神经系统(卷名:生物学)
nervous system
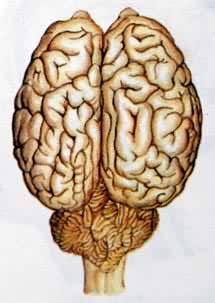
众多有组织的神经细胞(神经元)的集合体,是调节人和动物体内各种器官活动以适应内、外环境变化全部神经装置的总称。人的神经系统包含约1011~1012个神经元以及为数更多的神经胶质细胞。庞大的神经细胞数量以及神经细胞之间的复杂组织联系使神经系统具有复杂的功能。人和高等动物的神经系统是地球上最复杂的物质结构。神经系统的主要功能是接受和处理体内外各种感觉信息、调节和影响躯体和内脏的运动,维持机体内环境的相对稳定,发动和控制各种行为,司理学习、记忆、情绪、思维及语言等高级功能。在神经系统的直接或间接的控制、调节下,机体各器官系统才能相互联系,相互协调,完成统一的生理过程。脊椎动物和人的神经系统可分为中枢神经系统和外周神经系统两部分,前者包括脑和脊髓,后者包括外周神经和神经节。神经系统的形成以及结构和功能的完善是由低等无脊椎动物向高等脊椎动物长期进化的结果。(见彩图)。




无脊椎动物的神经系统 腔肠动物门 原始的神经系统。水螅及海葵的体壁上具有由间质细胞分化而成的神经分泌细胞、神经感觉细胞以及神经节细胞。这些细胞具有两个或多个长的突起,被称为轴索。轴索之间相互连接构成了神经网,即它们的神经系统(图1)。水母体壁上有一些神经细胞聚集,形成了简单的神经节。但是遍布体壁的仍然是神经网,而没有形成中枢系统。神经细胞及其所支配的细胞间形成突触,可以传递兴奋以影响被支配的细胞活动。神经网只借着弥散的兴奋传导以协调身体的运动,因而这类动物缺乏明确的定向活动。
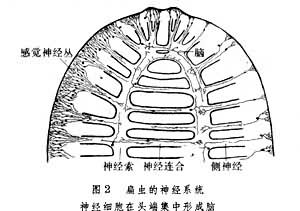
扁形动物门 开始出现中枢神经系统。动物的头端集中了相当数量的神经细胞,形成了明显的脑(图2)。由脑向身体后方发出两条神经索,延伸至动物的尾端。神经索之间又有横向的神经连合相互连接,构成形若绳梯的中枢神经系统。扁虫的脑内已经出现功能相同的神经细胞轴突集聚而形成的不同传导束。此外,脑、神经索以及神经连合内的神经细胞还向外发出纤维支配身体各部,调节各种器官和组织的功能。随着无脊椎动物的进化,在体内形成按体节成对分布的神经节,神经节之间有纵向的神经索和横向神经联合相连接。而后,一些头端神经节组合为脑,每个体节内的一对神经节也融合为一,由纵行腹面单一的神经索相串连,形成贯连首尾的腹神经索。更进一步的演化则使分散在各体节的神经节合并成为数不多的胸、腹、足和内脏等神经节。这种神经成分逐渐集中,使中枢神经系统与外周神经系统区分更加明显的过程叫做中枢化;在动物的尖端有越来越多的神经组织集中,并承担更为复杂的生理功能的过程,则被称为头部优势发育。

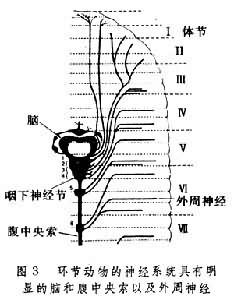
环节动物门 神经系统得到了进一步发展:脑内已经分化出与感觉有关的特殊部位,脑的外层由单极神经元组成,这些神经元的轴突在脑的中央部分构成神经堆,并通过轴突与轴突的接触构成脑细胞间的突触联系。环节动物的每一体节有一对神经节,神经节前后相串连,构成一对行走于体表之下的腹神经索。脑一般均位于咽的背面,向左右两侧下方发出环咽连接,与第一对腹神经节相连。有些环节动物每一体节两侧的神经节已经融合,因而成对的腹神经索也并成单一的腹中央索(图3)。
 在环节动物的脑和神经节内,有支配体壁肌肉的运动神经元,也有联系不同神经元的中间神经元,而感受外周环境刺激的神经细胞则位于皮肤之内或皮肤之下,形成外周神经网,这些神经元的向中轴突则进入腹神经节内。
在环节动物的脑和神经节内,有支配体壁肌肉的运动神经元,也有联系不同神经元的中间神经元,而感受外周环境刺激的神经细胞则位于皮肤之内或皮肤之下,形成外周神经网,这些神经元的向中轴突则进入腹神经节内。节肢动物门 低等节肢动物每个体节均有一对神经节,由纵行的腹神经索将同侧的神经节相连,同一体节的两个神经节之间有横向的连合。在咽的背面有脑,借助于环咽连接将脑与第一对腹神经节相连(图4)。
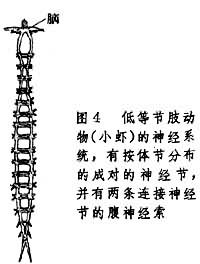 因此,节肢动物的神经系统基本上保持着环节动物神经系统的模式。节肢动物的外周神经也是由脑和神经节发出,体壁上已经没有外周神经网。高等节肢动物的神经系统有进一步集中的趋向,例如,蟹在身体前端按体节分布的神经节已经融合。作为腹神经索第一个神经节的咽下神经节,一般由身体前端 2对、3对或更多的神经节融合而成(图5)。
因此,节肢动物的神经系统基本上保持着环节动物神经系统的模式。节肢动物的外周神经也是由脑和神经节发出,体壁上已经没有外周神经网。高等节肢动物的神经系统有进一步集中的趋向,例如,蟹在身体前端按体节分布的神经节已经融合。作为腹神经索第一个神经节的咽下神经节,一般由身体前端 2对、3对或更多的神经节融合而成(图5)。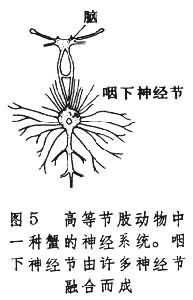 咽下神经节接受来自感觉器官的传入纤维,同时也发出纤维支配心脏、肌肉以及其他器官。脑的结构和功能都比环节动物复杂,一般分为3个主要区域:①原脑,包括一对视叶以及接受触角、各种感受器和其他脑部发来信息的整合中枢;②次脑,第一对触角发来信息的联合中枢;③后脑,发出纤维支配上段消化道以及上唇部位。节肢动物脑的复杂化还表现在它具有多种多样的神经元。根据脑神经元的大小,脑体的位置、轴突的走向和分枝情况,约可分为10多种。一些高等节肢动物脑内神经元可以多达10万,神经内分泌的功能也很发达。与此相应,昆虫外周感受器的分化程度也远比环节动物复杂。
咽下神经节接受来自感觉器官的传入纤维,同时也发出纤维支配心脏、肌肉以及其他器官。脑的结构和功能都比环节动物复杂,一般分为3个主要区域:①原脑,包括一对视叶以及接受触角、各种感受器和其他脑部发来信息的整合中枢;②次脑,第一对触角发来信息的联合中枢;③后脑,发出纤维支配上段消化道以及上唇部位。节肢动物脑的复杂化还表现在它具有多种多样的神经元。根据脑神经元的大小,脑体的位置、轴突的走向和分枝情况,约可分为10多种。一些高等节肢动物脑内神经元可以多达10万,神经内分泌的功能也很发达。与此相应,昆虫外周感受器的分化程度也远比环节动物复杂。软体动物门 由于生活习性和行为相差很大,不同纲属动物的神经系统的结构及复杂程度也有很大差异。一些低等软体动物(如石决明)的神经系统仍然保持分节的结构(图6),
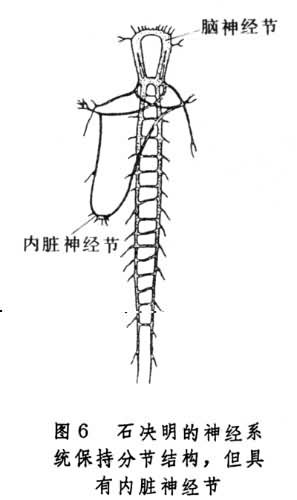 但是多数成体的神经系统,已没有明显的分节现象。中枢神经系统一般都由 6对分界明显的神经节组成。各神经节之间由纵向的神经链和横向的联合所连接。高等软体动物(如头足类的乌贼)中枢神经系统的神经节进一步集中,围聚在食管的周围,形成了脑。脑的附近由神经节形成了许多叶。此外,还有一些星状神经节分布在外周,调节身体各部分的功能。在脑的内部神经网中还有许多神经细胞集聚而成的小岛,相当脊椎动物脑内的神经核团。软体动物的神经细胞多是单极的,但头足类脑内部还有多极神经元。它们的感觉器官也有进一步的发展。这种中枢神经系统结构的进化以及感觉器官的形成,使这类动物活动灵便,并可猎食。
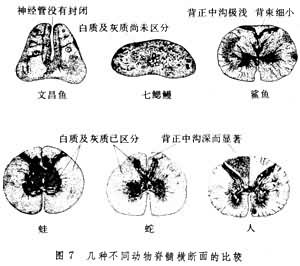
但是多数成体的神经系统,已没有明显的分节现象。中枢神经系统一般都由 6对分界明显的神经节组成。各神经节之间由纵向的神经链和横向的联合所连接。高等软体动物(如头足类的乌贼)中枢神经系统的神经节进一步集中,围聚在食管的周围,形成了脑。脑的附近由神经节形成了许多叶。此外,还有一些星状神经节分布在外周,调节身体各部分的功能。在脑的内部神经网中还有许多神经细胞集聚而成的小岛,相当脊椎动物脑内的神经核团。软体动物的神经细胞多是单极的,但头足类脑内部还有多极神经元。它们的感觉器官也有进一步的发展。这种中枢神经系统结构的进化以及感觉器官的形成,使这类动物活动灵便,并可猎食。原始脊索动物 原始脊索动物的胚胎早期,由于外胚层内褶在脊索上方形成一个中空的背神经索,后者发展成为中枢神经系统,这是重要神经胚的形成过程。此外,视觉、嗅觉、味觉以及听觉等主要感觉器官都有集中在身体前端的趋势,出现头部发育优势。这种向脊椎动物演化的痕迹可以在头索动物亚门的文昌鱼的神经系统中看到。文昌鱼的中枢神经系统是它的背神经索,是原始的脊髓。由于胚胎时期外胚层内陷的神经褶皱还没有愈合,因而脊髓的中央管尚未形成,整个脊髓背面存留一条明显的槽缝。脊髓内的神经元分散,没有白质和灰质的区分(图7)。
 由于头部还没有形成真正的脑,因而脊髓内也未形成专门的传导束。脊髓的背面发出成对的脊神经,叫做背侧脊神经,按节段分出。背脊神经中的纤维都不具髓鞘。这些纤维既包括躯体感觉,也包括内脏感觉和内脏运动纤维,所以文昌鱼没有脊神经节,一切感觉神经元的胞体均位于脊髓之内;没有自主神经系统与躯体神经系统之分,由脊髓神经元发出的支配内脏的神经直接终结在消化道上,缺少内脏神经节。此外,位于背部分成节段的特殊肌肉纤维直接与脊髓表面相接触,在那里与运动神经元构成联系,形成运动终板,接受支配。因此,文昌鱼的脊神经也没有背根和腹根之分。文昌鱼的脊椎虽然明显分节,但椎骨上方仍然没有形成神经弓,因而脊髓仍然不位于椎管之内。这一切都表明,这是一种较为原始的神经系统。
由于头部还没有形成真正的脑,因而脊髓内也未形成专门的传导束。脊髓的背面发出成对的脊神经,叫做背侧脊神经,按节段分出。背脊神经中的纤维都不具髓鞘。这些纤维既包括躯体感觉,也包括内脏感觉和内脏运动纤维,所以文昌鱼没有脊神经节,一切感觉神经元的胞体均位于脊髓之内;没有自主神经系统与躯体神经系统之分,由脊髓神经元发出的支配内脏的神经直接终结在消化道上,缺少内脏神经节。此外,位于背部分成节段的特殊肌肉纤维直接与脊髓表面相接触,在那里与运动神经元构成联系,形成运动终板,接受支配。因此,文昌鱼的脊神经也没有背根和腹根之分。文昌鱼的脊椎虽然明显分节,但椎骨上方仍然没有形成神经弓,因而脊髓仍然不位于椎管之内。这一切都表明,这是一种较为原始的神经系统。脊椎动物 脊椎动物神经系统的进化表现在 3个重要方面:在神经系统中枢化进一步发展的同时,外周神经系统也得到充分的发展,出现了自主神经系统头部优势发育更为明显,同时大脑的结构和功能更为复杂和完善;由于椎骨神经弓的形成,脊髓均位于骨质的椎管内,而脑则位于颅骨腔中。
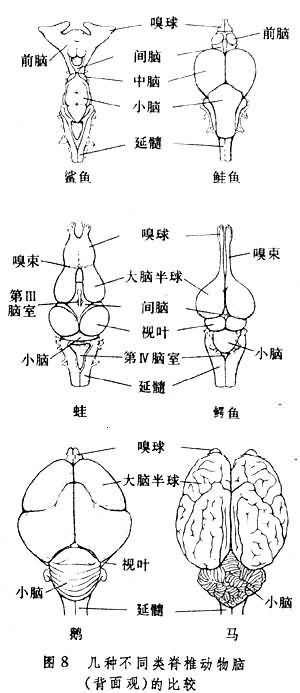
圆口鱼纲 头部优势发育已较原始脊索动物明显,脊髓前端的脑位于颅腔内,但是发育很差。大脑半球被挤压在嗅球后方,很不显著,表面只是一层简单的神经结构,被称为皮质。间脑的发育也处于萌芽阶段,只是在上方的顶器,松果腺和缰比较明显。视叶很小,延髓相对地比较发达。由于这类动物少动及营寄生活,因而小脑也不发达。脊髓有了进一步的发展,但是灰质和白质之间的界线仍不明显(图8)。
 以七鳃鳗为例,在脊椎节段之间发出成对的背侧脊神经和腹侧脊神经。一部分的感觉神经元的胞体仍然位于脊髓内,但是另一部分感觉神经元已经集聚在脊髓之外,形成背侧神经节。此外,与每一侧背侧脊神经相对应,每一脊髓节段都具有腹侧脊神经。后者主要由支配躯体的运动神经纤维所组成。但是,腹侧脊神经与背侧脊神经并没有合并,而是分别独立地走向它们所支配的效应器和感受器,并且在背侧和腹侧脊神经中都混合有一些支配内脏的运动纤维。圆口鱼纲动物的中枢神经系统内,神经胶质细胞数量很少,并且很少分化,是以一种原始的形式出现。
以七鳃鳗为例,在脊椎节段之间发出成对的背侧脊神经和腹侧脊神经。一部分的感觉神经元的胞体仍然位于脊髓内,但是另一部分感觉神经元已经集聚在脊髓之外,形成背侧神经节。此外,与每一侧背侧脊神经相对应,每一脊髓节段都具有腹侧脊神经。后者主要由支配躯体的运动神经纤维所组成。但是,腹侧脊神经与背侧脊神经并没有合并,而是分别独立地走向它们所支配的效应器和感受器,并且在背侧和腹侧脊神经中都混合有一些支配内脏的运动纤维。圆口鱼纲动物的中枢神经系统内,神经胶质细胞数量很少,并且很少分化,是以一种原始的形式出现。软骨鱼纲 大脑比圆口鱼纲发达,一对大的嗅球分得很开,各自与侧脑室相接。两侧大脑半球之间的联合广泛,中间隔着第三脑室。大脑半球的表面集聚着分层的细胞,可以分为古皮质和居于半球外侧的旧皮质,纹状体也很明显。除了松果腺和缰,间脑开始分化,可以明显地区分出丘脑与下丘脑。在丘脑后部出现了视觉转换核团外侧膝状体,使丘脑开始成为感觉信息中转和整合的重要中枢部位。下丘脑具有后叶以及一个腹侧与脑下垂体相连接的薄薄脉管囊,这两者都是鱼类下丘脑所特有的结构。中脑的视叶很发达包围着向后伸展的中脑脑室管,红核也开始出现。在一些快速游泳的鱼类(如鲨鱼),小脑发达,两半球融合,小脑皮层具有像高等脊椎动物一样的3层结构,并且可以区分出古小脑和旧小脑(图8)。在延髓中分化出橄榄核团以及网状结构。软骨鱼纲的中脑可以经过转换神经元与脊髓联系,而小脑已发出下行至脊髓的纤维,控制脊髓的运动神经元,这些结构的发展使动物的运动调节更为协调和完善。此外,在软骨鱼类的中枢神经系统中,胶质细胞已经有了分化,出现了星形胶质细胞、少突神经胶质细胞,甚至小神经胶质细胞,这和哺乳动物的情况十分近似,当然也是神经系统进化的一种表现。在圆口鱼类的神经系统中,神经纤维都不具髓鞘,但是软骨鱼类已经有完好的有鞘纤维。脊神经按节段分布,有背根、腹根之分,并且融合。感觉神经元已经聚集在脊神经节内。这也反映出软骨鱼类神经系统的进化。
硬骨鱼纲 神经系统有了进一步发展。属于端脑的3个主要部分,嗅球、纹状体和大脑皮质变化较大,不同种动物之间的差异也很明显。肺鱼端脑与软骨鱼类相近似,大脑半球表面外突,而辐鳍亚纲中的一些鱼类大脑半球的表面则向外侧翻出,所以形成中凹。由于皮质和纹状体靠近,显得皮质增厚,但分化程度仍然不高。硬骨鱼类的间脑较为发达,已经可以区分出上丘脑、下丘脑、腹侧丘脑以及丘脑等4个主要部分,其中以下丘脑最为明显,具有脉管囊。所有硬骨鱼类的视叶和小脑都相当发达(图8)。在一些鱼类,由小脑发出神经纤维,建立了和中脑之间的联系。延髓与味觉关系密切,并且与吞食运动有关。在有些鱼类延髓有一对巨大的茅斯纳尔氏细胞,向脊髓发出轴突控制动物的逃遁运动。脊髓中传导兴奋的纤维已经集聚,细胞在已经集中,因而有白质和灰质的区分。通常腹侧灰质柱更为明显。由于传导束和灰质核团的形成,脊髓随之膨大,并形成了脊髓背面和腹面的背正中沟和腹中央裂。按节段发出的脊神经背根和腹根已经汇合成为单一的脊神经。背根主要由传入的感觉纤维组成,腹根则由运动纤维组成。感觉神经元胞体位于背根神经节内。脊神经进一步分为背枝、腹枝和内脏枝发向躯体和内脏,基本上具有了在哺乳动物所看到的脊神经模式。
两栖纲 大脑分化程度不高,与硬骨鱼相比进化并不显著。端脑的两个半球分得较开(图8),中间隔着狭小的脑室。古皮质形成了原始的海马,旧皮质构成了梨状区的雏形。纹状体也很小。上丘脑和鱼类相似,只是松果腺较为发达(尤其是无尾目)。丘脑的体积增大,表明作为感觉信息的转换和整合的部位已经开始发展。下丘脑内开始出现乳头体。中脑的主要部分仍然是视叶,但视叶的体积相对地较鱼类为小。在中脑开始出现一些神经元,它们的轴突组成下行的顶盖脊髓束,这表明脑干部分对脊髓反射活动调节的加强。两栖类的小脑不甚发达,与之相应的是这类动物的运动能力较差。在脊髓的横断面上,白质与灰质的区分更为明显(图7)。在白质中,不同的上行和下行传导束比鱼类分化得更为明显。在灰质中已经出现不同的核团。运动神经元集聚在腹侧灰质柱,使后者变得更为发达。两栖类的自主神经系统亦较鱼类分化程度为高;具有较完整的交感神经系统和副交感神经系统。
爬行纲 大脑狭长,嗅球较小,但嗅束却比鱼类及两栖类长。前脑的纹状体及新皮质(neopallium)比两栖类发达,致使爬行类的大脑体积增大。更多的神经元在大脑表面集聚,但新皮层依然很薄。从大脑皮层的结构以及纹状体的体积和位置看来,爬行纲动物的前脑在发展上具有两种倾向:一类是趋向哺乳动物的大脑(如玳瑁),另一类倾向于鸟类(如鳄)。爬行类的背侧丘脑比两栖类复杂。松果腺发达,与顶眼之间具有神经纤维联系,其功能与对光反应的行为活动及内分泌调节有关。腹侧丘脑有了进一步发展,分化出许多神经核团,与哺乳动物腹侧丘脑的复杂程度十分近似。与前脑相比,视叶相对地较小,但仍然明显,这与大多数爬行动物具有良好视觉有关。此外,中脑出现黑质,红核也比两栖类的明显。小脑表面光滑、没有哺乳动物以及鸟类小脑表面所具的皱折,但在小脑内,一些神经元已经聚集成为核团。爬行类的延髓及脊髓结构更接近哺乳动物,外周神经系统也与哺乳动物类似。
鸟纲 脑相对地较爬行动物的大,分化的程度更高。由于鸟类在进化过程中的特殊位置,难以将其脑与哺乳动物脑的相应部分作比较,但是它的前脑纹状体比爬行类发达得多。尤其是古纹状体以及外纹状体明显增大,开始和丘脑发生了联系,接受由丘脑转换核团发来的听觉、视觉以及其他感觉信息。上纹状体的细胞分化与分层更为突出,被认为是新皮层的萌芽。鸟类的嗅球与嗅束比较小,但视神经及视交叉发达。背侧丘脑发展明显。居于大脑、小脑交界处外侧的视叶仍然很大,并具有分层结构,它接受来自一切感受器以及小脑的信息。除哺乳动物而外,鸟类小脑最为发达,表面具有回旋的皱褶(图8)。介于小脑中线处的新小脑开始发展,它与鸟类的飞行活动有关。由于小脑的发展,致使脑干下方出现脑桥,橄榄核团也进一步分化和增大。爬行动物脊髓的灰质与白质界线分明。白质由上行和下行的许多神经纤维束组成。灰质内也分化出不同的神经核团。脊髓的每一节段向两侧各分出背根与腹根一对。背根全由感觉神经组成,腹根一般为躯体运动神经及内脏运动神经纤维组成。它们在脊柱内联合,然后再分枝走向与节段相应的所支配的器官。所以,爬行动物的脊髓结构和哺乳动物更为相似。
哺乳纲 神经系统得到更为充分的发展,与之相应的功能活动也更为复杂,高等动物的学习、记忆以及人类的语言和抽象思维等高级功能活动,都是神经系统进化和发展的结果。(见中枢神经系统、外周神经系统)
神经系统的发生 在高等动物和人的胚胎发育过程中,神经系统也经历一种由简单到复杂的过程,这过程犹如种系发生各阶段神经系统进化过程的重演。
人胚胎发育至第3周时,外胚层开始出现称为神经板(图9)
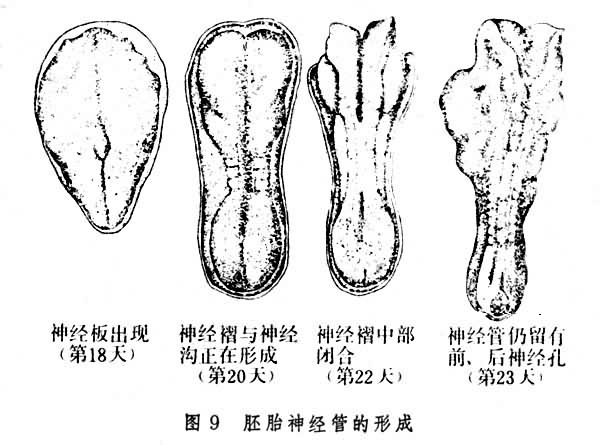 的区域。神经板的内侧微微增厚,中间部分下陷,出现一个浅槽,形成神经沟。第3 周末,神经板增厚部分的边缘渐渐隆起,形成神经褶。在神经沟不断加深的同时,两侧神经褶的边缘向内侧合拢,这时神经板的中段开始出现分节。第4周时,分节不断增多,两侧神经褶开始在第四体节处连接融合,这即是形成神经管的开始。接着,神经褶闭合过程向神经板的两端扩展,最后形成两端开孔的神经管。由于神经板两端发育不同,神经管的头端部分变得更为宽大,将来发育成脑,而较为狭窄的尾端部分伸延生长得较快,则发育成脊髓。当出现第20体节时,神经管的前端开孔封闭。稍后,在第26体节形成时,尾孔闭合。这时,神经管壁的细胞增生加快,每8小时,数量约增加1倍。快速的细胞增生不仅使神经管伸长,而且引起神经管发生弯曲,这种弯曲在头端尤为明显(图10)。
的区域。神经板的内侧微微增厚,中间部分下陷,出现一个浅槽,形成神经沟。第3 周末,神经板增厚部分的边缘渐渐隆起,形成神经褶。在神经沟不断加深的同时,两侧神经褶的边缘向内侧合拢,这时神经板的中段开始出现分节。第4周时,分节不断增多,两侧神经褶开始在第四体节处连接融合,这即是形成神经管的开始。接着,神经褶闭合过程向神经板的两端扩展,最后形成两端开孔的神经管。由于神经板两端发育不同,神经管的头端部分变得更为宽大,将来发育成脑,而较为狭窄的尾端部分伸延生长得较快,则发育成脊髓。当出现第20体节时,神经管的前端开孔封闭。稍后,在第26体节形成时,尾孔闭合。这时,神经管壁的细胞增生加快,每8小时,数量约增加1倍。快速的细胞增生不仅使神经管伸长,而且引起神经管发生弯曲,这种弯曲在头端尤为明显(图10)。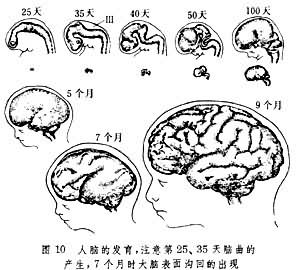 第4周时,神经管的头端形成了前、中、后3个脑泡。由于脑壁各部分发育不均,相继出现了3个脑曲:第Ⅰ脑曲凸向背方,位于中脑,亦称中脑曲;第Ⅱ脑曲发生在后脑与脊髓交界处,也凸向背方,称为颈曲;约在第六周时,出现第Ⅲ脑曲,称为脑桥曲(图10)。脑曲出现的同时,脑的各部分分化加快。第五周时,胚胎的脑已初步形成5个部分:前脑泡发育成端脑和间脑;中脑泡发育为中脑(mesencephalon);后脑泡即为菱脑(rhombencephalon),以后发展成为小脑、脑桥及延髓。
第4周时,神经管的头端形成了前、中、后3个脑泡。由于脑壁各部分发育不均,相继出现了3个脑曲:第Ⅰ脑曲凸向背方,位于中脑,亦称中脑曲;第Ⅱ脑曲发生在后脑与脊髓交界处,也凸向背方,称为颈曲;约在第六周时,出现第Ⅲ脑曲,称为脑桥曲(图10)。脑曲出现的同时,脑的各部分分化加快。第五周时,胚胎的脑已初步形成5个部分:前脑泡发育成端脑和间脑;中脑泡发育为中脑(mesencephalon);后脑泡即为菱脑(rhombencephalon),以后发展成为小脑、脑桥及延髓。端脑的发育 胚胎在第4周之后,前脑泡壁向两侧突出形成两个脑泡,然后发展成为两侧的大脑半球。半球内的空腔即为左右大脑侧室,并借室间孔与间脑的第三脑室相通。第6周时,大脑半球的底壁细胞增生迅速,使室壁增厚,形成纹状体,左右半球的前部腹侧各向前凸出一盲管,而后发展成为嗅球。与此同时,大脑半球前部的皮质很快发展成为额叶,上部发展成为顶叶,后部发展成为枕叶及颞叶。第 5个月的人胚胎大脑半球表面仍然光滑,第7个月时半球表面方出现主要沟、回。此时,发达的大脑半球已将间脑、中脑、脑桥、小脑及延髓掩盖(图10)。
在人的大脑半球上方,是具有6层结构的新皮层,它占据成年人整个大脑皮层表面的94%,亦被称为均匀皮层(见大脑)。这部分皮层在胚胎发育过程中形成较迟。半球其余表面结构包括古皮层及旧皮层,在神经系统发生中出现较早,这些部分统称为非均匀皮层,将来发展成为边缘系统的主要部分。新皮层是由端脑泡的假分层上皮演发而成。最初,假分层上皮中靠近脑室处的椭圆形细胞进行有丝分裂,而后位于其上方的细胞也进行分裂。分裂出的子细胞核并不与母细胞断离,而是随着母细胞的胞浆向大脑表面引延而迁移,然后再和母细胞的胞浆断离,变为成神经细胞。第7周的人胚上,这些迁移的成神经细胞聚集成和脑表面平行的层次,形成皮层板。10周时,这种迁移告一段落,在皮层板和脑室壁之间出现大量的神经纤维,使脑壁增厚。在第13~15周时,脑室壁发生第二次细胞的迁移,分裂的大多是位于假分层上皮上方的细胞。分裂出的不成熟成神经细胞,在迁向皮层板的途中需要胶质细胞的引导,后者具有和大脑表面垂直长的突起。新生的成神经细胞就沿着这些突起,穿越加厚的端脑壁,上升到脑的表面。后迁移的细胞更接近脑的表层。这些成神经细胞一般排列成为柱状,在感觉区皮层柱状排列尤为明显。成神经细胞的迁移使皮层板进一步加厚,并使它和相邻的区域明显地区别开来。18周后,脑室壁的假分层上皮细胞接近耗竭,变成了很薄的一层,这时上述细胞的迁移也告终止。由于成神经细胞的分化,皮层板出现了明显的层次,在这基础上,进一步形成具有分子层、外颗粒层、外锥体细胞层、内颗粒层、锥体细胞层和梭形细胞层等6层结构的新皮层。胚胎第6周,大脑的底壁细胞增生,形成纹状体原基。第10周时纹状体进一步增大,此时,由于神经纤维的穿行,遂将其分为两个部分:居于腹外侧的豆状核和居于背内侧方的尾状核。而行走于这两个核团之间的纤维即发展成为内囊。因此,豆状核、尾核及内囊统属于端脑部分。
丘脑的发育 从发生上看,纹状体原基和第Ⅲ脑室壁极为接近,后者是发展成为间脑的主要基础。由于纹状体原基细胞有丝分裂很旺盛,有些细胞即迁移至间脑背部,参与缰的形成,但是形成丘脑的主要基础仍然是第Ⅲ脑室壁的细胞。在神经管形成的早期,它的壁为一层柱状上皮,胚胎发育到6周时,管壁明显分为3层:内层为室管膜层;室管膜细胞不断分裂增生并迁移至外侧形成中层,称为外套层。外套层细胞不断分化成为神经细胞及神经胶质细胞。神经管的外层为边缘层,由室管膜层及外套层细胞的突起构成。由于外套层细胞迅速增生和分化,在神经管的左右两壁各出现两个增厚部分:背侧的称为翼板,腹侧的为基板。两板之间各形成了一条纵贯神经管前后的界沟。在间脑部分,侧壁的界沟并不明显,只留有丘脑下沟的痕迹,其上方即为上丘脑,是来自翼板和脑室的顶板,而丘脑则完全由翼板演化而成,下丘脑则由基板和脑室的底板共同形成。
中脑的发育 中脑内的界沟比较清楚,脑壁发育较厚。翼板部分主要形成顶盖,由于分裂后的成神经细胞迁移到背方,从而形成具有分层结构的上丘和下丘。翼板中的部分分裂细胞迁移到中脑的腹侧形成红核。中脑的基板部分分化形成第三和第四脑神经。基板的成神经细胞也分化成为被盖部分的网状结构。
小脑和脑桥的发育 小脑和脑桥主要由菱脑的后脑(metencephalon) 部分发育而成。在早期的后脑横切面上,可以看到扩大的第4脑室──菱形窝,其上方为宽阔的顶板,因而翼板与基板向两侧展开(图11)。
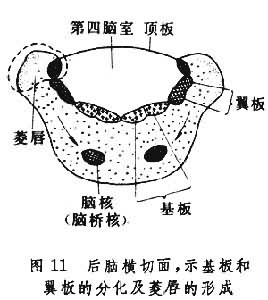 到第7周末,神经管在后脑处发生向腹侧方下凸的脑桥曲,此时顶板与翼板的连接处逐渐增厚,形成菱唇,菱唇由于脑桥曲的形成和深陷而变为横向排列的小脑板。小脑板的两侧较厚,中间一狭窄部分较薄,前者发育成为小脑半球,后者发育成为蚓部。这是小脑发育的第一阶段。在16周的胚胎,表面光滑的小脑板上出现横向的裂隙,将小脑本体和绒球分开。两侧的绒球由中间的小脑小结连接,构成了绒球小结叶,这是种系发生过程中小脑最古老的部分。然后,在小脑本体上出现横向裂隙,将小脑分为前叶、后叶。稍迟,在前叶、后叶上又出现裂隙,再将小脑分为许多叶。由于小脑的发生,菱形窝上方的顶板头侧部分变成一窄带,是为前髓帆,顶板的中间部分和小脑底部融合,顶板的尾侧部分形成了后髓帆。翼板和基板的细胞分裂增生,迁移至菱形窝的底部形成了许多运动和感觉神经核团。此外,还由于网状结构的出现以及小脑的大量传出、传入纤维,以及穿行在菱脑窝底部的上行、下行神经纤维的存在,使得这里特别增厚,形成脑桥。
到第7周末,神经管在后脑处发生向腹侧方下凸的脑桥曲,此时顶板与翼板的连接处逐渐增厚,形成菱唇,菱唇由于脑桥曲的形成和深陷而变为横向排列的小脑板。小脑板的两侧较厚,中间一狭窄部分较薄,前者发育成为小脑半球,后者发育成为蚓部。这是小脑发育的第一阶段。在16周的胚胎,表面光滑的小脑板上出现横向的裂隙,将小脑本体和绒球分开。两侧的绒球由中间的小脑小结连接,构成了绒球小结叶,这是种系发生过程中小脑最古老的部分。然后,在小脑本体上出现横向裂隙,将小脑分为前叶、后叶。稍迟,在前叶、后叶上又出现裂隙,再将小脑分为许多叶。由于小脑的发生,菱形窝上方的顶板头侧部分变成一窄带,是为前髓帆,顶板的中间部分和小脑底部融合,顶板的尾侧部分形成了后髓帆。翼板和基板的细胞分裂增生,迁移至菱形窝的底部形成了许多运动和感觉神经核团。此外,还由于网状结构的出现以及小脑的大量传出、传入纤维,以及穿行在菱脑窝底部的上行、下行神经纤维的存在,使得这里特别增厚,形成脑桥。延髓的发育 延髓由胚胎的后脑泡,即菱脑的末脑部分发育而成,头侧与脑桥相连接,尾侧和延髓连续,为第四脑室后半部的底部。在胚胎早期,后脑泡由于脑曲的出现,以及脑室壁的外翻而形成开阔的菱形窝,其底部翼板及基板部分均可辨认(图12),
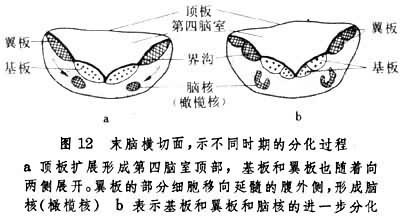 翼板与基板之间是为界沟。和脑桥的发育过程一样,延髓的感觉神经核团及中继核团是由翼板发展而来,如孤束核,橄榄核,而运动性神经核团,如下涎核等,则由基板发育而成。第四脑室的顶板与间充质细胞及毛细血管一起组成了脉络丛突入在脑室上方。
翼板与基板之间是为界沟。和脑桥的发育过程一样,延髓的感觉神经核团及中继核团是由翼板发展而来,如孤束核,橄榄核,而运动性神经核团,如下涎核等,则由基板发育而成。第四脑室的顶板与间充质细胞及毛细血管一起组成了脉络丛突入在脑室上方。脊髓的发育 脊髓系由末脑的尾侧神经管演发而成。在6周的胚胎上,这部分的神经管壁基本上是一个3层结构,两侧翼板、基板及界沟明显(图13)。
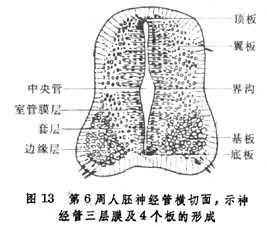 由于两边基板的细胞增生,使神经管的腹侧特别膨大,在3个月的人胚脊髓断面上,已出现明显的腹正中裂。两侧的翼板由于细胞增生,向内侧挤压,结果使背侧中央管壁合拢,形成背正中隔,中央管的管腔也因之变得细小。在背侧的外表面正中线上出现一条纵行浅沟,称为背正中沟(图14)。
由于两边基板的细胞增生,使神经管的腹侧特别膨大,在3个月的人胚脊髓断面上,已出现明显的腹正中裂。两侧的翼板由于细胞增生,向内侧挤压,结果使背侧中央管壁合拢,形成背正中隔,中央管的管腔也因之变得细小。在背侧的外表面正中线上出现一条纵行浅沟,称为背正中沟(图14)。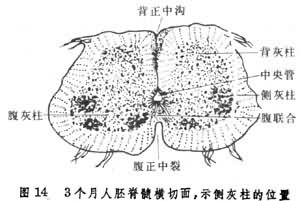 此时白质与灰质的界线已经分明。脊髓的背侧灰柱是由翼板的细胞增生、分化而成,而前侧灰柱则由基板发展而成。左右两侧灰质在中央管周围并相互连接,在脊髓的横切面上构成蝶样的图形。由基板发育而成的前侧灰柱,主要为运动神经元构成,而来自翼板的背侧灰柱则含有联合神经元,接受来自外周的传入神经冲动。此外,在胸段和腰段的灰质两侧,各发生一个侧灰柱,内为自主性神经元,它们发出节前纤维支配内脏器官(见自主神经系统)。脊髓的尾端发育较特殊,它不是神经板演发的产物,而是由其末端的尾结节发育而成。尾结节分裂出成束的细胞,称为终丝。终丝的末端有一个膨大的吸盘状结构,固着在尾骨附近的皮肤下方。
此时白质与灰质的界线已经分明。脊髓的背侧灰柱是由翼板的细胞增生、分化而成,而前侧灰柱则由基板发展而成。左右两侧灰质在中央管周围并相互连接,在脊髓的横切面上构成蝶样的图形。由基板发育而成的前侧灰柱,主要为运动神经元构成,而来自翼板的背侧灰柱则含有联合神经元,接受来自外周的传入神经冲动。此外,在胸段和腰段的灰质两侧,各发生一个侧灰柱,内为自主性神经元,它们发出节前纤维支配内脏器官(见自主神经系统)。脊髓的尾端发育较特殊,它不是神经板演发的产物,而是由其末端的尾结节发育而成。尾结节分裂出成束的细胞,称为终丝。终丝的末端有一个膨大的吸盘状结构,固着在尾骨附近的皮肤下方。脑神经节和脊神经节的发生 胚胎早期,当神经板内侧两边增厚形成神经褶及神经管时,两侧神经褶边缘的上皮细胞遂分化成为神经嵴原基(图15)。
 神经嵴细胞在皮肤与神经管之间形成一板状结构。而后,由于细胞的增生和迁移,在神经管的两旁,形成一些不相连续的细胞团,进一步发展成为脊神经节和脑神经节。此外,自主神经系统的神经节、肾上腺髓质的嗜铬细胞以及外周神经干中的许旺氏细胞也是由神经嵴细胞迁移分化而成。
神经嵴细胞在皮肤与神经管之间形成一板状结构。而后,由于细胞的增生和迁移,在神经管的两旁,形成一些不相连续的细胞团,进一步发展成为脊神经节和脑神经节。此外,自主神经系统的神经节、肾上腺髓质的嗜铬细胞以及外周神经干中的许旺氏细胞也是由神经嵴细胞迁移分化而成。