有孔虫目(卷名:生物学)
Foraminifera
原生动物门肉鞭动物亚门根足纲粒网亚纲的1目。是具壳的海洋原生动物。约有3.4万种,其中6000多种为现代种。
形态结构 有孔虫的整个细胞质团由含各种不同矿物成分的壳体包裹着。原生质分外质、内质两层。外质色浅透明,可通过口孔或壳壁孔伸延壳外,成细线状或粒网状伪足,具有运动、摄食、消化、呼吸、排泄、分泌、造壳等功能。内质色深,有各种细胞器,如细胞核、环片、线粒体、微体、高尔基器、溶酶体、微管、黄色素体、共生体等。
有孔虫胞核的数目因种类而异,有单核的或多核的。多核种类又分异型核的和同型核的两类。在生殖发育阶段,也存在着二型核现象:有性生殖世代(显球型)是单核的;无性生殖世代(微球型)则为多核的。
壳体依壳壁组成成分及微细构造,可分为以下几种基本类型:①假几丁质壳,最原始类型,薄而易变形(如网足虫亚目);②胶结质壳,壳壁由自身分泌的物质胶结外来物质而成(如编织虫亚目);③钙质壳,主要由自身分泌的碳酸钙组成,依壳壁的结晶体和微细构造,可分为微粒状无孔壳(亚目)、似瓷质无孔壳(粟虫亚目)、钙质透明有孔单层放射状壳(瓶虫亚目)、钙质透明有孔多层粒状壳(轮形虫亚目)、钙质透明放射微粒状壳(包旋虫亚目)、钙质透明单晶壳(旋虫亚目)、钙质骨针壳(卡特虫亚目);④硅质壳,壳壁成分为二氧化硅(如硅室虫亚目)。
壳体有单房室、双房室或多房室。房室间的隔板有孔相通,有孔虫即以此特征而得名。单房室类群多数呈近球形或平旋管状;多房室类群的房室排列多数为螺旋式,其次为平旋、双列、盘状、粟虫式旋卷、单列等,还有混合排列,如从平旋到单列或双列,从双列到单列,从螺旋到三列等形式(见图)。
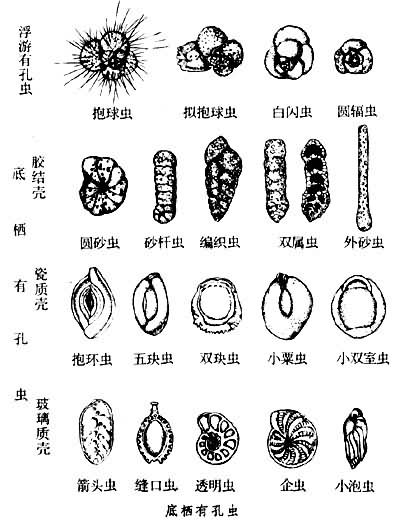
生殖与生活史 有孔虫的生殖方式为有性生殖的配子母体世代与无性生殖的裂殖体世代交替进行。成熟的裂殖体减数分裂,行无性裂配生殖,形成许多配子母体。配子母体成熟后,产生许多带有2根鞭毛的配子,每两个配子融合成合子,行有性配子生殖。从合子发育成裂殖体为二倍体期。从减数分裂进行裂殖生殖至配子母体产生配子,则为单倍体期。有性生殖配子母体世代的初房较大,称显球型壳;无性生殖体的壳体初房则较小,称微球型壳,这就是有孔虫壳体的二型现象。这种二型现象仅见于底栖种类,浮游有孔虫无此现象。有些底栖有孔虫配子的形态与鞭毛数目是有变化的,例如旋虫、帽贝虫的配子为变形虫状,不带鞭毛,有的种具3根鞭毛,还有如玫瑰虫在配子母体世代发育1个浮室以产生配子。 有孔虫经世代交替完成整个生活史所需时间一般为1~2年或更长时间,但也有只需半个月左右的。
生活习性与生态 有孔虫主要生活于正常海洋环境中,有些种类能适应半咸水、沼泽或超高盐泻湖等变化剧烈的环境,淡水种极少。多数种底栖生活,浮游性种类较少。有孔虫是微杂食性动物,也有腐食性的,有的在细胞质内有虫黄藻等共生藻类。
底栖有孔虫自潮间带至深海盆均有分布,分布最深记录是在千岛-堪察加海沟(10687米)。在东海,底栖有孔虫种类和数量的分布趋势是:河口区稀少,自内陆架向中、外陆架迅速增多;越过陆坡向冲绳海槽区,种类逐渐减少,数量则显著减少。浮游有孔虫一般是近岸稀少,越远离海岸,种类和数量越增加。浮游有孔虫有昼夜垂直移动现象。
盐度对有孔虫的影响很显著,在正常海洋盐度区,底栖有孔虫种类多、数量大;在低盐的河口区、泻湖和沼泽区,有孔虫种类贫乏;在高碳酸钙、超高盐的泻湖中,较常见的是粟虫科的种类(如三浃虫)。
温度是影响有孔虫分布的重要因素。在热带,有孔虫的生殖临界温度远比温、寒带的狭窄。 热带-亚热带有孔虫的种类与数量远比温、寒带繁多。浮游有孔虫的纬度分布和底栖有孔虫的深度分布,明显受到水温的制约,生活于热带深海盆的底栖冷水种,在高纬度则可分布于较浅海底。温度还可以影响有孔虫壳体的形态构造,如厚壁新方球虫在两极区,壳体左旋;在温暖海域,壳体则为右旋。
大洋环流控制着浮游有孔虫的地理分布。布拉德绍,1959年所划分北太平洋的4个浮游有孔虫区系(亚北极区系、过渡带区系、中央区系、西赤道中央区系)与北太平洋几个大水团的分布范围适相对应。马维骅1977年把世界大洋浮游有孔虫划分为3个生物地理区(环热带区、北冷水区、南冷水区)、5个省(北极与南极省、亚北极与亚南极省、南北过渡省、亚热带省、热带省)。大洋环流的界线与浮游有孔虫区系界线十分一致。
底质对有孔虫有明显影响,在富含有机质的粉砂和泥质底质中,有孔虫的数量常较多,且多数是薄壳或细长类型。在贫营养的砂或砂砾底质中,有孔虫数量常较少,且多数为厚壳透镜形或纺锤形类型。生活在硬底质如岩石、多贝壳或多海藻环境中的有孔虫,多数是便于贴附或附着的壳形。碳酸钙在暖水区的溶解度比在冷水区小,所以厚钙质壳有孔虫、有孔虫石灰岩和有孔虫软泥都出现在低纬度区。碳酸钙溶解度随着水深增加而增大。在碳酸钙溶解度变化最大的溶跃面以下,钙质有孔虫壳大多数被溶解,因而深海有孔虫动物群是以胶结种类占优势。
演化 最早出现于寒武纪-奥陶纪的古老有孔虫多为假几丁质或胶结质,构造为简单球形或袋状等原始类型。泥盆纪出现多房室类群、房室排列多样化,有双列、三列或多列。石炭纪出现房室为旋式排列的类,二叠纪为古生代有孔虫繁盛时期,以类和内卷虫占绝对优势,堪称为类时期,类发展到最高峰(已有5000种),成为石灰岩的主要组成部分。似瓷钙质无孔有孔虫,也在这一时期开始出现。晚古生代发生了剧烈的构造运动和大陆运动,海退大幅度改变了水圈条件,导致三叠纪有孔虫种类发展缓慢。到了侏罗纪,气候转暖而平稳、海浸广泛,世界有和暖的边缘海和环赤道暖流系。这一时期堪称有孔虫盛期,各种类型的钙质和胶结底栖有孔虫十分丰盛,并出现文石质壳的罗伯特虫类群,浮游有孔虫在此期开始出现。到白垩纪,浮游有孔虫更为丰盛,成为白垩纪优异的标准化石。但到了白垩纪晚期,大多数白垩纪浮游有孔虫和浅水底栖有孔虫绝灭,而大多数深水底栖有孔虫仍延续出现。这一事实提示了当时海洋环境的剧烈变化仅局限于200~300米的上面水层。第三纪初期古新世一些白垩纪浮游有孔虫类群如轮孔虫科、截球虫科、沙科虫科已全部绝灭,而抱球虫科成为最主要的浮游类群。同时,还出现汉京虫科、圆辐虫科等类群。始新世大型钙质底栖有孔虫,如货币虫科、鳞环虫科、蜂巢虫科等都很丰盛,形成石灰岩。中新世有孔虫基本具备了现代有孔虫的属种面貌。
意义 各个地质时期的有孔虫,种类演化明显,化石保存较好,常被用作确定地质年代的标准化石和古沉积环境的指相化石,特别是被用于恢复新生代的古气候、古海洋环境(如古水深、古温度、古盐度、古海流、古水团)以及估计沉积率等等。例如,根据岩心各层次所含有孔虫暖水种与冷水种的比率、某些种类壳的旋向与壳壁孔隙度的变化,或根据有孔虫钙质壳同位素O16与O18含量的比率,重建古温度变化曲线,恢复古气候;利用编织虫亚目、粟虫亚目和轮形虫亚目的比例关系,可以指示古盐度、恢复古环境;研究白垩纪浮游有孔虫的分布,可以重建白垩纪的古海流;根据P(浮游有孔虫)/B(底栖有孔虫)比值,可以指示离岸的方向与远近:根据现代浮游有孔虫参数,可以判别海流的趋向、洋峰的位置与水团的界限。在中国海区,浮游有孔虫是暖流的优越指示生物。利用有孔虫推断古沉积环境和地层对比,有助于石油勘探。
20世纪60年代以来,世界各国对各大洋有孔虫的调查研究,“深海钻探计划”(DSDP)的成功,电子显微镜和电子探针等新技术的应用,细胞学、生理学、生物化学、生态学等现代生物学概念以及新的研究手段、新的资料分析方法的引入,促使对有孔虫的研究向新的高度发展。