位置效应(卷名:生物学)
position effect
基因由于变换在染色体上的位置从而改变表型效应的现象。位置效应的研究对于了解染色体的结构和功能具有重要的意义。
美国遗传学家 A.H. 斯特蒂文特根据黑腹果蝇棒眼(Bar,B)突变型的研究结果,于1925年提出了位置效应概念。此后在果蝇的白眼、黄体等突变型,在月见草的萼片色和花瓣色、玉米籽粒的斑点,以及在小鼠的皮毛色等性状上都先后发现位置效应现象。美国学者E.B.刘易斯于1950年把位置效应分为稳定型和花斑型两大类型。
稳定型位置效应 简称S型位置效应,表型改变是稳定的。
果蝇的复眼由许多小眼组成。野生型的正常复眼呈椭圆形;棒眼突变型由于小眼数的显著减少而呈不同程度的狭棒形。棒眼基因B为显性,位于X染色体上。纯合的棒眼果蝇的后代中常出现少数野生型个体;同时出现少数复眼比棒眼更狭细的超棒眼个体。这两种个体出现的频率都约占1/1600,远远超过一般的突变频率。
斯特蒂文特在1925年曾用棒眼果蝇进行一系列不同组合的杂交,计数了后代各种棒眼的小眼数,同时注意分析与棒眼座位紧密连锁的基因的行为,发现野生型和超棒眼的个体的出现总是伴随着棒眼座位相邻基因的交换。针对这一情况,他提出不对等交换假说。这一假说在1936年为美国遗传学家H.J.马勒和C.B.布里奇斯所证实。他们发现在正常果蝇唾腺细胞的 X染色体上只有一个编号为16A的区段,而棒眼果蝇的X染色体上有连接着的两个16A区段,超棒眼果蝇中则有三个重复的16A区段(图1)。这说明棒眼果蝇来源于16A区段的重复,由棒眼果蝇产生的野生型和超棒眼个体是由于少数雌性棒眼果蝇的一对 X染色体在减数分裂过程中染色体发生了不对等交换,于是极少数配子中获得一个只有一个16A区段的X染色体,而同数的另一些配子中却获得一个有三个16A区段的X染色体。
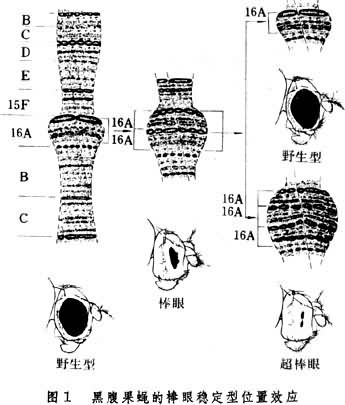
比较各种基因型的果蝇的每一复眼的小眼数,还发现棒眼纯合体B/B的复眼的小眼数是68,而正常/超棒眼杂合体+/BB的复眼的小眼数是45。两者的 16A区段数相同,只是由于它们所在染色体的位置不同而使小眼数不同,因此称之为位置效应。
花斑位置效应 简称 V型位置效应。这种效应的表型改变是不稳定的,因而导致显性和隐性性状嵌合的花斑现象。果蝇眼色的红、白嵌合,小鼠皮毛色的棕、灰嵌合,玉米籽粒的颜色斑点等现象都属于这一类型。除了颜色的花斑现象以外,在果蝇中还发现异淀粉酶和6-磷酸葡萄糖酸脱氢酶活性也由于位置效应而呈现花斑现象。花斑型位置效应起因于染色体的重新排列。一般当原来在常染色质区的基因被变换到接近异染色质区的位置时,便会出现花斑现象。
果蝇的白眼基因(white,w)位置在X染色体的末端的常染色质区。如果带有它的等位基因即野生型红眼基因(W+)的 X染色体末端被易位到另一染色体的异染色质区,例如易位到第四染色体的着丝粒附近,而与W+等位的白眼基因(w)仍处在正常X染色体的原末端位置上,则雌性易位杂合体果蝇(W+/w)的复眼呈现红白嵌合的花斑。如果被易位的基因是w而不是W+,则雌性果蝇(W+/w)的复眼还是正常的红色(图2)。
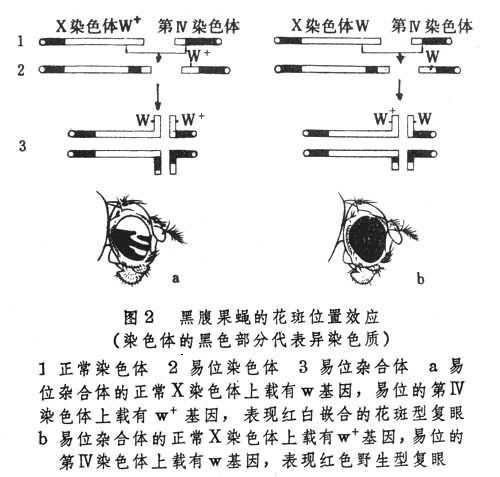
此外,月见草的萼片色和花瓣色的花斑是由于3~4和11~12两个染色体的互换产生的;小鼠的皮毛色的花斑是常染色体和X染色体易位的结果。
花斑位置效应的强度常受外界和内在条件的影响而有所变化。根据果蝇的研究,饲养温度的高低,Y染色体的增多等都对花斑表现的强弱有一定程度的影响。
花斑位置效应常具有扩散性。如果被转移的常染色质区段所包含的基因不止一个,而是两个以上,那么这几个基因同样可以由于位置变换而表现花斑现象。这种由于倒位或易位而使一系列基因同时失活的现象称为扩散性位置效应。例如果蝇的 X染色体的常染色质区有五个依次排列着的基因座位:粗糙眼(rough,rst)、小糙眼(facet,fa)、小体(diminutive,dm)、海胆眼(echinus,ec)和二裂翅 (bifid,bi)。如果由于染色体倒位而使这些座位的野生型基因顺序颠倒并且被连接到接近着丝粒的异染色质区,那么位置效应可自rst座位一直延伸到bi座位,各个基因将表现出不同程度的野生型性状和突变型性状相嵌合的花斑现象。在果蝇的唾腺染色体上,这种扩散性位置效应有时可延伸到50个横纹(相当于大约六个图距单位)。小鼠常染色体的一些基因如果被易位到X染色体上,那么由于相邻的异染色质的作用,也表现有不同程度的花斑现象。扩散性位置效应所表现的强弱与各个基因距离异染色质的远近有关,一般愈接近异染色质区的基因的位置效应愈强,愈远则位置效应愈微弱。
花斑位置效应比较普遍,但是它所表现的花斑现象经常容易与另一些类似现象相混淆,例如由于体细胞交换而出现的花斑(见连锁和交换)等。验证花斑位置效应的方法是通过染色体交换或诱发染色体畸变而使染色体排列恢复原来状态,然后观察表型是否同样恢复正常,如果表型不再呈现花斑状,那就说明这是一种花斑位置效应。有关果蝇的多毛基因(hairy,h)、白眼基因(w)、小鼠的棕色毛基因 (brown,b)等都曾通过这些方法证明花斑的出现来自位置效应。
玉米籽粒的颜色斑点也属于花斑位置效应。据美国遗传学家B.麦克林托克从50年代初到60年代中多年的研究,这种颜色斑点的出现与玉米基因组中所存在的控制因子有关(见转座因子)。例如在玉米的Ds-Ac系统(解离-激活因子系统)中,解离因子Ds和激活因子Ac是两个各自能在同一染色体上或各个染色体间转移位置的遗传因子。Ds必须在Ac存在的情况下才有作用;Ac除了激活Ds以外,没有其他表型效应。Ds能抑制相邻基因的表达,并能转移到另一染色体上,同时引起它原来位置附近的染色体断裂。在胚乳细胞中如果Ds处在使胚乳具有颜色的有色基因C的近旁,那么C的表达便被抑制而成为白色;如果Ds离开有色基因C,基因C便得以表达,表现为有色。Ds从有色基因C近旁解离的时间愈早,有色斑点愈大;反之则愈小。Ds的这种抑制作用是可扩散的,可以影响到相距五个图距单位的基因,在这点上它类似于果蝇和小鼠的花斑位置效应。Ac在任何位置上对于Ds都有激活作用;Ac对于Ds的激活作用随着Ac因子的数目增加而改变。基因型为ccC的胚乳是有色的,ccC Ds也是有色的,ccC Ds Ac、ccC Ds Ac Ac和 ccC DsAc Ac Ac的胚乳则都是白色而有颜色斑点,斑点的大小则依次递减而以后者为最小。Ds和Ac的关系类似于细菌的操纵子中的操纵基因和调节基因之间的关系(见基因调控),不过它们是可以改变位置的遗传因子。因此在这个意义上,它们又是一种转座因子。
位置效应的机制 从有关花斑位置效应的事例中可以看到如果基因位置转移到异染色质附近就会出现花斑效应,如果恢复原来位置则花斑效应便随着丧失。这说明基因由于染色体畸变而被转移到异染色质附近时,虽然它在功能上发生了改变,但基因本身并未发生改变。异染色质大部分是由较短的重复顺序所组成,它的螺旋化和固缩程度远远大于常染色质,在细胞分裂期间它的DNA复制落后于常染色质,大多不能转录。在果蝇中已经证实异淀粉酶活性和6-磷酸葡萄糖酸脱氢酶活性的花斑现象是由于某些细胞中两个有关基因的转录受阻而不是翻译受阻的缘故。目前对于位置效应的机制的了解还不很多,从染色体结构和基因调控角度进行深入的研究将会阐明位置效应的本质,同时位置效应的研究也将会加深对于染色体结构和功能的了解。