基因(卷名:生物学)
gene
━━━━━━━━━━━━━━━━━━━━━━━━认识的发展
功能、类别和数目
功能
类别
编码蛋白质的基因
没有翻译产物的基因
不转录的DNA区段
数目
相互作用
非等位基因的相互作用
互补基因
异位显性基因
累加基因
修饰基因
抑制基因
调节基因
微效多基因
背景基因型
等位基因的相互作用
无效基因
亚效基因
超效基因
新效基因
反效基因
镶嵌显性
基因和环境因素的相互作用
内在环境
性别
年龄
背景基因型
外在环境
温度
营养
演化━━━━━━━━━━━━━━━━━━━━━━━━
含特定遗传信息的核苷酸序列,是遗传物质的最小功能单位。除某些病毒的基因由核糖核酸 (RNA)构成以外,多数生物的基因由脱氧核糖核酸 (DNA)构成,并在染色体上作线状排列。基因一词通常指染色体基因。在真核生物中,由于染色体都在细胞核内,所以又称为核基因。位于线粒体和叶绿体等细胞器中的基因则称为染色体外基因、核外基因或细胞质基因,也可以分别称为线粒体基因、质粒和叶绿体基因。
在通常的二倍体的细胞或个体中,能维持配子或配子体正常功能的最低数目的一套染色体称为染色体组或基因组,一个基因组中包含一整套基因。相应的全部细胞质基因构成一个细胞质基因组,其中包括线粒体基因组和叶绿体基因组等。原核生物的基因组是一个单纯的DNA或RNA分子,因此又称为基因带,通常也称为它的染色体。
基因在染色体上的位置称为座位,每个基因都有自己特定的座位。凡是在同源染色体上占据相同座位的基因都称为等位基因。在自然群体中往往有一种占多数的(因此常被视为正常的)等位基因,称为野生型基因;同一座位上的其他等位基因一般都直接或间接地由野生型基因通过突变产生,相对于野生型基因,称它们为突变型基因。在二倍体的细胞或个体内有两个同源染色体,所以每一个座位上有两个等位基因。如果这两个等位基因是相同的,那么就这个基因座位来讲,这种细胞或个体称为纯合体;如果这两个等位基因是不同的,就称为杂合体。在杂合体中,两个不同的等位基因往往只表现一个基因的性状,这个基因称为显性基因,另一个基因则称为隐性基因。在二倍体的生物群体中等位基因往往不止两个,两个以上的等位基因称为复等位基因。不过有一部分早期认为是属于复等位基因的基因,实际上并不真正的等位,而是在功能上密切相关、在位置上又邻接的几个基因,所以把它们另称为拟等位基因。某些表型效应差异极少的复等位基因的存在很容易被忽视,例如从不同地区采集的黑腹果蝇的复眼虽然都是暗红色的,可是对它们的复眼色素进行定量分析后,发现不同的品系或个体还是能够区分的,而且通过特殊的遗传学分析可以分辨出存在于野生群体中的几个等位基因。像这种从性状上难以区分的复等位基因称为同等位基因。许多编码同工酶的基因也是同等位基因。
属于同一染色体的基因构成一个连锁群(见连锁和交换)。基因在染色体上的位置一般并不反映它们在生理功能上的性质和关系,但它们的位置和排列也不完全是随机的。在细菌中编码同一生物合成途径中有关酶的一系列基因常排列在一起,构成一个操纵子(见基因调控);在人、果蝇和小鼠等不同的生物中,也常发现在作用上有关的几个基因排列在一起,构成一个基因复合体或基因簇或者称为一个拟等位基因系列或复合基因。
在孟德尔定律发现以前,有所谓的融合遗传学说,认为双亲的遗传物质在子代中就象血液那样混合,一旦混合后,双亲的遗传物质只能被稀释而不能再分开。但孟德尔的实验结果表明亲代的隐性基因并不在杂交子一代中消失,因为由它所决定的性状还能在子二代中出现。与融合遗传学说相对立的颗粒性遗传学说则强调基因在世代间传递时于行为和功能上的相对独立性──基因的颗粒性。
认识的发展
从孟德尔定律的发现到现在,100多年来人们对基因的认识在不断地深化。
1866年,奥地利学者G.J.孟德尔在他的豌豆杂交实验论文中,用大写字母A、B等代表显性性状如圆粒、子叶黄色等,用小写字母a、b等代表隐性性状如皱粒、子叶绿色等。他并没有严格地区分所观察到的性状和控制这些性状的遗传因子。但是从他用这些符号所表示的杂交结果来看,这些符号正是在形式上代表着基因,而且至今在遗传学的分析中为了方便起见仍沿用它们来代表基因。
20世纪初孟德尔的工作被重新发现以后,他的定律又在许多动植物中得到验证。1909年丹麦学者W.L.约翰森提出了基因这一名词,用它来指任何一种生物中控制任何性状而其遗传规律又符合于孟德尔定律的遗传因子,并且提出基因型和表(现)型这样两个术语,前者是一个生物的基因成分,后者是这些基因所表现的性状。
1910年美国遗传学家兼胚胎学家T.H.摩尔根在果蝇中发现白色复眼 (white eye,W)突变型,首先说明基因可以发生突变,而且由此可以知道野生型基因W+具有使果蝇的复眼发育成为红色这一生理功能。1911年摩尔根又在果蝇的 X连锁基因白眼和短翅两品系的杂交子二代中,发现了白眼、短翅果蝇和正常的红眼长翅果蝇,首先指出位于同一染色体上的两个基因可以通过染色体交换而分处在两个同源染色体上。交换是一个普遍存在的遗传现象,不过直到40年代中期为止,还从来没有发现过交换发生在一个基因内部的现象。因此当时认为一个基因是一个功能单位,也是一个突变单位和一个交换单位。
40年代以前,对于基因的化学本质并不了解。直到1944年 O.T.埃弗里等证实肺炎双球菌的转化因子是DNA,才首次用实验证明了基因是由 DNA构成。
1955年S.本泽用大肠杆菌T4噬菌体作材料,研究快速溶菌突变型rⅡ的基因精细结构,发现在一个基因内部的许多位点上可以发生突变,并且可以在这些位点之间发生交换,从而说明一个基因是一个功能单位,但并不是一个突变单位和交换单位,因为一个基因可以包括许多突变单位(突变子)和许多重组单位(重组子)(见互补作用)。
1969年J.夏皮罗等从大肠杆菌中分离到乳糖操纵子,并且使它在离体条件下进行转录,证实了一个基因可以离开染色体而独立地发挥作用,于是颗粒性的遗传概念更加确立。随着重组DNA技术和核酸的顺序分析技术的发展,对基因的认识又有了新的发展,主要是发现了重叠的基因、断裂的基因和可以移动位置的基因。
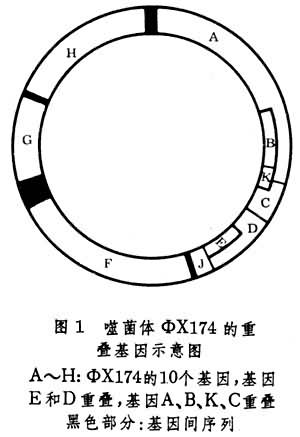
重叠基因是在1977年发现的。早在1913年A.H.斯特蒂文特已在果蝇中证明了基因在染色体上作线状排列,50年代对基因精细结构和顺反位置效应等研究的结果也说明基因在染色体上是一个接着一个排列而并不重叠。但是1977年F.桑格在测定噬菌体ΦX174的DNA的全部核苷酸序列时,却意外地发现基因D中包含着基因E(图1)。基因E的第一个密码子(见遗传密码)从基因D的中央的一个密码子TAT的中间开始,因此两个部分重叠的基因所编码的两个蛋白质非但大小不等,而且氨基酸也不相同。在某些真核生物病毒中也发现有重叠基因。
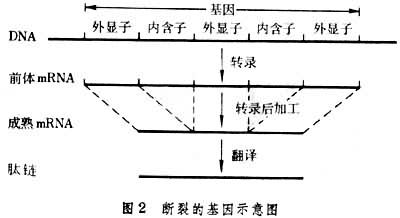
断裂的基因也是在1977年发现的,它是内部包含一段或几段最后不出现在成熟的mRNA中的片段的基因。这些不出现在成熟的mRNA中的片段称为内含子,出现在成熟的mRNA中的片段则称为外显子。例如下面这一基因(图2)有三个外显子和两个内含子。在几种哺乳动物的核基因、酵母菌的线粒体基因以及某些感染真核生物的病毒中都发现了断裂的基因。内含子的功用以及转录后的加工机制是真核生物分子遗传学的一个吸引人的课题。
可以移动位置的基因(见转座因子)首先于40年代中在玉米中由B.麦克林托克发现,当时并没有受到重视。60年代末在细菌中发现一类称为插入序列的可以转移位置的遗传因子IS,它们本身没有表型效应,可是在插入别的基因中间时能引起插入突变。70年代早期又发现细菌质粒上的某些抗药性基因可以转移位置。细菌中的这类转座子(Tn)到80年代已经发现不下20种,它们分别带有不同的抗药性基因,能在不同的复制子之间转移位置,例如从质粒转移到染色体、噬菌体以及别的质粒上等。当他们转移到某一基因中间时,便引起一个插入突变。类似于细菌转座子的可以转移位置的遗传因子在玉米以外的真核生物中也已经发现,例如酵母菌中的接合因子基因,以及果蝇白眼基因中的转座因子等。转座因子的研究也已成为分子遗传学中的一个重要方面。
功能、类别和数目
功能 到目前为止在果蝇中已经发现的基因不下于1000个,在大肠杆菌中已经定位的基因大约也有1000个,由基因决定的性状虽然千差万别,但是许多基因的原初功能却基本相同。
1945年G.W.比德尔通过对脉孢菌的研究,提出了一个基因一种酶假设,认为基因的原初功能都是决定蛋白质的一级结构(即蛋白质中各种氨基酸的排列顺序)。这一假设在50年代得到充分的验证。虽然以后在血红蛋白等的研究工作中,发现一个蛋白质可以由不同的肽链构成,而一个突变只影响一个肽链,因而把这一假设修改成为一个基因决定一种多肽。可是这一修改并没有改变前一假设的基本原理:基因的原初功能是编码组成肽链的氨基酸序列。即使重叠基因的发现也没有改变这一原理,因为在这里并不是把一个DNA片段看作一个基因,而是把编码某一肽链的一段 DNA序列看作一个基因。所以组成一段DNA的核苷酸序列如果有两种读码方式,这一段DNA就是两个基因。同样理由,断裂基因的发现也没有改变这一原理。
类别 60年代初F.雅各布和J.莫诺发现了调节基因。把基因区分为结构基因和调节基因是着眼于这些基因所编码的蛋白质的作用:凡是编码酶蛋白、血红蛋白、胶原蛋白或晶体蛋白等蛋白质的基因都称为结构基因;凡是编码阻遏或激活结构基因转录的蛋白质的基因都称为调节基因。但是从基因的原初功能这一角度来看,它们都编码蛋白质。根据原初功能(即基因的产物)基因可分为:
编码蛋白质的基因 包括编码酶和结构蛋白的结构基因以及编码作用于结构基因的阻遏蛋白或激活蛋白的调节基因;
没有翻译产物的基因 转录成为 RNA以后不再翻译成为蛋白质的转移核糖核酸(tRNA)基因和核糖体核酸(rRNA)基因;
不转录的DNA区段 如启动区、操纵基因等等。前者是转录时RNA多聚酶开始和DNA结合的部位;后者是阻遏蛋白或激活蛋白和DNA结合的部位(见基因调控)。
在真核生物的基因组中,除了编码血红蛋白等的结构基因以外,是否普遍存在着相当于细菌基因组中的调节基因还有待进一步研究。在许多真核生物的基因组中还有大量高度重复的脱氧核糖核酸重复顺序。重复顺序的功能的研究也是分子遗传学中的一个重要课题。
一个成长的个体的性状并不只决定于它有多少种蛋白质,还决定于这些蛋白质出现的时间和地点,各种细胞和组织的发生和功能都有一定的时空顺序,如果各种蛋白质的出现和活动的时空关系发生紊乱,那么即使全部蛋白质都存在,发育还是不能正常进行。已经发现在果蝇中有影响发育过程的各种时空关系的突变型,控制时空关系的基因有时序基因、格局基因、选择基因等(见发生遗传学)。
这些控制个体发育的基因是否都编码蛋白质,编码什么蛋白质,怎样控制发育途径,都是发生遗传学中有待解决的问题。
一个生物体内的各个基因的作用时间常不相同,这在最简单的生物如噬菌体也不例外。在大肠杆菌的λ噬菌体等病毒中,有一部分基因在复制前转录,称为早期基因;有一部分基因在复制后转录,称为晚期基因。早期基因包括和 DNA复制有关的基因以及晚期基因的激活物基因等。晚期基因包括头部蛋白和尾部蛋白基因以及使细菌裂解的内溶菌素的结构基因等。在哺乳动物细胞的病毒中也有早期基因和晚期基因之分。
基因往往是由于某一性状发生突变而被分辨和发现的,那么一个基因是否只有一种功能呢?当深入研究时也曾发现除某一性状发生突变外,其他性状也会受到影响。例如白色复眼突变型果蝇除了复眼变为白色以外,受精囊的长宽之比也明显地和野生型不同。一个基因发生突变而使几种看来没有关系的性状同时改变,这个基因就称为多效基因。许多所谓多效基因的多效性往往可以追溯到一个原初的突变效应,例如家鸡中显性的翻毛基因F存在时,使它的羽毛上翻。这一特性使体内的热量容易散失,因此这种鸡的体温较正常鸡为低,代谢作用相应地变得更加旺盛,由此又带来心跳加快,心脏扩大,以至消化器官和排泄器官也随之发生变化,代谢作用上的这些改变又影响肾上腺和甲状腺等内分泌腺体,从而使生殖能力下降。这样的基因严格说来不能称为多效基因。
酵母菌的组氨酸合成过程中的三种酶(环化水解酶、焦磷酸水解酶和组氨酸脱氢酶)的活性都和一种分子量为95000的蛋白质有关,由单个基因所编码。所以这是一个真正的多效基因。在脉孢菌等低等真核生物中也有类似的发现。
细菌的调节基因也可以称为多效基因。在大肠杆菌中和乳糖操纵子有关的调节基因 lacI 发生突变而成为lacIS的情况下,细胞中β-半乳糖苷透性酶、β-半乳糖苷酶和 β-半乳糖苷乙酰基转移酶就不再产生。虽然从基因的原初作用来讲,这一突变无非改变了阻遏蛋白和操纵基因 DNA的结合能力,但是从另一种意义来讲,可以认为lacI是一个多效基因,因为在这里一个基因发生突变时 3种不同的蛋白质都不再出现。启动区和操纵基因也同样可以看作是多效基因。
数目 已经确知RNA噬菌体MS2只有三个基因,它们都是结构基因,一个编码复制酶蛋白,另一个编码感染作用中所必要的A蛋白。对于一些较为复杂的DNA噬菌体来讲,它们需要更多的蛋白质来执行更多的功能,基因数目也相应地增加(表1)。

到1983年为止,大肠杆菌中已经定位的基因有1027个,这一数目也和根据基因组的分子量所预期的基因数属于同一数量级。按照基因组的分子量计算,哺乳动物的每一细胞中的基因数至少应该是 100万个。这一数目远远超出它所必需的基因数。事实上发现哺乳动物的基因组中极大部分是重复序列。在非重复的序列中,编码肽链的基因估计不过占百分之几,大约不超过10万个,其余的部分以及大部分重复序列是否和基因调控有关,现在还不清楚。真核生物的基因调控是分子遗传学中的一个重要研究领域。
到现在为止所发现的最简单的生物,是一类没有蛋白质外壳的类病毒如马铃薯纺锤状块茎类病毒(简称为PSTV)。从它的RNA的分子量来看,它只可能有一个基因。不过迄今还没有发现它编码任何蛋白质,也不知道它怎样引起宿主的病变。此外还发现了 DNA类病毒。地球上最初的生命形式不可能是寄生的,但必定是十分简单的,很可能是既单纯又较小的DNA或RNA分子。生命起源问题当然也是生物学中普遍感到兴趣的问题。
对于多数基因来讲,每一基因组中只有一个基因序列,但是对于某些基因来讲却不是这样。不论是原核生物或是真核生物,每一基因组中的rRNA基因和几乎每一种tRNA基因都不止一个。据估计,对于每一个基因组中每一种tRNA基因来讲,酵母菌有5~7个,果蝇有8个,而非洲爪蟾则有200个。非洲爪蟾的rRNA前体基因估计有500个。组蛋白是真核生物染色体的一个成分,组蛋白基因也有很多。据估计,对于每一种组蛋白基因来讲,非洲爪蟾有10~20个,海胆则有300~1000个。
除了这些单纯的重复基因以外,还有一些结构和功能都相似的为数众多的基因,它们往往紧密连锁,构成所谓基因复合体或叫做基因家族,例如哺乳动物中编码免疫球蛋白的基因(见免疫遗传学)、编码组织相容性抗原的基因等。
相互作用
生物的一切表型都是蛋白质活性的表现。生物体内的蛋白质多数是酶,而多数性状是许多酶作用的结果,所以生物的多数性状都不是单个基因决定的。换句话说,生物的各种性状几乎都是基因相互作用的结果。所谓相互作用,一般都是代谢产物的相互作用,只有少数情况涉及基因直接产物,即蛋白质之间的相互作用。基因转变曾经被认为是染色体在联会过程中一个基因同它的等位基因相互作用的结果,现在知道这是染色体交换的结果。
非等位基因的相互作用 依据非等位基因相互作用的性质可以将它们归纳为互补基因、异位显性基因、累加基因、修饰基因、抑制基因、调节基因和微效多基因等类型。
互补基因 若干非等位基因只有同时存在时才出现某一性状,其中任何一个发生突变时都会导致同一突变型性状,这些基因称为互补基因。例如在香豌豆中C(c)P(p)植株开紫花,ccP(p)或C(c)pp或ccpp植株都开白花,说明基因C和P对于紫色都属必要,任何一个发生突变时都使花色变白。又例如鼠伤寒沙门氏菌中组氨酸的合成从ATP和磷酸核糖焦磷酸开始,通过10个反应完成,这些酶为9个基因所编码。这9个基因对于组氨酸的合成都是必要的,其中任何一个发生突变都导致同一突变性状,即组氨酸缺陷型。所以它们也是互补基因。
互补基因的相互作用可以用下面这一通式表示,但这并不排斥其他的相互作用机制:
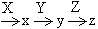 式中X、Y和Z是三个互补基因,x和y是中间产物,z是决定表型的最终产物。在这里x和y这两种中间产物没有表型效应。基因X、Y、Z对于z的产生都属必要,X或Y的突变和Z的突变一样导致同一突变型,即由z的缺乏所导致的表型。
式中X、Y和Z是三个互补基因,x和y是中间产物,z是决定表型的最终产物。在这里x和y这两种中间产物没有表型效应。基因X、Y、Z对于z的产生都属必要,X或Y的突变和Z的突变一样导致同一突变型,即由z的缺乏所导致的表型。异位显性基因 影响同一性状的两个非等位基因在一起时,得以表现性状的基因称为异位显性基因或称上位基因。例如在南瓜中,结白色瓜的品种和结黄色瓜或绿色瓜的品种的杂交子代都结白色瓜,说明白色对于黄色或绿色是异位显性。结黄色瓜的品种和结绿色瓜的品种的杂交子代都结黄色瓜,说明黄色对于绿色是异位显性。白色品种的基因型是WWYY或WWyy,黄色品种的基因型是wwYY,绿色品种的基因型是wwyy。
构窠曲霉的分生孢子是绿色的,黄色孢子和白色孢子突变型是两个非等位的突变型,它们对于绿色分生孢子来讲都是隐性的。黄色、白色杂合子产生白色分生孢子,说明白色对于黄色是异位显性。
异位显性基因的相互作用可以用下面这一通式表示,但是也不排斥其他相互作用机制:
式中X、Y、Z和x、y、z同样代表基因和代谢产物,与互补基因不同的是中间产物x、y也具有表型效应,当基因Z发生突变时z不能形成而表现突变性状y,当基因Y发生突变时产物y不能形成而表现突变性状x,在Y和Z都发生突变的个体表现突变性状x,所以Y对于Z是异位显性。累加基因 对于同一性状的表型来讲几个非等位基因中的每一个都只有部分的影响,这样的几个基因称为累加基因或多基因。例如野生型脉孢菌的分生孢子是橙红色。一个基因的突变使颜色变为桃红色,另一个基因的突变使颜色变为象牙色,两个突变基因同时存在时分生孢子呈白色。已经知道野生型细胞中含有几种胡萝卜素,橙红颜色是这许多色素同时存在的结果,也是许多基因效应累加的结果,其中某一或某些基因发生突变时,在不同程度上影响颜色。
累加基因的相互作用可以用如下这一通式表示,但也不排斥其他相互作用机制:
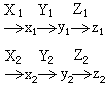 式中z1和z2是对于同一性状各有一定程度的作用的代谢最终产物,例如不同的胡萝卜素。合成不同胡萝卜素的基因对于分生孢子的颜色来讲都是累加基因。在累加基因中每一个基因只有较小的一部分表型效应,所以又称为微效基因。相对于微效基因来讲,由单个基因决定某一性状的基因称为主效基因。
式中z1和z2是对于同一性状各有一定程度的作用的代谢最终产物,例如不同的胡萝卜素。合成不同胡萝卜素的基因对于分生孢子的颜色来讲都是累加基因。在累加基因中每一个基因只有较小的一部分表型效应,所以又称为微效基因。相对于微效基因来讲,由单个基因决定某一性状的基因称为主效基因。修饰基因 本身具有或者没有任何表型效应,可是和另一突变基因同时存在便会影响另一基因的表现程度的基因。如果本身具有同一表型效应则和累加基因没有区别。
抑制基因 一个基因发生突变后使另一突变基因的表型效应消失而恢复野生型表型,称前一基因为后一基因的抑制基因。如果前一基因本身具有表型效应则抑制基因和异位显性基因没有区别。
调节基因 一个基因如果对另一个或几个基因具有阻遏作用或激活作用则称该基因为调节基因。调节基因通过对被调节的结构基因转录的控制而发挥作用。具有阻遏作用的调节基因不同于抑制基因,因为抑制基因作用于突变基因而且本身就是突变基因,调节基因则作用于野生型基因而且本身也是野生型基因。许多调节基因的直接产物只有阻遏蛋白或激活蛋白的功能,没有其他的表型效应;可是某些调节基因直接产物还具有酶的活性,所以它们既是调节基因,又是结构基因。例如大肠杆菌的组氨酸合成中的磷酸核糖焦磷酸磷酸化酶便是一种阻遏蛋白,编码这种蛋白质的基因hisG既是结构基因又是调节基因。它和其他组氨酸基因的关系既是互补基因的关系,又是调节基因和结构基因的关系。
微效多基因 影响同一性状的基因为数较多,以致无法在杂交子代中明显地区分它们的类型,这些基因统称为微效多基因或称多基因。在和生产有关的数量性状的遗传中,往往涉及微效多基因遗传,例如高度、产量的遗传等。微效多基因遗传是数量遗传学和育种学的主要研究对象。
背景基因型 从理论上看,任何一个基因的作用都要受到同一细胞中其他基因的影响,在许多遗传学工作中也都可以看到在同一基因引入不同的品系时,它的表型效应常有不同。除了人们正在研究的少数基因以外,其余的全部基因构成所谓的背景基因型或称残余基因型。
等位基因的相互作用 孟德尔发现两个不同的遗传因子(即等位基因)同时存在时,其中只有一个的表型效应得以显示,这是一种最简单的等位基因之间的相互作用即完全显性。但是杂合体也可以是中间状态的,例如紫茉莉 (Mirabilis jalapa)的红花基因(red,R)和白花基因 (white,r)杂合体的花色是浅红的。这一现象称为不完全显性。1932年H.J.马勒依据突变型基因与野生型等位基因的关系归纳为无效基因、亚效基因、超效基因、新效基因和反效基因。
无效基因 不能产生野生型表型的、完全失去活性的突变型基因。无效基因对于野生型基因来讲都是隐性的,即使带有两个隐性无效基因和一个野生型基因的个体,它的表型仍是野生型的而不是突变型。果蝇的白眼基因和造成完全遗传性障碍的营养缺陷型基因都是无效基因。这些都是基因突变使有关的酶完全丧失活性的结果。一个基因的缺失,从遗传学效应来看也是一个无效突变,所不同的是缺失不能回复,而一般的无效基因却能通过回复突变而成为野生型基因。
亚效基因 表型效应在性质上相同于野生型,可是在程度上次于野生型的突变型基因。亚效基因对于野生型等位基因来讲也是隐性的,虽然它导致某种突变型性状,可是并不随着亚效基因数量的增加而带来更为极端的突变型表型,相反地却接近于野生型表型。例如果蝇的截毛基因 (bobbed,bb)导致刚毛变细而短,这一基因既存在于X染色体上也存在于Y染色体上。正常的雌性果蝇的性染色体是XX,正常雄性果蝇是XY,也可以得到例外的具有三个性染色体的果蝇XXY或XYY,并且使它们分别带有三个或两个截毛基因。实验结果说明具有三个bb基因的果蝇的刚毛的长度超过具有两个bb基因的果蝇而接近于野生型果蝇。微生物中的渗漏性营养缺陷型也是一类亚效基因,它们是由于基因突变而使有关的酶活性降低的结果。
超效基因 表型效应超过野生型等位基因的突变型基因。由于自然选择的结果,野生型基因所编码的酶往往具有最高的活性,因此几乎任何突变都使酶的活性下降。可是这决不是说对于任何一种酶来说,野生型基因的结构都是十全十美的。对于世代周期很短的细菌来讲,不难在实验室中对细菌群体长久地施加某一选择压力,从而得到某种酶的活性超过野生型的突变型。例如在腐臭假单孢菌(Pseudomonas putida)中从只能稍稍利用2-氯丙酸钠但不能利用除莠剂2,2-二氯丙酸钠作为碳源的野生型细菌中获得了能充分利用后者的突变型细菌。从只能分解乙烯胺和丙烯胺的细菌获得能分解丁烯胺、戌烯胺、苯丙烯胺和乙酰苯胺的突变型细菌,而且通过实验证明这是单一基因突变的结果。这些突变型基因便是超效基因。
新效基因 产生野生型等位基因所没有的新性状的突变型基因。果蝇中的一个显性的突变基因毛翅(hairy wing,Hw)的存在使翅上出现野生型所没有的细毛,这是一个新效基因。不同的抗原基因和血型基因对于它们的原始基因来说都是新效基因,这是因为抗原和血型都是蛋白质分子的直接反映。新效基因对于野生型等位基因来讲彼此都是显性,所以这种情况称为共显性。许多无效基因在细胞中往往没有相应的酶活性,这倒不是因为没有产生相应的蛋白质,而是产生了没有酶活性的蛋白质,这些蛋白质和原来的具有活性的蛋白质在血清学反应上具有交叉反应,所以称为交叉反应物质。因此从分子水平来看,许多突变型基因是新效基因,不过由于所编码的蛋白质没有活性,所以从表型效应来看是无效基因。
反效基因 作用和野生型等位基因相对抗的突变型基因。例如粗糙脉孢菌的谷氨酸脱氢酶基因通过突变成为一系列突变型,其中一个突变型am19与野生型联合形成的异核体的生长能力不如野生型,而且它的谷氨酸脱氢酶的活力低于野生型。这是因为谷氨酸脱氢酶由相同的几个亚基构成,在上述异核体中野生型基因产物和突变型等位基因产物结合在一起,构成活力较低的谷氨酸脱氢酶。在这里野生型对于突变型来讲是不完全显性。这种抑制野生型基因作用的基因便是反效基因。如果突变型基因的直接产物和野生型等位基因的直接产物相互作用的结果不是低于而是高于野生型等位基因的酶活性,就称为超显性。
镶嵌显性 对于某一性状来讲,一个等位基因影响身体的一个部分,另一等位基因则影响身体的另一部分,而在杂合体中两个部分都受到影响的现象称为镶嵌显性。在异色瓢虫 (Harmonia axyridis)的鞘翅色斑遗传、果蝇的盾片刚毛遗传和玉米的花青素遗传中都有这种现象。镶嵌显性也是一种共显性现象,它的基因相互作用机制还有待于研究。
除了以上这些等位基因之间的相互关系以外,在免疫遗传学中还有等位排斥现象。免疫球蛋白杂合体只产生一种免疫球蛋白,似乎两个等位基因中只有一个是活动的,而另一个则被排斥。等位排斥现象中的等位基因相互排斥作用机制还有待研究。
基因和环境因素的相互作用
基因作用的表现离不开内在的和外在的环境的影响。在具有特定基因的一群个体中,表现该基因性状的个体的百分数称为外显率;在具有特定基因而又表现该一性状的个体中,对于该一性状的表现程度称为表现度。外显率和表现度都受内在环境和外在环境的影响。
内在环境 指生物的性别、年龄等条件以及背景基因型。
性别 只在某一性别中表现的性状称为限性性状。例如奶牛的产乳量基因只在雌性奶牛中显示它的作用。在两个性别中都表现,可是表现程度不同的性状称为从性性状。例如同一早秃基因对男子的影响大于女子。性别对于基因作用的影响实际上是性激素对基因作用的影响。性激素为基因所控制,所以实质上这些都是基因相互作用的结果。
年龄 人类中各个基因显示它的表型的年龄有很大的区别(表2)。

背景基因型 通过选择,可以改变动植物品系的某一遗传性状的外显率和表现度,说明一些基因的作用往往受到一系列修饰基因或者背景基因型的影响。由于背景基因型的差异而造成的影响,在下述三种情况中可以减低到最低限度:①由高度近交得来的纯系;②一卵双生儿;③无性繁殖系(包括某些高等植物的无性繁殖系、微生物的无性繁殖系以及高等动物的细胞株)。用这些体系作为实验系统,可以更为明确地显示环境因素的影响,更为确切地说明某一基因的作用。双生儿法在人类遗传学中的应用及纯系生物在遗传学和许多生物学研究中的应用都是根据这一原理。
外在环境 温度 温度敏感突变型只能在某些温度中表现突变型性状,例如温度敏感的致死突变型只在某些温度中致死,而在另一些温度中可以存活。对于一般的突变型来说,温度对于基因的作用也有程度不等的影响。在果蝇中,提高饲养温度可使第四翅脉中断突变型(cubitus interuptus, ciD)的外显率下降,使棒眼突变型(bar,B)的表现度上升。
营养 家兔脂肪的黄色决定于基因 y的纯合状态以及食物中的叶黄素的存在。如果食物中不含有叶黄素,那么 yy纯合体的脂肪并不呈黄色。y基因的作用显然和叶黄素的同化有关。微生物中的营养缺陷型在所需要的物质不存在的情况下是致死突变型,如果存在则和野生型难以区别。由于先天的苯丙氨酸代谢发生障碍而出现苯丙酮尿症的病儿,如果出生后立即限制食物中的苯丙氨酸则可使儿童发育完全正常。
演 化
就细胞中 DNA的含量来看,一般愈是低等的生物含量愈低,愈是高等的生物含量愈高。就基因的数量和种类来讲,一般愈是低等的生物愈少,愈是高等的生物愈多。DNA 含量和基因数的增加与生理功能的逐渐完备是密切相关的。对于生物进化过程中基因的数目怎样增加而使生理功能逐渐完备,美国遗传学家N.霍罗威茨提出了如下假说:
在从无生命物质产生生命的时刻,地球表面充斥着各式各样的有机化合物,可以设想最初的生物无需自己合成各种氨基酸和核苷酸等而可以从周围环境中取得它们。所以又可以设想这些最原始生物不具备今天地球上的生物所具备的那些有关合成氨基酸和核苷酸的酶也能生存繁衍。能自己合成这些物质的生物是在以后的进化过程中逐步出现的。
下述从 a到z的一系列反应中,每一步应分别由酶A、B、…、X、Y、Z催化。
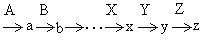
与A…Z这一系列酶相对应的基因,在进化过程中同时出现的可能性很小,而且从现有生物的氨基酸或核苷酸的代谢过程来看,a…y的中间产物除了导致终产物z的合成以外,很少具有其他的生理意义,因此在进化过程中基因逐个地出现的次序不可能是先有A,然后出现B…以至于Z。因为在出现Z以前,基因A…Y都是无用的。很难想象这样许多无用的基因能在长期进化过程中保存下来。但是当地球上各种有机化合物由于大量生物的生长繁殖而被消耗时,若偶然出现一个基因Z而使y产生终端产物z,就会有利于携带Z基因生物的生存,因而可以长久保存这个基因。依次类推,以后也会出现基因Y、X等等,最后出现A。
随着代谢途径的进化,基因数目必定随之增加。那么,新的基因从何而来呢?从细胞遗传学研究结果可以看到,不对等交换可以带来一个染色体上一小部分染色体的重复和同源染色体上一小部分染色体的缺失(见位置效应)。基因突变往往给生物个体带来有害的甚至致死效应,可是如果通过任何机制而出现了重复,那么两个基因中的一个发生突变时不会对生物个体带来有害的效应。所以通过基因的一再重复和其中一个基因的突变,可以使一个生物个体中的新的基因不断地增加。这可能是对于进化过程中新基因和新功能演变的一种合理的解释。对于现有生物的蛋白质的研究结果也支持这一解释(见分子进化)。
基因最初是一个抽象的符号,后来证实它是在染色体上占有一定位置的遗传的功能单位。大肠杆菌乳糖操纵子中的基因的分离和离体条件下转录的实现进一步说明基因是实体。现在已经可以在试管中对基因进行改造(见重组DNA技术)甚至人工合成基因。对基因的结构、功能、重组、突变以及基因表达的调控和相互作用的研究始终是遗传学研究的中心课题。