphorbiaceae), or provides protection, as in the woody, boat-shaped bracts in many palms (palm). Bracteoles in the inflorescence of Bougainvillea also are brightly coloured to attract pollinators (see photograph-->). In some angiosperms, the receptacle becomes fleshy; in the strawberry, for example, the receptacle is the fleshy edible part of the strawberry and, when eaten by small mammals and birds, aids in seed dispersal. In others, the peduncle or pedicel becomes fleshy; in the cashew (Anacardium occidentale; Anacardiaceae), for example, the pedicel is made into a drink in the Neotropics, and it also aids in fruit dispersal of the much smaller cashew nut. In cacti (cactus) (e.g., prickly pear), the fleshy part of the edible fruit forms from the receptacle and peduncle, and several internodes below that grow up and surround the carpels; this is why there are axillary buds in cacti (areoles) with spines on the fruit surface.
Often the bract subtending an inflorescence is brightly coloured, as in the poinsettia (Euphorbia pulcherrima; Euphorbiaceae), or provides protection, as in the woody, boat-shaped bracts in many palms (palm). Bracteoles in the inflorescence of Bougainvillea also are brightly coloured to attract pollinators (see photograph-->). In some angiosperms, the receptacle becomes fleshy; in the strawberry, for example, the receptacle is the fleshy edible part of the strawberry and, when eaten by small mammals and birds, aids in seed dispersal. In others, the peduncle or pedicel becomes fleshy; in the cashew (Anacardium occidentale; Anacardiaceae), for example, the pedicel is made into a drink in the Neotropics, and it also aids in fruit dispersal of the much smaller cashew nut. In cacti (cactus) (e.g., prickly pear), the fleshy part of the edible fruit forms from the receptacle and peduncle, and several internodes below that grow up and surround the carpels; this is why there are axillary buds in cacti (areoles) with spines on the fruit surface.The calyx
 The sepals (collectively called the calyx) most resemble leaves because of their generally green colour. From their base and along most of their length, sepals remain either separate (aposepalous, or polysepalous) or marginally fused (synsepalous), forming a tube with terminal lobes or teeth (see photograph-->). The number of calyx lobes equals the number of fused (connate) sepals.
The sepals (collectively called the calyx) most resemble leaves because of their generally green colour. From their base and along most of their length, sepals remain either separate (aposepalous, or polysepalous) or marginally fused (synsepalous), forming a tube with terminal lobes or teeth (see photograph-->). The number of calyx lobes equals the number of fused (connate) sepals.The sepals enclose and protect the unopened flower bud. The calyx is commonly persistent and evident when the fruit matures (e.g., persimmon, Diospyros virginiana; Ebenaceae), in contrast to the more short-lived petals and stamens. Sepals may be brightly coloured and function as petals when true petals are missing—for example, the virgin's bower (Clematis; Ranunculaceae) and the Bougainvillea. Petaloid sepals in this case differ from tepals because the first group of stamens are on the same radii as the sepals, indicating the absence of the petals, which would normally be positioned on alternating radii in the next floral whorl.
The corolla

 The petals composing the corolla are typically brightly coloured or white and attract insects and birds for pollination (see below Reproduction: Pollination (angiosperm)). The number of petals is usually the same as the number of sepals. Floral symmetry is defined by the petals (Figure 13-->). When the petals of the corolla are of the same size and shape and when they are equidistant from each other, the flower has radial symmetry, and the flower is called regular or actinomorphic (e.g., buttercup, Ranunculus; Ranunculaceae). In regular flowers, any line drawn through the centre will divide the flower into two identical halves. When at least one petal of the corolla is different, the flower has bilateral symmetry and is called irregular or zygomorphic (e.g., violets, Viola; Violaceae 【see photograph-->】).
The petals composing the corolla are typically brightly coloured or white and attract insects and birds for pollination (see below Reproduction: Pollination (angiosperm)). The number of petals is usually the same as the number of sepals. Floral symmetry is defined by the petals (Figure 13-->). When the petals of the corolla are of the same size and shape and when they are equidistant from each other, the flower has radial symmetry, and the flower is called regular or actinomorphic (e.g., buttercup, Ranunculus; Ranunculaceae). In regular flowers, any line drawn through the centre will divide the flower into two identical halves. When at least one petal of the corolla is different, the flower has bilateral symmetry and is called irregular or zygomorphic (e.g., violets, Viola; Violaceae 【see photograph-->】). The petals of the corolla may be separate, or apopetalous, or marginally fused (fusion of like floral parts is called connation), or sympetalous, for all or part of their length. When joined, they form a tubular corolla with terminal lobes (see photograph-->). A tubular corolla may be present in regular flowers (e.g., blueberries (blueberry), Vaccinium; Ericaceae) or irregular flowers (e.g., sage, Salvia officinalis; Lamiaceae). Stamens are commonly united to a tubular corolla (fusion of two unlike floral parts is called adnation). A marginally fused (synsepalous) calyx, a marginally fused (sympetalous) corolla, and stamens may fuse to form a cuplike floral tube called a hypanthium that surrounds the carpels, as in cherries (cherry) (Prunus; Rosaceae), for example. Fusion and reduction of flower parts are common and have occurred in many unrelated lineages. Many wind-pollinated angiosperms do not have petals, nor do they have floral parts modified as petals; examples of wind-pollinated species include the amaranth family (Amaranthaceae) and the birch family (Betulaceae).
The petals of the corolla may be separate, or apopetalous, or marginally fused (fusion of like floral parts is called connation), or sympetalous, for all or part of their length. When joined, they form a tubular corolla with terminal lobes (see photograph-->). A tubular corolla may be present in regular flowers (e.g., blueberries (blueberry), Vaccinium; Ericaceae) or irregular flowers (e.g., sage, Salvia officinalis; Lamiaceae). Stamens are commonly united to a tubular corolla (fusion of two unlike floral parts is called adnation). A marginally fused (synsepalous) calyx, a marginally fused (sympetalous) corolla, and stamens may fuse to form a cuplike floral tube called a hypanthium that surrounds the carpels, as in cherries (cherry) (Prunus; Rosaceae), for example. Fusion and reduction of flower parts are common and have occurred in many unrelated lineages. Many wind-pollinated angiosperms do not have petals, nor do they have floral parts modified as petals; examples of wind-pollinated species include the amaranth family (Amaranthaceae) and the birch family (Betulaceae).Petals often bear nectaries that secrete sugar-containing compounds, and petals also produce fragrances to attract pollinators; the fragrance of a rose (Rosa; Rosaceae) is derived from the petals. Petals often develop a nectar-containing extension of the tubular corolla, called a spur. This may involve one petal, as in the larkspur (Delphinium), or all the petals, as in columbine (Aquilegia), both members of the family Ranunculaceae.
The androecium
Stamens (microsporophylls) are structures that produce pollen in terminal saclike structures (microsporangia) called anthers. The number of stamens comprised by the androecium is sometimes the same as the number of petals, but often the stamens are more numerous or fewer in number than the petals. There are generally two pairs of spore-containing sacs (microsporangia) in a young stamen; during maturation the partition between the adjacent microsporangia of a pair breaks down so that there are only two pollen-containing sacs (one in each lobe of the anther) at the time the stamen releases the pollen.
The least-modified stamens are similar to leaves, with the paired microsporangia located at or near the margins; an example is found in the magnolia family (Magnoliaceae). In more derived stamens, the blade has become modified into a slender stalk, the filament, with the microsporangia at or near the filament apex (the anther). The filaments are very often united with the corolla, but with the anthers either separate, as in primroses (primrose) (Primula; Primulaceae), or united with each other to form a staminal tube that encloses the gynoecium, as in the mallow family ( Malvaceae). In thistle (Cirsium; Asteraceae) and in other members of the sunflower family (Asteraceae), the staminal tube is fused to the lower half of the corolla tube.
 There are several trends in stamen modification. In many angiosperms, one or more of the stamens is modified and lacks functional anthers. In the most common modification, the filament is expanded to form a petallike blade called a staminode (in the same manner that a sepal forms a petallike blade in some flowers without true petals). The apparent petals in some angiosperm families, such as are found in many members of the pink family (Caryophyllaceae), are staminodial in origin. Wild roses have only five petals and many stamens; however, cultivated roses have been selected for the many apparent petals (but actually staminodes) and few functional stamens. In other cases, stamens have been modified into sterile nectaries involved in pollination. If flowers have a large number of stamens, then the stamens often occur in groups or clusters (fascicles; see photograph-->), as in the myrtle family (Myrtaceae).
There are several trends in stamen modification. In many angiosperms, one or more of the stamens is modified and lacks functional anthers. In the most common modification, the filament is expanded to form a petallike blade called a staminode (in the same manner that a sepal forms a petallike blade in some flowers without true petals). The apparent petals in some angiosperm families, such as are found in many members of the pink family (Caryophyllaceae), are staminodial in origin. Wild roses have only five petals and many stamens; however, cultivated roses have been selected for the many apparent petals (but actually staminodes) and few functional stamens. In other cases, stamens have been modified into sterile nectaries involved in pollination. If flowers have a large number of stamens, then the stamens often occur in groups or clusters (fascicles; see photograph-->), as in the myrtle family (Myrtaceae).The gynoecium
The gynoecium is composed of carpels. In more basal families (e.g., Magnoliaceae) the carpels are spirally arranged, and in more advanced families they tend to be arranged in a single whorl. Carpel number varies from one (e.g., bean or legume family 【Fabaceae】) to many (e.g., buttercups or raspberries 【Rubus】).
At the base of a carpel is the ovary, within which develop one or more multicellular structures called ovules that each contain an egg. The upper part of the carpel, the stigma, receives the pollen. A slender stalk called the style often connects the ovary and stigma. The carpels may be separate (apocarpous) or fused together (syncarpous), with the individual carpel walls and cavities (locules) still present. Syncarpy may involve only the ovaries, leaving the styles and stigmas free, as is found in the wood sorrel (Oxalis), or it may involve both the ovaries and styles, leaving only the stigmas free, as in the waterleaf (Hydrophyllum). The number of carpels in a syncarpous (or compound) ovary generally equals the number of locules (in some cases the inner carpel walls break down, leaving a single locule); in an orange or a grapefruit, for example, the juice sacs are actually trichomes that line the inner carpel walls of each cavity.
The position of the gynoecium with respect to the petals, sepals, and stamens on the floral axis also characterizes the flower (Figure 11-->). In hypogynous flowers, the perianth and stamens are attached to the receptacle below the gynoecium; the ovary is superior to these organs, and the remaining floral organs arise from below the point of origin of the carpel. In perigynous flowers, a hypanthium (a floral tube formed from the fusion of the stamens, petals, and sepals) is attached to the receptacle below the gynoecium and surrounds the ovary; the ovary is superior, and the free parts of the petals, sepals, and stamens are attached to the rim of the hypanthium. In epigynous flowers, the hypanthium is fused to the gynoecium, and the free parts of the sepals, petals, and stamens appear to be attached to the top of the gynoecium, as in the apple (Malus; Rosaceae); the ovary is inferior, and the petals, sepals, and stamens appear to arise from the top of the ovary.Fruits (fruit)
Fertilization of an egg within a carpel by a compatible pollen grain results in seed development within the carpel. (Formation of fruit without the fertilization of an egg and subsequent seed development is called parthenocarpy.) A fruit is a ripened ovary (or compound ovary) and any other structure, usually the hypanthium, that ripens and forms a unit with it. This clearly separates a fruit from a vegetable, because a vegetable is derived only from vegetative (nonreproductive) organs. Tomatoes, eggplants, and squashes are fruits, because they are derived from floral parts, whereas carrots, turnips (Brassica rapa), and beets are vegetables, because they are roots modified as storage organs in the same manner that potatoes, ginger (Zingiber officinale), and onions are modified stems.
Simple fruits develop from a single carpel or from a compound ovary. Aggregate fruits consist of several separate carpels of one apocarpous gynoecium (e.g., raspberries (raspberry) where each unit is a single carpel). Multiple fruits consist of the gynoecia of more than one flower and represent a whole inflorescence, such as the fig and pineapple. Accessory fruits incorporate other flower parts in the development of the mature fruit; for example, the hypanthium is used in forming the pear (Pyrus; Rosaceae), and the receptacle becomes part of the prickly pear.
The form, texture, and structure of fruits are varied (notably in simple fruits), but most fall within a few categories. The fruit wall, or pericarp, is divided into three regions: the inner layer, or endocarp; the middle layer, or mesocarp; and the outer layer, or exocarp. These regions may be fleshy or dry (sclerified) or any combination of the two, but they are classified as either one or the other.
The three main types of fleshy fruits are berries (berry), drupes, and pomes. Berries are many-seeded simple fruits composed of one carpel or a syncarpous ovary. They are fleshy throughout, but the exocarp ranges in texture: a soft, thin exocarp, as in tomatoes (a berry); a leathery exocarp, as in oranges (a hesperidium); and a somewhat hard exocarp, as in pumpkins (a pepo). In drupes (drupe), or stone fruits, there is usually only one seed per carpel or locule. Drupes are fleshy fruits and consist of an inner stony or woody endocarp, which adheres to the seed (peaches, plums, and cherries). The term druplet is used for each unit of aggregate fruit of this type (e.g., raspberries and blackberries). Pomes are fleshy fruits of the rose family (Rosaceae) in which an adnate hypanthium becomes fleshy (apples and pears).
Simple dry fruits are either dehiscent or indehiscent. They are dehiscent if the pericarp splits open at maturity and releases the seeds, or indehiscent if the pericarp remains intact when the fruit is shed from the plant. The three principal types of dehiscent fruits are follicles, legumes (legume), and capsules (capsule). Follicles and legumes are each derived from an ovary with a single carpel, and a capsule is derived from several united carpels. As the fruit matures, the pericarp dries and the fruit splits. Whereas follicles split along a single side of the fruit, such as in the milkweeds, columbines, and magnolias, legumes split along both sides, as in the bean family. Capsules have two or more carpels and split open to release their seeds in various ways. They may open longitudinally to expose the seeds within each locule (cavity) or longitudinally along each septum between the locules, as in the agave (Agave; Agavaceae). Still others form an operculum (a lid) at the top of the ovary, as in the Brazil nut family (Lecythidaceae).
Indehiscent fruits are derived from either single carpels or compound ovaries. Single carpel forms include the achene, the samara, and the caryopsis. Forms derived from a compound ovary include nuts and schizocarps. An achene is a fruit in which the single seed lies free in the cavity, attached only by a single point. The strawberry, for example, is really an aggregate fruit, and each “seed” is an achene. The samara is a winged achene and is found in the tree of heaven (Ailanthus altissima; Simaroubaceae) and ash (Fraxinus; Oleaceae). In the caryopsis, or grain, the seed adheres to the fruit wall (pericarp). The caryopsis is found among the cereal grasses, such as corn. Nuts (nut) have a stony pericarp, and usually only a single seed in each carpel matures, as in acorns of oaks (Quercus; Fagaceae) and hazelnuts (Corylus avellana; Betulaceae). Schizocarps are fruits in which each carpel of a compound ovary splits apart to form two or more parts, each with a single seed (seed and fruit). Schizocarps are found in the carrot family (Apiaceae). Winged schizocarps are found in maples.
Seeds
Seeds are the mature ovules. They contain the developing embryo and the nutritive tissue for the seedling. Seeds are surrounded by one or two integuments (integument), which develop into a seed coat that is usually hard. They are enclosed in the ovary of a carpel and thus are protected from the elements and predators.
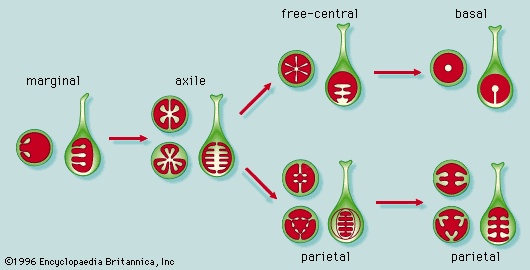 The ovule is attached to the ovary wall until maturity by a short stalk called the funiculus. The area of attachment to the ovary wall is referred to as the placenta. The arrangement of placentae (placentation) in the compound ovary of angiosperms is characterized by the presence or absence of a central column in the ovary and by the site of attachment (Figure 14-->). In axile placentation the placentae are located on a central column; partitions from the central column to the ovary wall create chambers (locules) that separate the placentae and attached ovaries from each other. Free-central placentation resembles axile placentation; however, the column is not connected by partitions to the ovary wall, and thus no locules are formed. In basal placentation ovules are attached to the base of the ovary, and in parietal placentation the placentae are located directly on the ovary wall or on its extensions.
The ovule is attached to the ovary wall until maturity by a short stalk called the funiculus. The area of attachment to the ovary wall is referred to as the placenta. The arrangement of placentae (placentation) in the compound ovary of angiosperms is characterized by the presence or absence of a central column in the ovary and by the site of attachment (Figure 14-->). In axile placentation the placentae are located on a central column; partitions from the central column to the ovary wall create chambers (locules) that separate the placentae and attached ovaries from each other. Free-central placentation resembles axile placentation; however, the column is not connected by partitions to the ovary wall, and thus no locules are formed. In basal placentation ovules are attached to the base of the ovary, and in parietal placentation the placentae are located directly on the ovary wall or on its extensions.Mature seeds are enclosed in integuments that may become hard and stony or that may have an outer fleshy, usually brightly coloured sarcotesta with an inner stony sclerotesta. Seed coats also may be winged or variously ornamented with prickles or sclerified hairs. In some seeds, there may be an extra covering, the aril, which is an outgrowth of the funiculus (e.g., the spice mace is derived from the red aril of Myristica fragrans; Myristicaceae). The aril of tomato seeds makes them slippery.
Mechanisms of dispersal
Fruits and seeds are the primary means by which angiosperms are dispersed (dispersion). The chief agents of dispersal are wind, water, and animals. Some fruits and seeds have modifications that aid in wind dispersal. Fruit modifications include samaras, samaroid schizocarps, and the feathery calyx lobes (e.g., dandelion). Seeds may be modified in various ways to promote dispersal: they may be extremely small and light (e.g., orchids, Orchidaceae), winged (e.g., common catalpa, Catalpa bignonioides; Bignoniaceae), plumed (e.g., milkweed), covered with woolly hairs (e.g., willows), or surrounded by explosive capsules that forcefully eject them into the air, as, for example, the touch-me-not (Impatiens; Balsaminaceae) and the witch hazel (Hamamelis; Hamamelidaceae). The fruits or seeds of many aquatic and shore plants are adapted to float on water as a means of dispersal; for this reason, coconuts (Cocos nucifera (coconut palm); Arecaceae) are readily transported across oceans to neighbouring islands. Adaptations for water dispersal include aerenchyma in fruits or seeds and light weight (e.g., water chestnut, Trapa natans; Lythraceae).
Animals (animal) disperse fruits and seeds either by ingesting and subsequently excreting them or by passively transporting them once they have adhered to an external part of the body, such as the fur or a claw. The evolution of fleshy fruits and seeds exemplifies the coevolution of plants and their animal agents of dispersal. An animal diet often consists solely of fruits and seeds that are designed to be eaten and dispersed, and in many cases these seeds require full or partial digestion to stimulate germination. Most fruits with a fleshy pericarp are eaten whole by vertebrates, including the stony endocarp or the stony seed coat. The seeds then either are regurgitated by the animal or pass through the alimentary canal and are excreted, often some distance from the original site. Seeds with an aril that encloses a stony seed coat or seeds with a sclerotesta and a fleshy, coloured sarcotesta are found in dehiscent fruits. They are eaten by animals after the fruit has ripened and split open. Often these seeds dangle from the fruit by long stalks (e.g., the follicles of Magnolia). The fleshy portion, whether originally a fruit or seed, is brightly coloured and sweet so as to attract vertebrates, particularly birds and mammals. Many fruits and seeds in the Amazon, however, are actually eaten and dispersed by fish during times of high water.
Inflorescences (inflorescence)
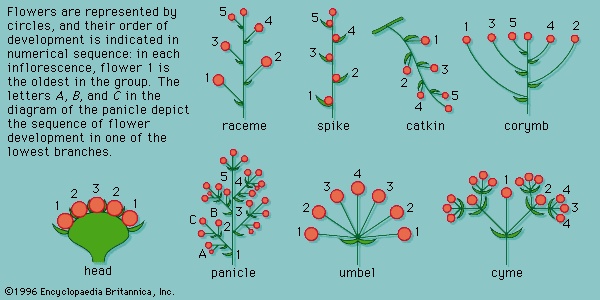 Inflorescences are clusters of flowers on a branch or a system of branches. They are categorized generally on the basis of the timing of their flowering and by their arrangement on an axis. In indeterminate inflorescences, the youngest flowers, and therefore the last to open, are either at the top of the inflorescence (in elongated axes) or in the centre (in truncated axes). Branching and the associated flowers develop at some distance from the main stem (monopodial growth). Indeterminate inflorescences are of varied types (Figure 15-->): racemes, panicles, spikes, catkins (or aments), corymbs, and heads.
Inflorescences are clusters of flowers on a branch or a system of branches. They are categorized generally on the basis of the timing of their flowering and by their arrangement on an axis. In indeterminate inflorescences, the youngest flowers, and therefore the last to open, are either at the top of the inflorescence (in elongated axes) or in the centre (in truncated axes). Branching and the associated flowers develop at some distance from the main stem (monopodial growth). Indeterminate inflorescences are of varied types (Figure 15-->): racemes, panicles, spikes, catkins (or aments), corymbs, and heads.


 A raceme is an inflorescence in which a flower develops at the axil of each leaf along an elongated, unbranched axis (see photograph-->). Each flower terminates a short stalk called a pedicel. The main axis has indeterminate growth; therefore, its growth does not cease at the onset of flowering. A spike is a raceme except that the flowers are attached directly to the axis at the axil of each leaf rather than to a pedicel (see photograph-->). An example of a spike is the cattail (Typha; Typhaceae). The fleshy spike characteristic of the Araceae is called a spadix, and the underlying bract is known as a spathe. A catkin (or ament) is a spike in which all the flowers are of only one sex, either staminate or carpellate. The catkin is usually pendulous, and the petals and sepals are reduced to aid in wind pollination when the inflorescence as a whole is shed (see photograph-->). An example of a catkin is found in oaks. A corymb is a raceme in which the pedicels of the lower flowers are longer than those of the upper ones so that the appearance of the inflorescence overall is that of a flat flower (see photograph-->). The lower flowers open first, and the axis of a corymb continues to produce flowers (indeterminate growth). Corymbs are found in the hawthorn (Crataegus; Rosaceae).
A raceme is an inflorescence in which a flower develops at the axil of each leaf along an elongated, unbranched axis (see photograph-->). Each flower terminates a short stalk called a pedicel. The main axis has indeterminate growth; therefore, its growth does not cease at the onset of flowering. A spike is a raceme except that the flowers are attached directly to the axis at the axil of each leaf rather than to a pedicel (see photograph-->). An example of a spike is the cattail (Typha; Typhaceae). The fleshy spike characteristic of the Araceae is called a spadix, and the underlying bract is known as a spathe. A catkin (or ament) is a spike in which all the flowers are of only one sex, either staminate or carpellate. The catkin is usually pendulous, and the petals and sepals are reduced to aid in wind pollination when the inflorescence as a whole is shed (see photograph-->). An example of a catkin is found in oaks. A corymb is a raceme in which the pedicels of the lower flowers are longer than those of the upper ones so that the appearance of the inflorescence overall is that of a flat flower (see photograph-->). The lower flowers open first, and the axis of a corymb continues to produce flowers (indeterminate growth). Corymbs are found in the hawthorn (Crataegus; Rosaceae). If the axis is short or stunted, the flowers arise from a common point and appear to be at approximately the same level. This pattern, called an umbel (see photograph-->), is actually a flattened raceme because the internodes of the axis, or peduncle (the point of origin of the leaves and flower axes), are shortened so that the pedicels are of the same length (e.g., the carrot family). A head is a raceme in which the peduncle is flattened and the flowers are attached directly to it (e.g., aster family, Asteraceae). This results in a grouping of small flowers in such a way as to appear as a single flower. In many members of the Asteraceae (e.g., sunflowers, Helianthus annuus), for instance, the outer (or ray) flowers have a well-developed zygomorphic corolla, and the inner (disk) flowers have a small actinomorphic corolla. The inner disk flowers generally are complete flowers, and the ray flowers generally are sterile.
If the axis is short or stunted, the flowers arise from a common point and appear to be at approximately the same level. This pattern, called an umbel (see photograph-->), is actually a flattened raceme because the internodes of the axis, or peduncle (the point of origin of the leaves and flower axes), are shortened so that the pedicels are of the same length (e.g., the carrot family). A head is a raceme in which the peduncle is flattened and the flowers are attached directly to it (e.g., aster family, Asteraceae). This results in a grouping of small flowers in such a way as to appear as a single flower. In many members of the Asteraceae (e.g., sunflowers, Helianthus annuus), for instance, the outer (or ray) flowers have a well-developed zygomorphic corolla, and the inner (disk) flowers have a small actinomorphic corolla. The inner disk flowers generally are complete flowers, and the ray flowers generally are sterile.
 In the compound indeterminate inflorescences, the main axis is branched so that the many inflorescences form off the main axis. A panicle (see photograph-->) is a branched raceme in which the branches are themselves racemes (e.g., yuccas, Yucca). In a compound umbel (see photograph-->), all the umbel inflorescences arise from a common point and appear to be at about the same level (e.g., wild carrot). This organization is the same for compound spikes, catkins, corymbs, and heads. The change from elongated axes (racemes and panicles) to flattened axes (corymbs and umbels) results in inflorescences in which the flowers are arranged close together. This close association encourages efficient pollination, and the extreme condensation of the inflorescences, as in the head, gives rise to an inflorescence that appears to be a single flower (e.g., sunflowers).
In the compound indeterminate inflorescences, the main axis is branched so that the many inflorescences form off the main axis. A panicle (see photograph-->) is a branched raceme in which the branches are themselves racemes (e.g., yuccas, Yucca). In a compound umbel (see photograph-->), all the umbel inflorescences arise from a common point and appear to be at about the same level (e.g., wild carrot). This organization is the same for compound spikes, catkins, corymbs, and heads. The change from elongated axes (racemes and panicles) to flattened axes (corymbs and umbels) results in inflorescences in which the flowers are arranged close together. This close association encourages efficient pollination, and the extreme condensation of the inflorescences, as in the head, gives rise to an inflorescence that appears to be a single flower (e.g., sunflowers).

 In the determinate (cymose) inflorescences, the youngest flowers (those that are the last to open) are at the bottom of an elongated axis or on the outside of a truncated axis (e.g., in the cymose umbel of onions, Allium; Alliaceae). These inflorescences are determinate because, at the time of flowering, the whole apical meristem produces a flower; thus, the entire axis ceases to grow. Each unit of a cyme consists of a dichasium, which has a central flower and two lateral flowers (see photograph-->). The branching is primarily sympodial, and the inflorescence may be compound (e.g., catchfly, or campion, Silene; Caryophyllaceae). Many monocotyledons have a one-sided cyme called a helicoid cyme (see photograph-->). A cymose inflorescence arranged in pairs at the nodes, in the manner of a false whorl, is called a verticillaster (see photograph-->). Finally, there are mixed inflorescences, as, for instance, the cymose clusters arranged in a racemose manner (e.g., lilac, Syringa vulgaris; Oleaceae) or other types of combinations.
In the determinate (cymose) inflorescences, the youngest flowers (those that are the last to open) are at the bottom of an elongated axis or on the outside of a truncated axis (e.g., in the cymose umbel of onions, Allium; Alliaceae). These inflorescences are determinate because, at the time of flowering, the whole apical meristem produces a flower; thus, the entire axis ceases to grow. Each unit of a cyme consists of a dichasium, which has a central flower and two lateral flowers (see photograph-->). The branching is primarily sympodial, and the inflorescence may be compound (e.g., catchfly, or campion, Silene; Caryophyllaceae). Many monocotyledons have a one-sided cyme called a helicoid cyme (see photograph-->). A cymose inflorescence arranged in pairs at the nodes, in the manner of a false whorl, is called a verticillaster (see photograph-->). Finally, there are mixed inflorescences, as, for instance, the cymose clusters arranged in a racemose manner (e.g., lilac, Syringa vulgaris; Oleaceae) or other types of combinations. reproduction
General features
The vast array of angiosperm floral structures is for sexual reproduction. The angiosperm life cycle consists of a sporophyte phase and a gametophyte phase. The cells of a sporophyte body have a full complement of chromosomes (i.e., the cells are diploid, or 2n); the sporophyte is the typical plant body that we see when we look at an angiosperm. The gametophyte arises when cells of the sporophyte, in preparation for reproduction, undergo meiotic division and produce reproductive cells that have only half the number of chromosomes (i.e., haploid, or n). A two-celled microgametophyte called a pollen grain germinates into a pollen tube and through division produces the haploid sperm. (The prefix micro- denotes gametophytes emanating from a male reproductive organ.) An eight-celled megagametophyte called the embryo sac produces the egg. (The prefix mega- denotes gametophytes emanating from female reproductive organs.)
Angiosperms are vascular plants, and all vascular plants have a life cycle in which the sporophyte phase (vegetative body) is the dominant phase and the gametophyte phase remains diminutive. In the nonvascular plants, such as the bryophytes, the gametophyte phase is dominant over the sporophyte phase. In bryophytes, the gametophyte produces its food by photosynthesis (is autotrophic) while the nongreen sporophyte is dependent on the food produced by the gametophyte. In nonseed vascular plants, such as ferns and horsetails, both the gametophyte and sporophyte are green and photosynthetic, and the gametophyte is small and without vascular tissue. In the seed plants (gymnosperms and angiosperms), the sporophyte is green and photosynthetic and the gametophyte depends on the sporophyte for nourishment. Within the seed plants, the gametophyte has become further reduced, with fewer cells comprising the gametophyte. The microgametophyte (pollen grain), therefore, is reduced from between 4 and 8 cells in the gymnosperms to a 3-celled microgametophyte in the angiosperms. A parallel reduction in the number of cells comprising a megagametophyte (ovule) has also taken place: from between 256 and several thousand cells in the gymnosperms to an 8-celled megagametophyte in most of the angiosperms. The significance of the reduction in megagametophyte cells appears to be related to pollination and fertilization. In many gymnosperms, pollination leads to the formation of a large gametophyte with copious amounts of stored starch for the nourishment of the potential embryo regardless of whether fertilization of the ovule can actually take place (i.e., whether the pollen is from the same species as the ovule). If the pollen is from a different species, fertilization or embryo development fails, so that the stored food is wasted. In angiosperms, however, the megagametophyte and egg are mature before the food is stored, and this is not ever accomplished until after the egg has been adequately fertilized and an embryo is present. This reduces the chances that the stored food will be wasted.
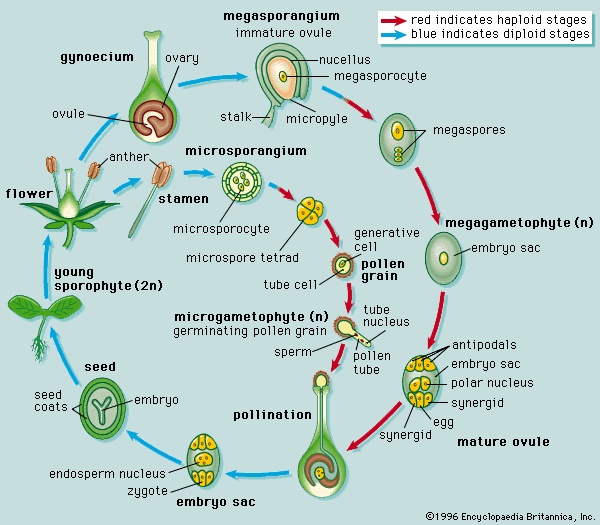 The process of sexual reproduction (Figure 16-->) depends on pollination to bring these gametophytes in close association so that fertilization can take place. Pollination is the process by which pollen that has been produced in the anthers is received by the stigma of the ovary. Fertilization occurs with the fusion of a sperm with an egg to produce a zygote, which eventually develops into an embryo. After fertilization, the ovule develops into a seed, and the ovary develops into a fruit.
The process of sexual reproduction (Figure 16-->) depends on pollination to bring these gametophytes in close association so that fertilization can take place. Pollination is the process by which pollen that has been produced in the anthers is received by the stigma of the ovary. Fertilization occurs with the fusion of a sperm with an egg to produce a zygote, which eventually develops into an embryo. After fertilization, the ovule develops into a seed, and the ovary develops into a fruit.Anthers
A transverse section of the anther reveals four areas of tissue capable of producing spores. These tissues are composed of microsporocytes, which are diploid cells capable of undergoing meiosis to form a tetrad (four joined cells) of haploid microspores. The microspores become pollen grains and may eventually separate.
During pollen development, the layer of cells beneath the dermis of the anther wall (the endothecium) develops thickenings in the cell walls. The cell layer immediately inside the endothecium (the tapetum) develops into a layer of nutritive cells that either secrete their contents into the area around the microsporocytes or lose their inner cell walls, dissociate from each other, and become amoeboid among the microsporocytes. The pollen grains develop a thick wall of at least two layers, the intine and the exine. The intine, or inner layer, consists primarily of cellulose and pectins. The exine, or outer layer, is composed of a highly decay-resistant chemical called sporopollenin. The exine usually has one or more thin areas, or pores, through which the pollen tubes germinate, and the thick area of the exine is usually highly sculptured. The number of pores and pattern of exine sculpturing are characteristic within an angiosperm family, genus, and often within a species.
The terminology to describe the various sculpturing patterns and position and number of pores is highly complex and only a basic description as related to functional aspects of sculpturing is given here. For example, smooth or essentially smooth pollen is loosely correlated with wind pollination, as in oaks (Quercus) and grasses (corn, Zea mays). Many plants pollinated by birds, insects, and small mammals have highly sculptured patterns of spines, hooks, or sticky threadlike projections by which pollen adheres to the body of the foraging pollinator as it travels to other flowers.
Each microspore (pollen grain) divides mitotically to form a two-celled microgametophyte; one cell is a tube cell (the cell that develops into a pollen tube), and the other is a generative cell, which will give rise to two sperm as a result of a further mitotic division. Thus, a mature microgametophyte consists of only three haploid cells—the tube cell and two sperm. Most angiosperms shed pollen at the two-celled stage, but in some advanced cases it is shed at the mature three-celled stage. When the pollen grains are mature, the anther wall either splits open (dehisces) longitudinally or opens by an apical pore.
Because the sporopollenin is resistant to decay, free pollen is well represented in the fossil record. The distinctive patterns of the exine are useful for identifying which species were present as well as suggesting the conditions of early climates. The proteins in the pollen walls are also a major factor in hay fever and other allergic reactions, and the spinose sculpturing patterns may cause physical irritation.
Ovules (ovule)
An ovule is a saclike structure that produces the megaspores and is enclosed by layers of cells. This megasporangium is called the nucellus in angiosperms. After initiation of the carpel wall, one or two integuments arise near the base of the ovule primordium, grow in a rimlike fashion, and enclose the nucellus, leaving only a small opening called the micropyle at the top. In angiosperms the presence of two integuments is plesiomorphic (unspecialized), and one integument is apomorphic (derived). A single large megasporocyte arises within the nucellus near the micropyle and undergoes meiotic division, resulting in a single linear tetrad of megaspores. Three of the four megaspores degenerate, and the surviving one enlarges. The resulting megagametophyte produces the female gametes (eggs). This development (called megagametogenesis) involves free-nuclear mitotic divisions. The cell wall remains intact while the nucleus divides until the megagametophyte, or embryo sac, is formed. The embryo sac typically has eight nuclei. Free-nuclear mitotic division is also found in gametophyte formation in gymnosperms.
Four nuclei migrate to either end of the embryo sac. One nucleus from each group then migrates to the centre of the embryo; these become the polar nuclei. The two polar nuclei merge to form a fusion nucleus in the centre of the embryo sac. A cell wall develops around the fusion nucleus, leaving a central cell in the sac. Cell walls form around each of the chalazal nuclei to form three antipodal cells. During development, enlargement of the embryo sac leads to the destruction of most of the nucellus. This sequence of megasporogenesis and megagametogenesis, called the Polygonum type, occurs in 70 percent of the angiosperms in which the life cycle has been charted. Variations found in the remaining 30 percent represent derivations from the Polygonum type of seed development.
pollination
Effective pollination involves the transfer of pollen from the anthers to a stigma of the same species and subsequent germination and growth of the pollen tube to the micropyle of the ovule.
Pollen transfer (pollination) is effected by wind, water, and animals, primarily insects and birds. Wind-pollinated flowers usually have an inconspicuous reduced perianth, long slender filaments and styles, covered with sticky trichomes and often branched stigmas, pendulous catkin inflorescences, and small, smooth pollen grains.
 Wind pollination is derived in angiosperms and has developed independently in several different groups. For example, within the aster family wind pollination accompanied by floral reduction has developed independently in the tribes Heliantheae and Anthemideae. Water pollination occurs in only a few aquatic plants and is highly complicated and derived.
Wind pollination is derived in angiosperms and has developed independently in several different groups. For example, within the aster family wind pollination accompanied by floral reduction has developed independently in the tribes Heliantheae and Anthemideae. Water pollination occurs in only a few aquatic plants and is highly complicated and derived.There is a wide range of animal pollinators of angiosperms as well as a wide range of adaptations by the flowers to attract those pollinators. Some of the living unspecialized families of basal angiosperms are pollinated by beetles (coleopteran). The beetles forage and feed on pieces of the perianth and stamens. There are no nectaries but rather food bodies on these organs.

 Bees (bee) are responsible for the pollination of more flowers than any other animal group. Bees usually feed on nectar and in some cases on pollen. They may be general pollinators by visiting flowers of many species, or they may have adapted (i.e., elongated) their mouthparts to different flower depths and have become specialized to pollinate only a single species. Flowers pollinated by bees commonly have a zygomorphic, or bilaterally symmetrical, corolla with a lower lip providing a landing platform for the bee (see photograph-->). Nectar is commonly produced either at the base of the corolla tube or in extensions of the corolla base. The bees partially enter the corolla mouth to feed with their long tongues on the nectar, at which point they deposit pollen picked up from other flowers and collect pollen from the new flower. Flowers pollinated by bees are often blue or yellow or exhibit patterns of both. Particular pattern markings and ultraviolet reflection patterns (see photograph-->) serve as recognition guides.
Bees (bee) are responsible for the pollination of more flowers than any other animal group. Bees usually feed on nectar and in some cases on pollen. They may be general pollinators by visiting flowers of many species, or they may have adapted (i.e., elongated) their mouthparts to different flower depths and have become specialized to pollinate only a single species. Flowers pollinated by bees commonly have a zygomorphic, or bilaterally symmetrical, corolla with a lower lip providing a landing platform for the bee (see photograph-->). Nectar is commonly produced either at the base of the corolla tube or in extensions of the corolla base. The bees partially enter the corolla mouth to feed with their long tongues on the nectar, at which point they deposit pollen picked up from other flowers and collect pollen from the new flower. Flowers pollinated by bees are often blue or yellow or exhibit patterns of both. Particular pattern markings and ultraviolet reflection patterns (see photograph-->) serve as recognition guides.
 A high degree of coevolution is common in orchids (orchid) (e.g., Ophrys speculum 【see photograph-->】), where the flower not only appears to resemble the female wasp of a particular species but also produces the pheromone released by the insect to attract males of the species. The male wasp effects pollination by pseudocopulation with the orchid flower. Other insect pollinators include flies, butterflies (see photograph-->), moths, and mosquitoes. Many flowers pollinated by flies are called carrion flowers because they look and smell like rotting meat. The skunk cabbage (Symplocarpus foetidus) and the carrion flowers (Stapelia schinzii) have evolved these characteristics independently.
A high degree of coevolution is common in orchids (orchid) (e.g., Ophrys speculum 【see photograph-->】), where the flower not only appears to resemble the female wasp of a particular species but also produces the pheromone released by the insect to attract males of the species. The male wasp effects pollination by pseudocopulation with the orchid flower. Other insect pollinators include flies, butterflies (see photograph-->), moths, and mosquitoes. Many flowers pollinated by flies are called carrion flowers because they look and smell like rotting meat. The skunk cabbage (Symplocarpus foetidus) and the carrion flowers (Stapelia schinzii) have evolved these characteristics independently. Vertebrate pollinators include birds, bats, small marsupials, and small rodents. Many bird-pollinated flowers are bright red, especially those pollinated by hummingbirds (hummingbird) (see photograph-->). Hummingbirds rely solely on nectar as their food source. Flowers (e.g., Fuchsia) pollinated by birds produce copious quantities of nectar but little or no odour because birds have a very poor sense of smell. Flowers pollinated (pollination) by bats produce large quantities of nectar and strong fragrances. They generally open only at night, when bats are the most active, and often hang down on long inflorescence stalks, which provide easy access to the nectaries and pollen. Some eucalypts (Eucalyptus) are pollinated by small marsupials (e.g., honey possums).
Vertebrate pollinators include birds, bats, small marsupials, and small rodents. Many bird-pollinated flowers are bright red, especially those pollinated by hummingbirds (hummingbird) (see photograph-->). Hummingbirds rely solely on nectar as their food source. Flowers (e.g., Fuchsia) pollinated by birds produce copious quantities of nectar but little or no odour because birds have a very poor sense of smell. Flowers pollinated (pollination) by bats produce large quantities of nectar and strong fragrances. They generally open only at night, when bats are the most active, and often hang down on long inflorescence stalks, which provide easy access to the nectaries and pollen. Some eucalypts (Eucalyptus) are pollinated by small marsupials (e.g., honey possums).Whatever the agent of dispersal, the first phase of pollination is successful when a pollen grain lands on a receptive stigma. The surface of the stigma can be wet or dry and is often composed of specialized glandular tissue; the style is lined with secretory transmitting tissue. Their secretions provide an environment that nourishes the pollen tube as it elongates and grows down the style. If mitosis in the generative cell has not yet occurred in the pollen grain, it does so at this time.
To prevent self-fertilization, many angiosperms have developed a chemical system of self-incompatibility. The most common type is sporophytic self-incompatibility, in which the secretions of the stigmatic tissue or the transmitting tissue prevent the germination or growth of incompatible pollen. A second type, gametophytic self-incompatibility, involves the inability of the gametes from the same parent plant to fuse and form a zygote or, if the zygote forms, then it fails to develop. These systems force outcrossing and maintain a wide genetic diversity.
The pollen tube ultimately enters an ovule through the micropyle and penetrates one of the sterile cells on either side of the egg (synergids). These synergids begin to degenerate immediately after pollination. Pollen tubes can reach great lengths, as in corn, where the corn silk consists of the styles for the corn ear and each silk thread contains many pollen tubes.
fertilization and embryogenesis
After penetrating the degenerated synergid, the pollen tube releases the two sperm into the embryo sac, where one fuses with the egg and forms a zygote and the other fuses with the two polar nuclei of the central cell and forms a triple fusion, or endosperm, nucleus. This is called double fertilization because the true fertilization (fusion of a sperm with an egg) is accompanied by another fusion process (that of a sperm with the polar nuclei) that resembles fertilization. Double fertilization of this type is unique to angiosperms. The zygote now has a full complement of chromosomes (i.e., it is diploid), and the endosperm nucleus has three chromosomes (triploid). The endosperm nucleus divides mitotically to form the endosperm of the seed, which is a food-storage tissue utilized by the developing embryo and the subsequent germinating seed. It has been shown that some of the most basal angiosperms actually form diploid endosperm, although they still experience double fertilization.
The three principal types of endosperm formation found in angiosperms—nuclear, cellular, and helobial—are classified on the basis of when the cell wall forms. In nuclear endosperm formation, repeated free-nuclear divisions take place; if a cell wall is formed, it will form after free-nuclear division. In cellular endosperm formation, cell-wall formation is coincident with nuclear divisions. In helobial endosperm formation, a cell wall is laid down between the first two nuclei, after which one half develops endosperm along the cellular pattern and the other half along the nuclear pattern. Helobial endosperm is most commonly found in the Alismatales (monocotyledons). In many plants, however, the endosperm degenerates, and food is stored by the embryo (e.g., peanut 【groundnut】, Arachis hypogaea), the remaining nucellus (known as perisperm; e.g., beet), or even the seed coat (mature integuments). Cellular endosperm is the least specialized type of endosperm with nuclear and helobial types derived from it.
The zygote undergoes a series of mitotic divisions to form a multicellular, undifferentiated embryo. At the micropylar end there develops a basal stalk or suspensor, which disappears after a very short time and has no obvious function in angiosperms. At the chalazal end (the region opposite the micropyle) is the embryo proper. Differentiation of the embryo—e.g., the development of cells and organs with specific functions—involves the development of a primary root apical meristem (or radicle) adjacent to the suspensor from which the root will develop and the development of one cotyledon (in monocotyledons) or two cotyledons (in dicotyledons) at the opposite end from the suspensor. A shoot apical meristem then differentiates between the two cotyledons or next to the single cotyledon and is the site of stem differentiation.
The mature embryo is a miniature plant consisting of a short axis with one or two attached cotyledons. An epicotyl, which extends above the cotyledon(s), is composed of the shoot apex and leaf primordia; a hypocotyl, which is the transition zone between the shoot and root; and the radicle. Angiosperm seed development spans three distinct generations, plus a new entity: the parent sporophyte, the gametophyte, the new sporophyte, and the new innovation—namely, the endosperm.
Seedlings
Mature seeds of most angiosperms pass through a dormant period before eventually developing into a plant. The life span of angiosperm seeds varies from just a few days (e.g., sugar maple, Acer saccharum) to over a thousand years (e.g., sacred lotus, Nelumbo nucifera). Successful germination requires the right conditions of light, water, and temperature and usually begins with imbibition of water and the subsequent release from dormancy. During its early growth stages and before it has become totally independent of the food stored in the seed (seed and fruit) or cotyledons, the new plant is called a seedling.
Two patterns of seed germination occur in angiosperms, depending on whether the cotyledons emerge from the seed: hypogeal and epigeal. In hypogeous germination, the hypocotyl remains short and the cotyledons do not emerge from the seed but rather force the radicle and epicotyl axis to elongate out of the seed coat. The seed, with the enclosed cotyledons, remains underground, and the epicotyl grows up through the soil. When the cotyledons contain seed-storage products, these products are transferred directly to the developing radicle and epicotyl (e.g., garden pea). When the endosperm or perisperm contains the storage products, the cotyledons penetrate the storage tissues and transfer the storage products to the developing radicle and epicotyl (e.g., garlic, Allium sativum).
In epigeous germination, the radicle emerges from the seed and the hypocotyl elongates, raising the cotyledons, epicotyl, and remains of the seed coat above ground. The cotyledons may then expand and function photosynthetically as normal leaves (e.g., castor bean (castor-oil plant), Ricinus communis). When the cotyledons contain seed-storage products, they transfer them to the rest of the seedling and degenerate without becoming significantly photosynthetic (e.g., garden beans, Phaseolus). Eventually, the seedling becomes independent of the seed-storage products and grows into a mature plant capable of reproduction. Although the dispersal of seeds is essential in the reproduction and spread of angiosperm species, it is equally important for successful germination and seedling establishment to take place in an appropriate habitat.
Paleobotany and evolution
The origins and diversity of flowering plants can best be understood by studying their fossil history. The fossil record provides important data to help show when and where early angiosperms lived, why flowering plants came to exist, and from what group or groups of plants they evolved.
The earliest plants generally accepted to be angiospermous are known from the Early Cretaceous Period (about 145.5 to 99.5 million years ago). Fossil pollen of angiosperms is first found in the Hauterivian (Hauterivian Stage) and Barremian (Barremian Stage) ages, which spanned from about 136 to 125 million years ago. A very few angiosperm leaves and flowers are found in layers dating to the early Aptian Age (about 125 to 112 million years ago). Many of the earliest fossils of angiosperms are most similar to small bushes or small herbaceous plants, such as those in the Chloranthaceae (Piperales), Ceratophyllaceae, and Ranunculaceae (Ranunculales) families. More diverse flora showing a larger variety of pollen, leaves, and reproductive organs with angiospermous affinities developed during the Albian (Albian Stage) Age (about 112 to 99.5 million years ago).
From the end of the Albian (the close of the Early Cretaceous) and the beginning of the Late Cretaceous (about 99.5 to 65.5 million years ago), angiosperms further diversified and dispersed. Many woody angiosperms evolved at that time, as did several modern groups, such as the magnolia, laurel, sycamore, and rose families. Herbaceous plants such as the water lilies (Nymphaeales), the family Ceratophyllaceae, and some of the early monocotyledons also persisted from the Albian until today.
Because some of the oldest and most diverse angiosperm floras are found in Africa near the Equator, followed by low-latitude, angiosperm-dominated floras in North America, angiosperms are thought to have radiated from the Equator and spread to either pole. The angiosperms developed a close association with insect pollinators early in their evolution. This promoted outcrossing resulting in genetically vigorous offspring. Also, the relatively short generation time in which the angiosperms reproduce—permitting rapid population growth and easier colonization of disturbed habitats—gave the flowering plants an adaptive advantage over the gymnosperms, which were dominant during the Early Cretaceous. The seeds of angiosperms were small and were probably eaten and carried to new areas by animals. Thus, the angiosperms were able to migrate into and occupy new areas of the world. At the beginning of the Cenomanian (Cenomanian Stage) Age (about 99.5 to 93.5 million years ago), angiosperms probably formed dominant pockets of vegetation along many low coastal tropical and warm temperate areas of the world. During the Cenomanian the angiosperms also spread to inland continental areas as well as northward and southward along the coasts. By the Middle to Late Cenomanian (about 95 to 93.5 million years ago), angiosperms became the dominant form of vegetation in many areas of the world.
One of the most conspicuous features of angiosperms is the flower. Most frequently, flowers are brightly coloured, often scented structures containing nectar and the male and female reproductive organs. Because it is important for the genetic integrity of a plant that it avoid pollinating itself or a nearby, possibly closely related, neighbour, pollen from one plant must be moved some distance to another plant. Wind is often an effective but imprecise pollination mechanism. Frequently, flowering plants are more accurately pollinated by animals, which carry the pollen some distance to another flower. Thus, development of showy flowers has involved the coevolution of insects or other animals and the early ancestors of the angiosperms.
Various groups of extinct seed plants have been proposed as the ancestral stock at different times in the evolution of the angiosperms. The Pteridospermales (seed ferns (seed fern)) are a group of extinct early seed plants that resemble small trees and shrubs with fernlike foliage. They bore seeds on their leaves or in specialized structures derived from leaves and had specialized pollen-bearing organs or simple anthers. The ovules and pollen organs were separate reproductive units, and wind may have been the most common agent of pollen transfer. Some seed ferns of the Paleozoic Era (about 542 to 251 million years ago) contained pollen grains that were much too large to be effectively dispersed by the wind. These plants probably depended on insects to carry the pollen grains from one plant to another.
The Cycadeoidophyta are a group of extinct seed plants that contain members that have widely different reproductive structures. In some the female and male reproductive organs were separate, while in others the reproductive structures were organized into a common reproductive unit in which the male organs surrounded the female organ. These reproductive organs sat on a receptacle similar to that in flowering plants and often were surrounded by sterile bracts or leaflike tissue, which may have opened to form a flowerlike structure in the genus Williamsoniella (Cycadeoidales). Some extinct Cycadeoidales may have been pollinated by insects. The female and male reproductive organs tend to be clustered when insect pollination is involved, which is probably why most flowers are bisexual.
It is not clear whether the flowering plants are derived from the Pteridospermales or the Cycadeoidales; however, in both groups the potential existed for modification of the plant body and the reproductive tissue to be responsive to both the physical and biological environments of the Mesozoic Era (about 251 to 65.5 million years ago). The pollen evidence suggests that the Gnetales, a modern group of gymnosperms closely related to the angiosperms, were present during the Triassic Period (about 251 to 199.5 million years ago). Thus, the evolution that produced the plants which were eventually recognized as the angiosperms must have been taking place during the Triassic, Jurassic, and earliest Cretaceous periods (which span from about 251 to 99.5 million years ago).
The ancestral stock probably was a small to medium-size plant in which large leafy shoots contained individual fertile female, fertile male, and sterile leaves. The form of the plant was modified: the leaf size was reduced, and some shoots were modified so that the ovules remained enclosed inside the leaf tissue, which was shortened so that the ovule and pollen organs were borne close together. The sterile leaves may have been lost in some evolutionary lines or may have evolved into sepals and petals in others. The pollen-bearing organs (stamens) or ovule-bearing organs (carpels) may have been lost in some lines of evolution, resulting in unisexual flowers, or both may have been retained together in others to produce bisexual flowers.
Those early lines of angiosperm evolution in which wind may have functioned in pollination retained small, inconspicuous, often unisexual flowers. In those evolutionary lines that developed close associations with specific insect pollinators, the organs become dramatically modified. Small, inconspicuous bisexual or unisexual flowers are known from the Aptian Age. Large petals developed by the late Albian (about 105 million years ago). In insect-pollinated flowers and bisexual flowers that contain their characteristic nectaries, very large petals and anthers with abundant small pollen are known from the earliest Cenomanian Age. The presence of small, inconspicuous unisexual flowers, probably pollinated by wind or water, from the Aptian and late Albian suggests that the form and mode of reproduction of angiosperms were beginning to diverge from those of their ancestors even before this is attested in macrofossils.
The special features of flowering plants that enhanced the coevolutionary links with animals evolved at various times in different groups of angiosperms. There were, however, three major nodes of coevolution in the development of flowering plants: the evolution of showy flowers attractive to animal (mainly insect) pollinators, the evolution of bilaterally symmetrical flowers with variously fused parts to direct the behaviour of particular animal pollinators (especially social insects and birds), and the evolution of larger energy-rich animals (especially mammals and birds) to disperse fruits and seeds. Each of these events had a dynamic effect on the evolution of angiosperms, increasing their diversity at different times in different groups and affecting their floral and fruit morphology in various ways.
The early angiosperms appear to have had few and radially arranged flower parts. The flowers were unisexual or bisexual, with superior ovaries, loosely closed to fully closed carpels, free flower parts, and small fruits and seeds. The fossil record of the early evolution of the flower demonstrates a tendency toward an increased number of flower parts, a loose to complete fusion of carpels, the development of a style, the elevation of the stigmatic surface upon the style, a slight increase in seed size, and a diversity of ways in which flowers were borne upon the plant.
The evolution of both female and male reproductive organs in the same flower was both beneficial and problematic in the early angiosperms. Insects visiting a unisexual flower either picked up pollen or deposited pollen, depending on the sex of the flower visited. Insect visits, therefore, only randomly fertilized flowers as the insect alternated between male and female flowers. It became beneficial to the flower to evolve a place for both sexes in a single flower so that each insect visit would deposit and remove pollen. When both sexes are present in a single flower, however, there develops a strong possibility that the flower may pollinate itself, a situation that would cause inbreeding depression, thereby reducing the vigour of the offspring over successive generations. It was probably very early in the evolutionary history of flowering plants that self-incompatibility was evolved, a mechanism that prevents flowers or plants from self-pollinating. The pollen of many modern insect-pollinated bisexual flowers is incompatible with the flower in which it is produced.
Another feature of flowers that developed as a result of insect pollination is pollen tube competition. When a pollen load of 50–200 pollen grains is deposited on a stigma at one time, each pollen grain grows a pollen tube into the stigmatic tissue. The pollen tubes that grow the fastest reach the ovules first and effect fertilization. It has been demonstrated that the pollen grain with the fastest-growing pollen tube carries genes that produce more vigorous offspring. By the Early Cenomanian the stigmas of some insect-pollinated flowers were elevated on styles, effectively establishing some distance for the pollen tubes to travel. This would establish pollen tube competition as a selective mechanism within some early flowers.
During the first 70 million years of angiospermous evolution, all the known flowers were radially symmetrical. It is only in the early Tertiary (Tertiary Period) Period—specifically, during the late Paleocene (Paleocene Epoch) and early Eocene (Eocene Epoch) (about 63 to 52 million years ago)—that the first evidence of bilaterally symmetrical flowers is found. The evolution of bilateral flowers—for example, that of the legumes and orchids—is an adaptation for specialized pollinators such as social insects (bees) and some birds. The sterile organs (sepals, petals) are modified to present a certain flower orientation to the pollinator, enabling the pollinator to enter the flower where the pollen organs and pollen-receptive tissue are positioned to maximize effective pollination. During the early Tertiary the bilateral organization of floral organs coevolved with animal behaviour independently at different times and in various groups of angiosperms.
The evolution of mammals and birds also influenced the evolution of flowering plants in the early Tertiary. During the first 70–80 million years of their existence, the fruits and seeds of the angiosperms were small. The initial radiation of larger energy-rich fruits and seeds, such as the acorns, chestnuts, walnuts, legume pods, and the earliest grasses, took place during the Eocene. These fruits appeared over a short period of time contemporaneously with the diversification of seed- and fruit-eating mammals and birds. Seeds of fleshy fruits, such as grapes, also became common in the Eocene (about 45 million years ago). Thus, a second important node of plant and animal coevolution apparently developed about 50–60 million years ago, when angiosperms began to produce fruits and seeds that were attractive to animals. The animals served as agents to carry fruits and seeds some distance from the parent plant, further enhancing the potential for outcrossing and aiding in the dispersal of angiospermous plants to new areas of the world.
In summary, the evolutionary history of angiosperms is intimately but not exclusively tied to their coevolution with animal pollinators and agents of fruit and seed dispersal. Wind and water pollination and fruit and seed dispersal also continued throughout the entire evolutionary history of flowering plants. This network of evolutionary pressures resulted in the variety of flowers and fruits representative of present-day angiosperms. Accordingly, some of the most useful characters in determining the particular taxon to which living angiosperms belong are flowers, fruits, and seeds. The evolution of such vegetative characteristics as wood and leaves is more complex and less well understood.
Classification
Diagnostic classification
General characteristics
The angiosperms are a well-characterized, sharply defined group. There is not a single living plant species whose status as an angiosperm or non-angiosperm is in doubt. Even the fossil record provides no forms that connect with any other group, although there are of course some fossils of individual plant parts that cannot be effectively classified.
Most typically, angiosperms are seed plants. This separates them from all other plants except the gymnosperms (gymnosperm), of which the most familiar representatives are the conifers and cycads.
The ovules (forerunners of the seeds) of angiosperms are characteristically enclosed in an ovary, in contrast to those of gymnosperms, which are exposed to the air at the time of pollination and never enclosed in an ovary. Pollen of angiosperms is received by the stigma, a specialized structure that is usually elevated above the ovary on a more slender structure known as the style. Pollen grains germinate on the stigma, and the pollen tube must grow through the tissues of the style (if present) and the ovary to reach the ovule. The pollen grains of gymnosperms, in contrast, are received at an opening (the micropyle) atop the ovule.
The female gametophyte of angiosperms (called the embryo sac) is tiny and contains only a few (typically eight) nuclei (nucleus); the cytoplasm associated more or less directly with these nuclei is not partitioned by cell walls. One of the several nuclei of the embryo sac serves as the egg in sexual reproduction, uniting with one of the two sperm nuclei delivered by the pollen tube. Two other nuclei of the embryo sac fuse with the second sperm nucleus from the pollen tube. This triple-fusion nucleus is characteristically the forerunner of a multicellular food-storage tissue in the seed, called the endosperm.
The process in which both nuclei from the pollen tube fuse is referred to as double fertilization. This is perhaps the most characteristic single feature of angiosperms and is not shared with any other group. Gymnosperms, in sharp contrast, have a multicellular female gametophyte that consists of many hundreds or even thousands of cells. Double fertilization does not take place in this case, and the female gametophyte develops into the food-storage tissue of the seed.
Furthermore, angiosperms have a more complex set of conducting tissues than do gymnosperms. The water-conducting tissue (xylem) ordinarily includes some long tubes called vessels. Only one small group of gymnosperms, the Gnetophyta (gnetophyte), has vessels. The food-conducting tissue (phloem) of angiosperms characteristically has companion cells that bear a direct ontogenetic relationship to the sieve tubes through which the actual conduction takes place. The phloem of gymnosperms has less-specialized sieve cells and lacks companion cells.
Eudicots
One of the major changes in the understanding of the evolution of the angiosperms was the realization that the basic distinction among flowering plants is not between monocotyledon groups (monocots) and dicotyledon groups (dicots). Rather, plants thought of as being “typical dicots” have evolved from within another group that includes the more-basal dicots and the monocots together. This group of typical dicots is now known as the eudicots. Molecular-based evidence supports their being a single evolutionary lineage (monophyletic), and they are characterized by pollen that fundamentally has three furrows or pores (tricolpate), in contrast to the single pore or furrow of the monocot and basal dicot group (monosulcates).
Within the eudicots there is a large clade called the core eudicots, nearly all members of which show major differences in floral morphology from that of other flowering plants. In particular, the basic construction of the flower is much more stereotyped than in the basal eudicots, monocots, and basal dicots. Within nearly every order of the core eudicots, there are families with a basic “5 + 5 + (5 or 10) + (3 or 5)” floral construction. This refers to five sepals, five petals, one whorl of five stamens, often another whorl of five stamens, and finally a whorl of three or five carpels. The members of the whorls alternate with each other so that the petals are on radii midway between the sepal radii; the carpels in the centre of the flower are on the same radii as the sepals but are opposite to them. When core eudicots have only five stamens, as is common, these stamens usually are the stamens of the outer whorl—that is, they alternate with the petals and are opposite the sepals. Furthermore, the carpels at least are more or less fused, and there is often a well-developed nectary disc either surrounding the base of the ovary or, less frequently, borne immediately outside the stamens. The flowers are usually perfect and are radially symmetric. It is interesting that some families in most of the core eudicot orders, including Asterid orders such as Cornales and Ericales, have members with many stamens, but in nearly all cases these stamens develop in a different way than the numerous stamens in families such as Ranunculaceae (a basal eudicot) or Magnoliaceae (a basal dicot).
Core eudicots commonly show other features as well. Instead of the stamens having the pollen sac, or anther, attached at the base to a stalk, or filament, the two being more or less continuous, in core eudicots the filament is often attached at the back of the anther, and it narrows considerably just before it joins the anther. The pollen of the core eudicots commonly has three longitudinal depressions, or colpi, as does the pollen of the rest of the eudicots, but in the middle of each colpus there is a circular pore through which the pollen tube emerges; that is, the pollen is basically tricolporate. There are many deviations from this generalized structure, but keeping it in mind as a reference is helpful for understanding angiosperm evolution.
Within the core eudicots there are a number of major clades. These include the Asterids and Rosids, which are very species-rich, the former particularly so. The basic arrangement of the flower parts in these eudicot clades does not change, but the petals are commonly fused in the Asterids, forming a corolla tube. There are also chemicals common in the Asterids that are very rare in any other flowering plants.
Annotated classification
The classification of flowering plants used here is a significant departure from the botanical classification system of the American botanist Arthur Cronquist (1981), which was based on similarities and differences in morphological, chemical, and anatomical characters. Since the early 1990s, studies of plant phylogeny have been transformed by newly available molecular techniques, mainly involving sequencing of segments of DNA from the chloroplasts and the nuclei of plant cells, as well as improved computer programs to analyze large amounts of data. These techniques, which provided more robust and testable data on plant phylogeny, often conflicted with older, morphological-based schemes such as the Cronquist system. In 1998 a group of scientists who were participating in large-scale molecular analysis of flowering plants proposed a new overall classification system for the angiosperms. They called themselves the Angiosperm Phylogeny Group, and their new scheme became known as the APG system.
The APG system focused mainly on the level of families (with related families grouped into orders) because they are the groups around which most botanists organize their understanding of plant diversity. It need not be assumed, however, that different families or orders are equivalent in any evolutionary sense; rather, the APG organization signals a relative level in a hierarchy. Within any particular family, though, the system does presume, with some possible exceptions, that the genera included in it are all related and that the family itself is monophyletic (a lineage with all its members derived from a common ancestor); the same holds for the families included within a particular order. One of the main departures from the Cronquist system in the APG system is a less hierarchical arrangement of the higher-level groupings, which Cronquist divided into two classes: the monocotyledons (monocotyledon) (monocots), or Liliopsida, with five subclasses, and the dicotyledons (dicotyledon) (dicots), or Magnoliopsida, with six subclasses. The APG system does recognize some higher-level groupings but only at an informal level, such as eudicots, Rosids, and Asterids. It continues to recognize the monocots as a monophyletic group; however, they are now seen as having evolved from within a more-basal group of primitive dicotyledonous angiosperms. In contrast, Cronquist portrayed the monocots as being the sister group to all other dicotyledonous groups.
The APG system was not intended to be definitive, since some families were not included in the first large molecular analyses, and some of the relationships suggested were fairly tentative. Following the original APG publication, more families were added to the molecular analyses, allowing these families to be placed in orders, and other new studies called for adjustments in the circumscription of particular families and orders. These changes were incorporated into an update in 2003 of the APG known as APG II, and the synopsis of flowering-plant classification presented here follows the APG II system. The number of recognized orders increased from 40 in the original APG system to 62 in APG II, depending on whether some single-family orders are recognized. It is important to recognize that modifications to the APG II system continue as new data become available.
Basalmost angiosperms (angiosperm)
The first three groups listed below are those that appear at the base of the angiosperm tree, although the relationships among them are still somewhat unclear. Claims of having identified the “most basal” living angiosperm have been put forth and emended repeatedly, but DNA evidence argues for Amborellaceae and Nymphaeaceae as the basalmost offshoots of the flowering plants. In APG II both families are ascribed to their own orders. The interesting feature about the basalmost groups is that they form a sequentially branching comb or “grade” rather than a more regular bifurcating pattern of distinct clades of monophyletic groups.
Order Amborellales
Family: Amborellaceae (a single genus and species, Amborella trichopoda, which is native to New Caledonia).
Order Austrobaileyales
Families: Austrobaileyaceae, Illiciaceae, Trimeniaceae.
Order Nymphaeales
Families: Nymphaeaceae, Cabombaceae, Hydatellaceae.
Magnoliid
A group of 5 orders of basal angiosperms.
Order Canellales
Families: Canellaceae, Winteraceae.
Order Chloranthales
Family: Chloranthaceae.
Order Laurales
Families: Atherospermataceae, Calycanthaceae, Gomortegaceae, Hernandiaceae, Lauraceae, Monimiaceae, Siparunaceae.
Order Magnoliales
Families: Annonaceae, Degeneriaceae, Eupomatiaceae, Himantandraceae, Magnoliaceae, Myristicaceae.
Order Piperales
Families: Aristolochiaceae, Hydnoraceae, Piperaceae, Saururaceae.
Monocotyledons
This large group of orders is an important angiosperm lineage long recognized for its essentially herbaceous members, a single cotyledon in the seedlings, vascular bundles scattered in a cross section of the stem, leaves not differentiated into a separate petiole and blade, venation usually parallel and converging toward the leaf apex, and flowers mostly in multiples of 3 parts.
Order Acorales
Family: Acoraceae (the basalmost branch of the monocots, with a single genus, Acorus 【sweet flag】).
Order Alismatales
Families: Alismataceae, Aponogetonaceae, Araceae, Butomaceae, Cymodoceaceae, Hydrocharitaceae, Juncaginaceae, Limnocharitaceae, Posidoniaceae, Potamogetonaceae, Ruppiaceae, Scheuchzeriaceae, Tofieldiaceae, Zosteraceae.
Order Asparagales
Families: Agapanthaceae, Agavaceae, Alliaceae, Amaryllidaceae, Aphyllanthaceae, Asparagaceae, Asphodelaceae, Asteliaceae, Blandfordiaceae, Boryaceae, Doryanthaceae, Hemerocallidaceae, Hyacintheaceae, Hypoxidaceae, Iridaceae, Ixioliriaceae, Lanariaceae, Laxmanniaceae, Orchidaceae, Ruscaceae, Tecophilaeaceae, Themidaceae, Xanthorrhoeaceae, Xeronemaceae.
Order Dioscoreales
Families: Burmanniaceae, Dioscoreaceae (including Trichopodaceae), Nartheciaceae, Taccaceae, Thismiaceae.
Order Liliales
Families: Alstroemeriaceae, Campynemataceae, Colchicaceae, Corsiaceae, Liliaceae, Luzuriagaceae, Melanthiaceae,Petermanniaceae, Philesiaceae, Rhipogonaceae, Smilacaceae.
Order Pandanales
Families: Cyclanthaceae, Pandanaceae, Stemonaceae, Triuridaceae, Velloziaceae.
Order Petrosaviales
Family: Petrosaviaceae.
Commelinids
An assemblage of 4 related monocot orders, with Dasypogonaceae unplaced among them.
Order Arecales
Family: Arecaceae.
Order Commelinales
Families: Commelinaceae, Haemodoraceae, Hanguanaceae, Philydraceae, Pontederiaceae.
Order Poales
Families: Anarthriaceae, Bromeliaceae, Centrolepidaceae, Cyperaceae, Ecdeiocoleaceae, Eriocaulaceae, Flagellariaceae, Joinvilleaceae, Juncaceae, Mayacaceae, Poaceae, Rapateaceae, Restionaceae, Thurniaceae, Typhaceae (including Sparganiaceae), Xyridaceae.
Order Zingiberales
Families: Cannaceae, Costaceae, Heliconiaceae, Lowiaceae, Marantaceae, Musaceae, Strelitziaceae, Zingiberaceae.
Eudicots
All the remaining dicotyledonous groups, with mainly 3-aperturate pollen and lacking the ethereal oils found in many of the basalmost angiosperm groups.
Basal eudicots
Order Buxales
Families: Buxaceae, Didymelaceae.
Order Ceratophyllales
Family: Ceratophyllaceae (an aquatic group once thought to be the basalmost angiosperm group).
Order Gunnerales
Families: Gunneraceae, Myrothamnaceae.
Order Proteales
Families: Nelumbonaceae, Platanaceae, Proteaceae.
Order Ranunculales
Families: Berberidaceae, Circaeasteraceae, Eupteleaceae, Lardizabalaceae, Menispermaceae, Papaveraceae (including Fumariaceae and Pteridophyllaceae), Ranunculaceae.
Order Sabiales
Family: Sabiaceae.
Order Trochodendrales
Family: Trochodendraceae (including Tetracentraceae).
Core eudicots
For the most part, the basic construction of the flower in the core eudicots is much more stereotyped than in the basal eudicots, monocots, or basal dicots. Within nearly every order of core eudicots, there are families with a basic floral pattern of 5 sepals, 5 petals, 5 or 10 stamens, and 3 or 5 carpels (with many exceptions). The members of the different whorls of the flower typically alternate with each other, the carpels are generally fused, and there is often a nectary disc surrounding the base of the ovary or (less often) outside the stamens. The flowers are often bisexual and radially symmetric, although there is much zygomorphy or biradial symmetry in the flowers of this group as well.
Basal core eudicots
Order Berberidopsidales
Families: Aextoxicaceae, Berberidopsidaceae.
Order Caryophyllales
Families: Achatocarpaceae, Aizoaceae, Amaranthaceae, Ancistrocladaceae, Asteropeiaceae, Barbeiuaceae, Basellaceae, Cactaceae, Caryophyllaceae, Didieraceae, Dioncophylleaceae, Droseraceae, Drosophyllaceae, Frankeniaceae, Giseckiaceae, Halophytaceae, Limeaceae, Lophiocarpaceae, Molluginaceae, Montiaceae, Nepenthaceae, Nyctaginaceae, Physenaceae, Phytolaccaceae, Plumbaginaceae, Polygonaceae, Portulacaceae, Rhabdodendraceae, Sarcobataceae, Simmondsiaceae, Stegnospermataceae, Talinaceae, Tamaricaceae.
Order Dilleniales
Family: Dilleniaceae.
Order Santalales
Families: Balanophoraceae, Erythropalaceae, Loranthaceae, Misodendraceae, Olacaceae, Opiliaceae, Santalaceae, Schoepfiaceae.
Unplaced eudicots
The position of the following 2 orders is not fully resolved.
Order Saxifragales
Families: Altingiaceae, Aphanopetalaceae, Cercidiphyllaceae, Crassulaceae, Daphniphyllaceae, Grossulariaceae, Haloragaceae, Hamamelidaceae, Iteaceae, Paeoniaceae, Penthoraceae, Pterostemonaceae, Saxifragaceae, Tetracarpaeaceae.
Order Vitales
Family: Vitaceae.
Rosids
A group that can be divided into several distinct lineages, which APG II identifies as the basal Rosids, Rosids I, and Rosids II.
Basal Rosids
The following 3 orders and the unplaced family Picramniaceae.
Order Crossosomatales
Families: Aphloiaceae, Crossosomataceae, Geissolomataceae, Guamatelaceae, Ixerbaceae, Stachyuraceae, Staphyleaceae, Strasburgeriaceae.
Order Geraniales
Families: Francoaceae, Geraniaceae, Ledocarpaceae, Melianthaceae, Vivianaceae.
Order Myrtales
Families: Alzateaceae, Combretaceae, Crypteroniaceae, Lythraceae, Melastomataceae, Memecylaceae, Myrtaceae, Oliniaceae, Onagraceae, Penaeaceae, Rhynchocalycaceae, Vochysiaceae.
Rosid I clade
The following 8 orders.
Order Celastrales
Families: Celastraceae, Lepidobotryaceae, Parnassiaceae, Pottingeriaceae.
Order Cucurbitales
Families: Anisophylleaceae, Begoniaceae, Coriariaceae, Corynocarpaceae, Cucurbitaceae, Datiscaceae, Tetramelaceae.
Order Fabales
Families: Fabaceae, Polygalaceae, Quillajaceae, Surianaceae.
Order Fagales
Families: Betulaceae, Casuarinaceae, Fagaceae, Juglandaceae, Myricaceae, Nothofagaceae, Rhoipteleaceae, Ticodendraceae.
Order Malpighiales
Families: Achariaceae (includes part of former Flacourtiaceae), Balanopaceae, Bonnetiaceae, Caryocaraceae, Centroplacaceae, Chrysobalanaceae, Clusiaceae, Ctenolophonaceae, Dichapetalaceae, Elatinaceae, Erythroxylaceae, Euphorbiaceae, Euphroniaceae, Goupiaceae, Humiriaceae, Hyperiaceae, Irvingiaceae, Ixonanthaceae, Lacistemataceae, Linaceae, Lophopyxidaceae, Malesherbiaceae, Malpighiaceae, Medusagynaceae, Ochnaceae, Pandaceae, Passifloraceae, Peraceae, Phyllanthaceae, Picrodendraceae, Podostemaceae, Putranjivaceae, Quiinaceae, Rafflesiaceae, Rhizophoraceae, Salicaceae (includes another part of former Flacourtiaceae), Trigoniaceae, Turneraceae, Violaceae.
Order Oxalidales
Families: Brunelliaceae, Cephalotaceae, Connaraceae, Cunoniaceae, Elaeocarpaceae, Huaceae, Oxalidaceae.
Order Rosales
Families: Barbeyaceae, Cannabaceae, Dirachmaceae, Elaeagnaceae, Moraceae, Rhamnaceae, Rosaceae, Ulmaceae, Urticaceae.
Order Zygophyllales
Families: Krameriaceae, Zygophyllaceae.
Rosid II clade
The following 4 orders.
Order Brassicales
Families: Akaniaceae, Bataceae, Brassicaceae, Capparaceae, Caricaceae, Cleomaceae, Emblingiaceae, Gyrostemonaceae, Koeberliniaceae, Limnanthaceae, Moringaceae, Pentadiplandraceae, Resedaceae, Salvadoraceae, Setchellanthaceae, Stixaceae, Tovariaceae, Tropaeolaceae.
Order Huerteales
Families: Dipentodontaceae (including Perrottetia), Gerrardinaceae, Tapisciaceae.
Order Malvales
Families: Bixaceae, Cistaceae, Cytinaceae, Dipterocarpaceae, Malvaceae (including Bombacaceae, Sterculiaceae, and Tiliaceae), Muntingiaceae, Neuradaceae, Sarcolaenaceae, Sphaerosepalaceae, Thymeleaceae.
Order Sapindales
Families: Anacardiaceae, Biebersteiniaceae, Burseraceae, Kirkiaceae, Meliaceae, Nitrariaceae, Rutaceae, Sapindaceae (including Aceraceae and Hippocastanaceae), Simaroubaceae.
Asterids
A strongly supported group of about 10 orders, most of them with a corolla tube and few stamens.
Basal Asterids
Order Cornales
Families: Cornaceae, Curtisiaceae, Grubbiaceae, Hydrangeaceae, Hydrostachyaceae, Loasaceae, Nyssaceae.
Order Ericales
Families: Actinidaceae, Balsaminaceae, Clethraceae, Cyrillaceae, Diapensiaceae, Ebenaceae, Ericaceae, Fouquieriaceae, Lecythidaceae, Maesaceae, Marcgraviaceae, Mitrastemonaceae, Myrsinaceae, Pentaphylacaceae, Polemoniaceae, Primulaceae, Roridulaceae, Sapotaceae, Sarraceniaceae, Sladeniaceae, Styracaceae, Symplocaceae, Tetrameristaceae, Theaceae, Theophrastaceae.
Asterid I clade
The following 4 orders and several unplaced families: Boraginaceae, Icacinaceae, Oncothecaceae, and Vahliaceae.
Order Garryales
Families: Eucommiaceae, Garryaceae.
Order Gentianales
Families: Apocynaceae, Gelsemiaceae, Gentianaceae, Loganiaceae, Rubiaceae.
Order Lamiales
Families: Acanthaceae, Bignoniaceae, Byblidaceae, Calceolariaceae, Carlemanniaceae, Gesneriaceae, Lamiaceae, Lentibulariaceae, Linderniaceae, Martyniaceae, Oleaceae, Orobanchaceae, Paulowniaceae, Pedaliaceae, Phrymaceae, Plantaginaceae, Plocospermataceae, Schlegeliaceae, Scrophulariaceae (including Buddlejaceae), Stilbaceae, Tetrachondraceae, Thomandersiaceae, Verbenaceae.
Order Solanales
Families: Convolvulaceae, Hydroleaceae, Montiniaceae, Solanaceae, Sphenocleaceae.
Asterid II clade
The following 4 orders and a number of unplaced families: Escalloniaceae (including Eremosynaceae), Paracryphiaceae, Polyosmaceae, Quintiniaceae, Sphenostemonaceae, Tribelaceae.
Order Apiales
Families: Apiaceae, Araliaceae, Griseliniaceae, Myodocarpaceae, Pennantiaceae, Pittosporaceae, Torricelliaceae.
Order Aquifoliales
Families: Aquifoliaceae, Cardiopteridaceae, Helwingiaceae, Phyllonomaceae, Stemonuraceae.
Order Asterales
Families: Alseuosmiaceae, Argophyllaceae, Asteraceae, Calyceraceae, Campanulaceae (includes Lobeliaceae), Goodeniaceae, Menyanthaceae, Pentaphragmataceae, Phellinaceae, Rousseaceae, Stylidiaceae.
Order Dipsacales
Families: Adoxaceae, Caprifoliaceae, Diervillaceae, Dipsacaceae, Linnaeaceae, Morinaceae, Valerianaceae.
Additional Reading
General works
Various aspects of plant life, including angiosperms, are dicussed in Peter H. Raven, Ray F. Evert, and Susan E. Eichhorn, Biology of Plants, 7th ed. (2005), an excellent general treatment; and D.J. Mabberley, Mabberley's Plant-Book, 3rd ed. (2008), an excellent source for specific information on genera and families, arranged alphabetically with common-name entries. Descriptive, illustrated listings of plants may be found in Kenneth A. Beckett et al., The RHS Encyclopedia of House Plants (1995); Jacqueline Hériteau et al., The National Arboretum Book of Outstanding Garden Plants (1990); and Pierre Anglade (ed.), Larousse Gardening and Gardens (1990).Economically important plants of all families are addressed in John C. Roecklein and Pingsun Leung, A Profile of Economic Plants (1987); F.N. Howes, A Dictionary of Useful and Everyday Plants and Their Common Names (1974); Charles B. Heiser, Jr., Of Plants and People (1985), and Seed to Civilization: The Story of Food, new ed. (1990); Beryl Brintnall Simpson and Molly Conner-Ogorzaly, Economic Botany: Plants in Our World, 3rd ed. (2001); N.T. Gill and K.C. Vear, Agricultural Botany, vol. 1, Dicotyledonous Crops, and vol. 2, Monocotyledonous Crops, 3rd ed. rev. (1980); Mas Yamaguchi, World Vegetables (1983); Martin Chudnoff, Tropical Timbers of the World (1984); Douglas Patterson, Commercial Timbers of the World, 5th ed. (1988); Steven Nagy and Philip E. Shaw, Tropical and Subtropical Fruits: Composition, Properties, and Uses (1980); J.A. Samson, Tropical Fruits, 2nd ed. (1986); Lawrence K. Opeke, Tropical Tree Crops (1982); Frederic Rosengarten, Jr., The Book of Edible Nuts (1984), and The Book of Spices, rev. ed. (1973, reissued 1981); J.W. Purseglove et al., Spices, 2 vol. (1981); and Gerhard Röbbelen, R. Keith Downey, and Amram Ashri (eds.), Oil Crops of the World (1989).Poisonous and medicinally useful plants are described in Walter H. Lewis and Memory P.F. Elvin-Lewis, Medical Botany: Plants Affecting Man's Health, 2nd ed. (2003); and Lewis S. Nelson, Handbook of Poisonous and Injurious Plants, 2nd ed. (2007).Insectivorous varieties are treated in Adrian Slack, Carnivorous Plants, rev. ed. (1988); James Pietropaolo and Patricia Pietropaolo, Carnivorous Plants of the World (1986); and B.E. Juniper, R.J. Robins, and D.M. Joel, The Carnivorous Plants (1989).
General features
V.H. Heywood (ed.), Flowering Plants of the World (1978, reissued 1985), is an illustrated guide to flowering plant families with distribution maps and uses. Flowering plants from several families are discussed together in Peter Bernhardt, Wily Violets & Underground Orchids (1989), a popular treatment of forest cyclicality, North American grassland pollination, Australian mistletoe, Amazonian giant water lilies, violets, and orchids. C.D. Sculthorpe, The Biology of Aquatic Vascular Plants (1967, reprinted 1985), is a comprehensive monograph on water plants.
Structure and function
Discussions of angiosperm anatomy are included in Arthur J. Eames, Morphology of the Angiosperms (1961, reprinted 1977), emphasizing floral structure and diversity; Katherine Esau, Anatomy of Seed Plants, 2nd ed. (1977), emphasizing developmental vegetative anatomy and growth; Ernest M. Gifford and Adriance S. Foster, Morphology and Evolution of Vascular Plants, 3rd ed. (1989), a succinct comparative treatment of reproductive and vegetative anatomy, morphology, and evolution; A. Fahn, Plant Anatomy, 4th ed. (1990), covering all aspects of reproductive and vegetative plant anatomy; F. Weberling, Morphology of Flowers and Inflorescences (1989; originally published in German, 1981), a thorough treatment of flower and inflorescence structure, development, and diversity; Ghillean Tolmie Prance and Kjell B. Sandved, Leaves (1985); and Francis E. Putz and Harold A. Mooney (eds.), The Biology of Vines (1992).
Reproduction
K. Faegri and L. van der Pijl, The Principles of Pollination Ecology, 3rd rev. ed. (1979), provides a thorough treatment of pollination biology and plant-animal interactions. B.M. Johri (ed.), Embryology of Angiosperms (1984), summarizes recent advances in the knowledge of flowering plant reproduction.
Paleobotany and evolution
The fossil record is analyzed in a number of books, including Wilson N. Stewart, Paleobotany and the Evolution of Plants, 2nd rev. ed. (1992); and Else Marie Friis, William G. Chaloner, and Peter R. Crane (eds.), The Origins of Angiosperms and Their Biological Consequences (1987).
- Winthrop Rockefeller
- Winthrop Sargeant
- Winton
- Winton, Alexander
- Wintun
- wire
- Wiremu Kingi
- wireworm
- Wirral
- Wirth, Joseph
- Wirth, Louis
- Wirt, William
- Wisbech
- Wisconsin
- Wisconsin Dells
- Wisconsin Evangelical Lutheran Synod
- Wisconsin, flag of
- Wisconsin Glacial Stage
- Wisconsin River
- Wisconsin, University of
- Wisdom of Solomon
- Wise, Isaac Mayer
- Wise, John
- Wiseman, Frederick
- Wiseman, Nicholas