DNA损伤修复(卷名:生物学)
repair of DNA damage
在多种酶的作用下,生物细胞内的DNA分子受到损伤以后恢复结构的现象。 DNA损伤修复的研究有助于了解基因突变机制,衰老和癌变的原因,还可应用于环境致癌因子的检测。
简史 1949年A.凯尔纳偶然发现灰色链丝菌等微生物经紫外线(UV)照射后如果立即暴露在可见光下则可减少死亡。此后在大量的微生物实验中都发现了这种现象,并证明这是许多种微生物固有的DNA损伤修复功能,并把这一修复功能称为光复活。1958年R.L.希尔证明即使不经可见光的照射,大肠杆菌也能修复它的由紫外线所造成的DNA损伤,而后又证明其他微生物也有这种功能,当时就把这种修复功能称为暗复活或暗修复。此后发现暗修复普遍地存在于原核生物、低等真核生物、高等真核生物的两栖类乃至哺乳动物中,并证实暗修复包括切除修复和复制后修复两种。1968年美国学者J.E.克利弗首先发现人类中的常染色体隐性遗传的光化癌变疾病──着色性干皮病(XP)是由基因突变造成的 DNA损伤切除修复功能的缺陷引起的。这一发现为恶性肿瘤的发生机理提供了一个重要的分子生物学证据,也使DNA损伤修复的研究进入了医学领域。
损伤类型 DNA分子的损伤类型有多种。UV照射后DNA分子上的两个相邻的胸腺嘧啶 (T)或胞嘧啶(C)之间可以共价键连结形成环丁酰环,这种环式结构称为二聚体。胸腺嘧啶二聚体的形成是 UV对DNA分子的主要损伤方式(图1)。

Χ射线、γ射线照射细胞后,由细胞内的水所产生的自由基既可使DNA分子双链间氢键断裂,也可使它的单链或双链断裂。化学物中的博莱霉素、甲基磺酸甲烷等烷化剂也能造成链的断裂。
丝裂霉素C可造成DNA分子单链间的交联,这种情况常发生在两个单链的对角的鸟嘌呤之间。链的交联也往往带来DNA分子的断裂。
DNA 分子还可以发生个别碱基或核苷酸的变化。例如碱基结构类似物5-溴尿嘧啶等可以取代个别碱基,亚硝酸能引起碱基的氧化脱氨反应,原黄素(普鲁黄)等吖啶类染料和甲基氨基偶氮苯等芳香胺致癌物可以造成个别核苷酸对的增加或减少而引起移码突变(见基因突变)。
一种 DNA损伤剂往往可以同时引起几种类型的损伤,其损伤效应的大小和类型与剂量及细胞所处的周期状态有关。
修复方式 光复活 又称光逆转。这是在可见光(波长3000~6000埃)照射下由光复活酶识别并作用于二聚体,利用光所提供的能量使环丁酰环打开而完成的修复过程 (图2)。光复活酶已在细菌、酵母菌、原生动物、藻类、蛙、鸟类、哺乳动物中的有袋类和高等哺乳类及人类的淋巴细胞和皮肤成纤维细胞中发现。这种修复功能虽然普遍存在,但主要是低等生物的一种修复方式,随着生物的进化,它所起的作用也随之削弱。

切除修复 又称切补修复。最初在大肠杆菌中发现,包括一系列复杂的酶促DNA修补复制过程,主要有以下几个阶段:核酸内切酶识别DNA损伤部位,并在5'端作一切口,再在外切酶的作用下从5'端到3'端方向切除损伤;然后在 DNA多聚酶的作用下以损伤处相对应的互补链为模板合成新的 DNA单链片断以填补切除后留下的空隙;最后再在连接酶的作用下将新合成的单链片断与原有的单链以磷酸二酯链相接而完成修复过程(图3)。

切除修复并不限于修复嘧啶二聚体,也可以修复化学物等引起的其他类型的损伤。从切除的对象来看,切除修复又可以分为碱基切除修复和核苷酸切除修复两类。碱基切除修复是先由糖基酶识别和去除损伤的碱基,在DNA单链上形成无嘌呤或无嘧啶的空位,这种空缺的碱基位置可以通过两个途径来填补:一是在插入酶的作用下以正确的碱基插入到空缺的位置上;二是在核酸内切酶的催化下在空位的5'端切开DNA链,从而触发上述一系列切除修复过程。对于各种不同类型的碱基损伤都有特异的糖基酶加以识别。不同的核酸内切酶对于不同类型损伤的识别也具有相对的特异性。
切除修复功能广泛存在于原核生物和真核生物中,也是人类的主要修复方式,啮齿动物 (如仓鼠、小鼠)先天缺乏切除修复的功能。
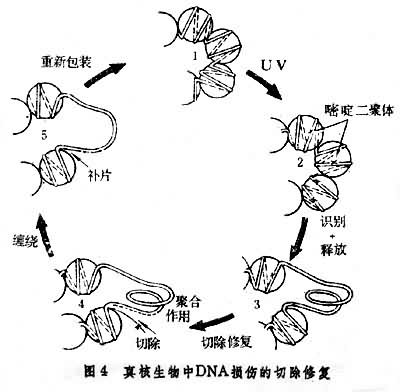
1978 年美国学者 J.L.马克斯发现真核生物与原核生物间由于染色质结构不同, 切除修复的过程也不相同。真核生物的DNA分子不象原核生物那样是裸露的,而是缠绕在组蛋白上形成串珠状的核小体结构。真核生物中的嘧啶二聚体的切除分两个阶段:快速切除期,约需2~3小时,主要切除未与组蛋白结合的DNA部分的损伤;缓慢切除期,至少要持续35小时而且需要有某种控制因子去识别这种损伤,使DNA受损部分从核小体中暴露出来,然后经过一系列步骤完成切除修复,然后修复的DNA分子再缠绕在组蛋白上重新形成核小体(图4)。
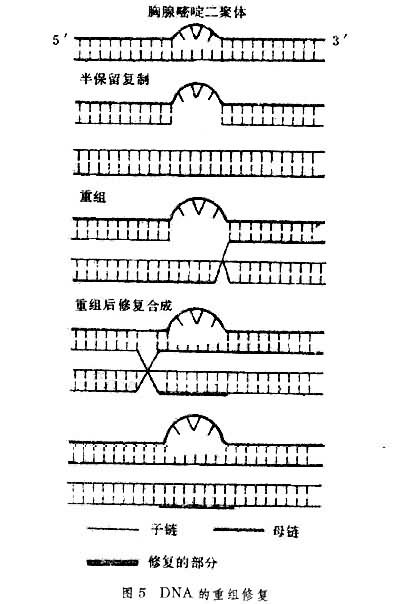
重组修复 重组修复从 DNA分子的半保留复制开始,在嘧啶二聚体相对应的位置上因复制不能正常进行而出现空缺,在大肠杆菌中已经证实这一DNA损伤诱导产生了重组蛋白,在重组蛋白的作用下母链和子链发生重组,重组后原来母链中的缺口可以通过DNA多聚酶的作用,以对侧子链为模板合成单链DNA片断来填补,最后也同样地在连接酶的作用下以磷酸二脂键连接新旧链而完成修复过程(图7)。重组修复也是啮齿动物主要的修复方式。重组修复与切除修复的最大区别在于前者不须立即从亲代的DNA分子中去除受损伤的部分,却能保证DNA复制继续进行。原母链中遗留的损伤部分,可以在下一个细胞周期中再以切除修复方式去完成修复。
SOS修复 是SOS反应的一种功能。SOS反应是DNA受到损伤或脱氧核糖核酸的复制受阻时的一种诱导反应。在大肠杆菌中,这种反应由recA-lexA系统调控。正常情况下处于不活动状态。当有诱导信号如 DNA损伤或复制受阻形成暴露的单链时,recA蛋白的蛋白酶活力就会被激活,分解阻遏物lexA蛋白,使SOS反应有关的基因去阻遏而先后开放,产生一系列细胞效应。引起SOS反应的信号消除后,recA蛋白的蛋白酶活力丧失,lexA蛋白又重新发挥阻遏作用(图6)。

SOS 反应发生时, 可造成损伤修复功能的增强。如uvrA、uvrB、uvrC、uvrD、ssb、recA、recN和ruv基因发达从而增强切除修复、复制后修复和链断裂修复。而recA和umuD.C则参与一种机制不清的易错修复,使细胞存活率增加,突变率也增加。
除修复作用外,SOS反应还可造成细胞分裂受阻、溶原性噬菌体释放和DNA复制形式的改变。后者指DNA聚合酶I*的形成,使DNA复制的准确性降低并可通过损伤部位。此时,DNA复制的起始也无需新合成蛋白。
在真核细胞中,虽然还不清楚具体过程,但肯定存在可诱导的易错修复。酵母RAD6系统就是一种易错修复系统。在哺乳类细胞中,DNA损伤可诱导细胞内病毒的释放、病毒转化作用的加强、染色体重组增强和细胞纤溶酶激活物的形成等。并且还发现了和大肠杆菌相似的ω-复活效应和ω-诱变效应。由于这种反应可增强突变、染色体重排和病毒的活动,以及对 DNA复制形式的影响,可能与癌基因激活和肿瘤形成有直接的关系。因而,SOS反应可作为检测药物致癌性的指标,而抑制SOS反应的药物则可减少突变和癌变。这类物质被称之为抗变剂。
适应性修复 1977年美国学者L.萨姆森等在大肠杆菌中发现的不同于 SOS修复的又一种诱导反应,它可以修复鸟嘌呤碱基的甲基化。如先以每毫升培养基 1微克的诱变剂N-甲基-N'-硝基-亚硝基胍 (MNNG)培养大肠杆菌两小时,就能使大肠杆菌对MNNG浓度高几百倍的环境产生抗性。这是由于 MNNG引起的DNA链上的鸟嘌呤甲基化诱导合成甲基受体蛋白,这种甲基受体蛋白分子的半胱氨酸能和甲基基团结合形成S-甲基半胱氨酸,从而使甲基化的鸟嘌呤碱基得以修复。
链断裂修复 包括 DNA分子的单链断裂修复、双链断裂修复和染色体的断裂重接修复。在连接酶的参与下这些断裂能够迅速地以重接的方式修复。这种修复有两个特点:一是不稳定性,重接后又可以再度离解;二是不正确性,经常发生随机的重接错误。
链交联修复 起始步骤是在糖基酶的催化下解开交联的一条臂, 通过碱基切除的方式先修复合成其中一条单链,然后再在内切酶的催化下,以核苷酸切除修复的方式从相反的方向修复对侧的单链片断。
检测方法 大部分DNA损伤修复都依赖于DNA的修复合成,所以对修复合成的测定常用来作为DNA修复的检测方法。常用的有以下几种:
放射自显影法 在细胞培养物中加入氚标记的胸腺嘧啶核苷
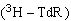 等放射源,用放射自显影方法计数银颗粒数来测定修复合成过程中
等放射源,用放射自显影方法计数银颗粒数来测定修复合成过程中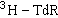 参入到DNA分子中的量。
参入到DNA分子中的量。液体闪烁计数法 用液体闪烁计数器测定培养物中的放射源因修复合成而参入到 DNA分子中的量。这一方法适用于大批量样本。
超速离心法 一种应用比较广泛的方法,可应用于切除修复、复制后修复及链断裂修复方式的检测。一般是用氚标记溴脱氧尿嘧啶核苷
 等参入到修复合成的 DNA分子中去以改变DNA分子的重量(BrdU的分子量比尿嘧啶核苷大),通过超速离心可以从沉降系数不同的各组分中收集修复合成中参入量不同的DNA片断,然后分别测定其放射性的强度来判断修复合成的多少。
等参入到修复合成的 DNA分子中去以改变DNA分子的重量(BrdU的分子量比尿嘧啶核苷大),通过超速离心可以从沉降系数不同的各组分中收集修复合成中参入量不同的DNA片断,然后分别测定其放射性的强度来判断修复合成的多少。病毒宿主细胞复活法 以SV40病毒、腺病毒、疱疹病毒、噬菌体等感染培养的人体细胞或细菌,然后以紫外线等处理以造成病毒DNA分子的损伤,因为病毒DNA分子损伤的修复是靠宿主细胞的修复酶系统,所以受损伤的病毒能否继续生存繁殖可间接地反映宿主细胞的修复功能。
姐妹染色单体互换(SCE)法 姐妹染色单体互换率的检测也能反映一部分 DNA修复功能。人类中的某些先天性DNA修复缺陷疾病如布卢姆氏综合征患者的自发SCE显著增高;另一些如着色性干皮病则诱发SCE增高。这是由于DNA修复功能的缺陷导致染色体稳定性减弱所致。
实践意义 DNA修复与肿瘤 各种原因引起的DNA损伤可以通过各种方式修复。如果修复功能有缺陷,DNA损伤就可能造成两种结果:一是细胞死亡;二是发生基因突变,或进而恶性转化为肿瘤细胞。先天性DNA修复缺陷疾病患者容易发生各种恶性肿瘤,例如人类的着色性干皮病患者的皮肤对阳光过度敏感, 照射后出现红斑、水肿,继而出现色素沉着、干燥、角化过度,结果可导致黑色素瘤、基底细胞癌、鳞状上皮癌及棘状上皮瘤的发生。通过细胞融合的研究表明具有不同临床表现的该病患者有明显的遗传异质性,可以分为A、B、C、D、E、F、G七个互补群及变种,A-G互补群表现为不同程度的核酸内切酶缺乏引起的切除修复功能缺陷,变种的切除修复功能正常,但复制后修复的功能有缺陷。又如范可尼贫血临床主要表现的特征如再生障碍性贫血、生长迟缓、易患白血病等是由于先天性链交联等修复缺陷所致。其他如布卢姆氏综合征和毛细血管扩张共济失调患者都易患白血病和淋巴肉瘤,也是先天性DNA修复缺陷造成的。
值得注意的是 DNA修复功能缺陷虽可引起肿瘤的发生,但已癌化细胞本身的 DNA修复功能并不低下,相反地却显著地升高,并能够充分地修复化疗药物引起的 DNA损伤, 这也是大多数抗癌药物不能奏效的原因。地鼠细胞的 DNA损伤修复的方式以复制后修复为主, 如果在地鼠的浆细胞瘤细胞的培养物中加入环磷酰胺等抗癌药后,瘤细胞照样生长,如果加入环磷酰胺的同时再加入咖啡因(复制后修复的抑制剂),则瘤细胞的生长受到了明显的抑制。所以 DNA修复的研究可为肿瘤联合化疗提供方案。
DNA修复与衰老 从 DNA修复功能的比较研究中发现寿命长的动物(象、牛等)修复功能较强;寿命短的动物 (仓鼠、小鼠、鼩鼱等)修复功能较弱。人的DNA修复功能也很强,但到一定年龄后逐渐减弱,同时突变细胞数也相应增加,所以老年人癌的发病率也比较高。检测各年龄组正常人的染色体畸变率和 DNA修复功能证实了这一点。人类中常染色体隐性遗传的早老症和韦尔纳氏综合征患者一般早年死于心血管疾病或恶性肿瘤;患者的体细胞极易衰老,是研究老年病与DNA修复关系的很好模型。
DNA修复与免疫 DNA修复功能先天缺陷的病人的免疫系统也常是有缺陷的,主要是 T淋巴细胞功能的缺陷。随着年龄的增长细胞中的DNA修复功能逐渐衰退,如果同时发生免疫监视机能的障碍,便不能及时清除癌化的突变细胞,从而导致发生肿瘤。所以, 衰老、DNA修复、免疫和肿瘤四者是紧密关联的。
DNA修复与环境致癌因子的检测 DNA修复的研究已被应用于检测各种化学致癌物。一般的方法是在体外传代培养的正常人皮肤成纤维细胞或大鼠原代培养的肝细胞中加入被检物,培养一定时间后再加入
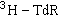 继续培养,然后收集细胞作放射自显影或液体闪烁的测试,如果参入量显著增高,表明被检物可疑为诱变剂或致癌剂。微生物培养的方法则更为简便、迅速,例如可以用枯草杆菌重组功能发生缺陷的突变型来进行检测,这些突变型由于丧失了重组功能而不能进行重组修复,因而更容易为许多诱变剂和致癌剂所杀伤致死。
继续培养,然后收集细胞作放射自显影或液体闪烁的测试,如果参入量显著增高,表明被检物可疑为诱变剂或致癌剂。微生物培养的方法则更为简便、迅速,例如可以用枯草杆菌重组功能发生缺陷的突变型来进行检测,这些突变型由于丧失了重组功能而不能进行重组修复,因而更容易为许多诱变剂和致癌剂所杀伤致死。关于 DNA修复机制方面的许多问题还有待于进一步的研究阐明。例如从原核生物开始到真核生物的高等哺乳类动物各依靠哪些方式来修复受损伤的DNA分子,修复方式又是怎样随物种的进化而发生演变的,修复缺陷的遗传异质性的本质又是什么,免疫缺陷和DNA修复功能缺陷的因果关系又是怎样的等等。
参考书目
P.C.Hanawalt, et al, DNA Repair Mechanisms,Academic Press,New York,1978.
L.Y.Malineux, et, al,Enzyme Involved in Repairof Damaged DNA, Plenum Press,New York & London,1980.