真菌(卷名:生物学)
fungus, eumycetes
━━━━━━━━━━━━━━━━━━━━━━━━
简史
结构
细胞
隔膜
菌丝的变态
组织
营养和生长
营养成分
营养方式
生长
生殖
无性生殖
有性生殖
准性生殖
核循环
生态
腐生真菌及其习性
土壤真菌
水生真菌
嗜热真菌
喜冷真菌
大气真菌
寄生真菌及其寄生
植物寄生真菌
动物和人的寄生真菌
真菌寄生真菌
共生真菌及其伙伴
起源和演化
分类
意义━━━━━━━━━━━━━━━━━━━━━━━━
具有真正细胞核,能产生孢子而没有叶绿素,一般都能进行有性和无性繁殖,常具分枝的丝状营养体。种属很多,已报道的属达1万以上,种超过10万个。真菌一词的拉丁文(fungus)原意蘑菇。
简 史
真菌是一个古老的生物类群,在泥盆纪(4亿多年前)的第一批化石植物中,就有腐生和寄生的真菌。中国古代文献记载有黄帝与歧伯讨论醪醴(即酒)、夏代仪狄酿酒、周代杜康制酒的传说,从龙山文化出土的陶器中有盛酒的樽、冲酒的盉、煮酒的斝,这表明中国在新石器时期已经有了酒。宋代陈仁玉所著《菌谱》(1250)描述了食用菌11种,明代潘之恒所著《广菌谱》(1500)描述了食用菌19种。1977年在浙江余姚县河姆渡村进行考古发掘,出土物中就有菌类,这表明在仰韶文化时期(距今6000~7000年),中国人已经采食蘑菇。
在1700~1850年间,各国科学家广泛开展对大型真菌的研究,意大利的P.A.米凯利、荷兰的D.C.H.佩尔松、瑞典的E.M.弗里斯家族、英国的M.J.伯克利、捷克的C.J.柯尔达、法国的L.R.蒂拉纳兄弟等,都对真菌学的发展有重大的贡献。
1851~1950年间是近代真菌学全面发展的时期。德国植物学家H.A.德巴里由于对真菌的系统发育、多型性和生活史的精湛研究而被誉为近代真菌学的奠基人。意大利P.A.萨卡尔多的巨著《真菌汇刊》是这一时期对真菌分类学的巨大贡献。
20世纪50年代以来,现代真菌学飞速发展,如:①研究真菌的细微结构;②应用新技术和新方法;③医学真菌飞跃发展;④出现真菌毒素学;⑤抗癌真菌药物受到重视;⑥创立分子真菌学;⑦发展食用真菌,提出健康食品的名称;⑧改变生物分类法,将真菌独立成界等。
20世纪初中国用现代方法研究真菌,1915年胡先骕发表《菌类鉴别法》,为中国真菌鉴定打下了基础。戴芳澜于1927年出版《江苏真菌名录》,于1930年发表南京钓丝壳新种,是中国人用现代方法研究中国真菌的第一个新种。1939年邓叔群出版《中国高等真菌志》;1951年王云章出版《中国锈菌索引》。此外,还有不少植物学家、植物病理学家、工业真菌学家和医学真菌学家,也从事一些有关真菌学的研究。1980年成立中国真菌学会,1982年创刊中国《真菌学报》,现已开始筹编中国真菌志。
结 构
真菌是多型性的生物。它是以吸收水分和养料进行营养增殖的菌体,由简单到复杂,一般可分为原质团、单细胞、假菌丝、两型菌丝和菌丝体。菌丝体是常见的典型性营养体,由微小的丝状物构成,它在基物上向一个方向分枝、延伸以便获取养料。
细胞 菌丝细胞的四周包有透明的薄膜。细胞壁的里面充满原生质体以及浸没在原生质体中的各种细胞器。细胞壁为多层薄片结构,主要化学成分是多糖,另外含有蛋白质、类脂以及其他物质。在多数真菌的细胞壁中最具特征性的是甲壳质,其次是纤维素。细胞壁的内侧是由蛋白质和类脂组成的连续结构质膜。位于质膜与细胞壁之间的须边体,在真菌中也比在其他生物中更为常见。
在质膜包裹着的细胞质中,常见的真菌细胞器有:细胞核、线粒体、微体、核糖体、液泡、溶酶体、泡囊、内质网、微管、鞭毛等;常见的内含物有肝糖、晶体、脂体等;沃罗宁体和脂体做超薄切片不易看见,而是实际上存在的一层薄膜。细胞核有核膜、核膜为双层结构,并具有特征性的核膜孔。细胞核常含有一个大部分为RNA所组成的核仁,当核分裂时,核仁有时消失。高尔基器在真菌中不经常出现,至少不以典型的形式出现。
隔膜 大多数丝状真菌的原生质是由横隔膜隔开的,这种隔膜叫做分隔或隔膜。低等丝状真菌的隔膜在繁殖器官的基部形成,繁茂的生长菌丝无隔多核或是无隔。如果菌丝变老时,隔膜可随处形成。菌丝死亡时,原生质就移向生长点,随即形成隔膜,将死的部分与活的部分分开,因此,真菌的原生质不死。
隔膜是由菌丝内壁向内生长而成,有的继续生长,直至变成完整的隔片(封闭隔);有的发育不全,常留有一个孔(单孔隔)或多个孔(多孔隔),容许包括细胞核在内的一切细胞器通过,菌丝细胞变老或受伤时,旁边的伏鲁宁体和蛋白质晶体便可进行堵塞。多数担子菌的隔膜有一中心孔,孔的边缘膨大使中心孔成为琵琶桶状的特殊结构,外面覆盖一层弧形的膜,即为桶孔覆垫,覆垫有孔或管,由内质网形成。这种隔膜称为桶孔隔膜。真菌隔膜的形态、结构、功能和性质涉及真菌核移机制和真菌细胞的基本概念,并可能有助于系统发育学说的发展。
菌丝的变态 在长期的自然选择下,真菌的营养菌丝发生多种变态,从而更有效地获取养料,以满足生长发育的需要。常见的变态菌丝有吸器、压力胞、菌网和菌套等。前两者多在植物寄生菌中形成,后两者多在捕食线虫等的真菌中出现。
组织 有些真菌在发育的某个阶段,菌丝体变成比较疏松的或紧密交织的组织,这与通常组成菌体的疏松菌丝不同,叫做密丝组织。常见的密丝组织有长轴组织和拟薄壁组织两种类型:前者颇为疏松,构成此种组织的菌丝大致平行排列,其典型的长形细胞容易识别;后者排列紧密,常是等径或卵形,类似管束植物的薄壁细胞。这两种组织构成许多真菌不同类型的营养结构,如菌索、菌核和子座等。
菌索是由菌丝体平行排列组成的长形索状物,周围有外皮,尖端为生长点,有帮助真菌运送物质和蔓延侵染的功能,而在不适环境下呈休眠状态。菌核是坚硬的、能抵抗不良环境的休眠体,外壁由深色厚壁细胞构成,内层为浅色拟薄壁细胞。不同的真菌形成不同的菌核(如药用的茯苓、猪苓、雷丸和麦角等)。它可以长期休眠,在适宜条件下重新萌发,产生子实体、菌丝和分生孢子。子座是坚实的营养结构,有时由菌丝和寄主组织混合构成,它很像座垫或床座,常常在座上或座中形成子实体。
营养和生长
真菌对营养的需要与动物和细菌相似,而不像绿色植物那样能够自己合成食物。
营养成分 真菌能产生各种水解酶,将糖类、淀粉、纤维素、木质素等碳水化合物以及蛋白质和脂肪分解,用作食物。大多数真菌能利用无机或有机氮以及各种矿物元素来合成自己的蛋白质。实验研究表明:绝大多数真菌都需要碳、氢、氧、氮、磷、钾、镁、硫、硼、锰、铜、钼、铁和锌,可能还需要钙。许多真菌需要硫胺素和生物素,有些真菌需要吡哆醇和肌醇,但还没有发现需要从食物中吸取核黄素的真菌。一般来说,葡萄糖是真菌最好的碳源,有机氮化合物是真菌最好的氮源。真菌的食物以植物性的物质为主,其次是动物性的物质。
营养方式 真菌在生活中所需要的有机物质都依赖于自然界的其他生物。从死有机体中吸取养料的真菌叫做腐生菌。这些养料一般称为基物或基质。能侵害活有机体,而不能生活在死有机体上的真菌叫做绝对寄生菌。被寄生菌侵害的活有机体叫做寄主。近几十年,人们已经开始设计能生长绝对寄生菌的合成培养基,例如一些锈菌能在人工培养基上生长并完成它们的生活史。寄生和腐生并不是绝对的,在一定条件下,一些真菌既能侵害活有机体又能生活在死有机体上,这种真菌叫做兼性寄生菌或兼性腐生菌。有许多真菌一方面从其他活有机体摄取养料,一方面又向同一活有机体提供养料或好处,这是一种共生现象,具有共生关系的真菌叫做共生菌。
生长 主要指营养性菌体体积的加大和数量的增多。单细胞真菌菌体的生长,主要是经过细胞膨大、细胞核分裂、细胞质合成,最后达到细胞的芽殖或裂殖,进入无性繁殖。丝状真菌的生长是以顶端延长的方式进行。菌丝顶部是菌丝体的生长点,长50~100微米,其中含有大量的泡囊、壳质体和细胞核等。泡囊以水泡状形式从内质网转移到高尔基器,在那里浓缩、加工,并把它们的类内质网膜转化为类质膜,然后从分散高尔基器分泌释放,移至菌丝顶部,与质膜融合,使菌丝顶部的质膜增加。同时,顶部泡囊的内含物也被用来合成细胞壁,这些内含物含有壁的溶解酶类、壁的合成酶类、以及一些合成细胞壁的前体物。泡囊具有 3种作用:①运输细胞壁溶解酶去破坏原来壁组成之间的键;②运输新的壁物质,在壁合成酶的作用下,将它并入细胞壁中;③在生长期间增加质膜的表面积。壳质体是比泡囊小得多的微泡囊结构,是运输甲壳质合成酶的容器,为菌丝的伸长创造条件。在逐渐硬化的细胞壁和逐渐扩大的液泡的压力下,菌丝中活跃的原生质从衰老部分流向顶端,使菌丝顶端不断向前伸长。菌丝的这种生长方式叫做菌丝顶端生长。
生 殖
具有种的典型特征的新个体的形成。真菌的繁殖器官由营养体产生或转变而来。有些真菌在形成繁殖器官时,整个菌体转变成一个至多个繁殖结构,这叫做整体产果式,这种结构分化少,比较原始。大多数真菌的繁殖器官只是由菌体的一部分构成,其余部分继续维持正常的营养活动,这叫做分体造果式。真菌的繁殖包括无性生殖、有性生殖和准性生殖。
无性生殖 菌的无性生殖是指不经过两性细胞的结合便产生新的个体。生殖方法有:①体细胞(菌丝)的断裂;②体细胞分裂成子细胞;③体细胞或孢子的芽殖;④各种无性孢子(如游动孢子、孢囊孢子、分生孢子、厚垣孢子等)的产生,每个孢子可萌生芽管,再形成菌丝体。无性生殖在真菌的繁衍和传播上起重要作用。
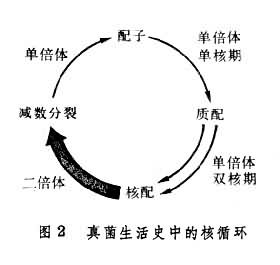
有性生殖 菌的两个可亲和性细胞核的结合。这种核的结合是通过游动配子配合、配子囊接触交配、配子囊交配、性孢子配合和体细胞配合来实现的。典型的有性生殖过程包括 3个明显不同的阶段:第一阶段叫质配,即两个带核的原生质体在同一细胞内互相配合;第二阶段叫核配,即上述细胞里两核的配合。有的真菌在质配后立即进行核配。另一些真菌不是立即进行核配,因此出现一个双核阶段,即每个细胞里有两个没有结合的核,细胞生长或分裂时,它们同时分裂,这是真菌特有的现象。这两个核一直要到相当晚的时候才能配合。核配后或迟或早会发生减数分裂,重新使染色体的数目减为单倍,这就是有性生殖的第三阶段。
真菌的有性生殖最后形成各种有性孢子,如休眠孢子囊、卵孢子、接合孢子、子囊孢子和担孢子。这些孢子是许多真菌用以渡过不良环境的休眠体,也是许多植物病原真菌每次初侵染的传染体,对于真菌的复壮或形成杂种优势具有重要意义。同时,有性孢子也是真菌分类的重要依据。
根据有性生殖的亲和性,真菌可分为同宗配合和异宗配合两类:前者是每一菌体自身可孕,不靠其他菌体的帮助便可单独进行有性生殖;后者是每一菌体自身不孕,不管是否雌雄同体,都需要借助其他可亲和性菌体的相对交配型来进行有性生殖。异宗配合又有两极性和四极性之分。孤雌生殖在真菌中也时有发现。
准性生殖 1952年,美国的G.蓬泰科尔沃和罗珀在丝状真菌中发现的一种导致基因重组的机制。在这种机制中,遗传性的重组不是依赖有性生殖的减数分裂,而是依赖准性生殖的有丝分裂。
准性生殖的第一步是形成异核体。异核体菌丝的形成有3种途径:①异核菌丝的结合;②同质菌丝内的核发生突变;③菌丝内一些单倍体核进行核配形成二倍体核。准性生殖的第二步是两个核的融合,同质的和异质的核都可以核配,形成同质的二倍体核和异质的二倍体核,结果,这个阶段的菌丝细胞中最少含有5类核:两种单倍体核,两种纯合二倍体核,一种异合二倍体核。但是,自发形成异合二倍体核的频率非常低,如构巢曲霉在100万个菌株中才出现一个异合二倍体。准性生殖的第三步包括二倍体核一系列非典型的、不规则的单倍体化。由于有丝分裂后期姊妹染色单体未分开,或者由于一个染色体分裂为2以后都趋向一极,而造成子核的染色体数目不相等,产生非整倍体,第1个子核多一个染色体(2n+1),第2个子核又少一个染色体(2n-1)。 “2n+1”非整倍体又称三体,往往失去一个染色体而成为二倍体。在这个过程中,可以由杂合体变为纯合体。“2n-1”非整倍体又称单体,常在不断的有丝分裂中继续丧失染色体,直至成为单倍体,准性生殖全过程可概括为质配、核配和单倍体化(图1)。半知菌不发生有性生殖,发生准性生殖的也只有曲霉、青霉和镰孢霉属中的少数种。准性生殖在演化中具有一定的重要性。有性生殖和准性生殖并不互相排斥,有的真菌,如构巢曲霉,既发生有性生殖也发生准性生殖。
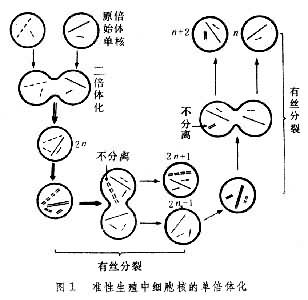
在异合二倍体繁殖的有丝分裂中,染色体之间偶尔也发生交换,形成新的组合和连锁(见连锁和交换)。有些新组合的二倍体核在形成分生孢子时进入孢子,而在孢子萌发时生出二倍体菌丝。这样的重组是准性生殖的主要方面,因为它使没有有性生殖的真菌也能得到有性生殖的益处。不过发生的频率很低,仅为有性生殖的1/500。
准性循环的遗传虽然具有有性生殖的内容,但与有性循环之间还有几点区别:①准性循环的过程类似有性循环,但不像有性循环那样固定,而且单倍体化的完成要牺牲大量的非整倍体;②在一个菌丝细胞内,异核现象和异合现象可以同时发生,不过异合机会极少,必须进一步分离才能发现。由于一个二倍体自发分离(体细胞交换和单倍体化)的频率很低,往往需要用各种化学的或物理的因子加以诱导,例如用对氟苯丙氨酸(PFA)诱导构巢曲霉单倍体化就有特别显著的效果,用氟尿嘧啶(FU)诱导构巢曲霉的有丝分裂交换(体细胞交换),效果也很明显。另外还有许多因子,如氮芥、甲醛和紫外线等,对构巢曲霉的单倍体化和有丝分裂交换都起诱导作用。
准性生殖的发现进一步深化了对真菌的认识。通过体细胞重组,首先可以在无性生殖的真菌中进行基因在染色体上的定性研究,其次可以在半知菌中进行杂交育种。在半知菌的演化中,有些似已固定用准性生殖来代替有性生殖了。
核循环 像其他生物一样,真菌通常也有一个相当于绿色有机体配子体和孢子体的单倍体和二倍体结构的循环(图2)。二倍体阶段由核配开始,到减数分裂完成。但是,在大多数真菌中,菌体并没有明显的单倍体和二倍体的交替,代表二倍体阶段的仅仅是合子〔见世代交替(植物)〕。
生 态
天然真菌或人工培养真菌与生存条件之间的相互关系。真菌种类很多,对生存条件的要求和反应各不相同。它们有的生活在水域,有的生活在陆地;有的生长在热带,有的生长在寒带;有的全年都能生长,有的只在某个季节才生长出来;有的寄生于其他生物,有的与其他生物共生,也有的只能生长在其他生物的腐败残骸上。这些不同的生活方式和生长习性,都是在漫长的发育历史中形成的。每种真菌对养料、温度、湿度、酸碱度、氧、光线等都有特殊的要求。
腐生真菌及其习性 腐生真菌是数量多、分布广、作用大的一群真菌,能把禁锢在有机体内的化学元素归还给大自然,使碳素循环顺利进行。当它们的菌丝与基物紧密接触时,在外酶的作用下,可将周围不溶性的有机物水解为可溶性的有机物,将大分子的有机物分解为小分子有机物,并通过菌丝壁的直接扩散作用进行吸收。首先吸收的是可溶性的糖和氨基酸。大多数真菌能够迅速分解简单的碳水化合物,如糖类、淀粉、半纤维素和某些蛋白质。对纤维素、脂肪和油类的分解比较慢,而且常常需要特殊的真菌类型。木质素、蜡和单宁最抗分解,只有少数真菌才能分解它们。
土壤真菌 土壤中有丰富的腐生真菌基质。真菌主要分布于距地表24厘米以内的土壤中,在0~10厘米间最多。它们的数量和种类随土层的加深而递减(见土壤微生物区系)。
水生真菌 水体中生活并能完成生活史的真菌(见水生真菌)。
嗜热真菌 40℃以上能生长繁殖的真菌。常见的有毛霉、毛壳菌、篮状菌、嗜热子囊菌等属的一些种。农业上利用它们制造堆肥和栽培蘑菇时制备培养基。
喜冷真菌 10℃以下能生长的真菌。它们生长在极地、高山、雪下和冷藏库中。南极的月平均温度一般在0℃以下,但真菌比较普遍,已发现有结实串孢霉、总丝霉等多属半知菌和酵母。在海拔4000米的高山和雪线附近也有蘑菇子实体生长。 冷藏库的温度一般都在0℃以下,但在贮藏的肉类和果蔬上常有枝孢、枝霉、青霉和毛霉等真菌为害。
大气真菌 空气不像土壤和水体那样能作为真菌的生活环境,但源出陆地的某些真菌菌丝的碎片和孢子却构成了空中孢子区系。大气真菌主要指对流层及其以上10公里处的真菌,只有少数的同温层中。空中真菌的类型和密度,因季节、时间、气候和人类的活动而发生变化。一般来说,在干旱气候的白昼,枝孢和链格孢等干孢子类真菌占优势;在潮湿的夜间,子囊孢子和担孢子比较多。
寄生真菌及其寄主 植物寄生真菌 几乎所有的植物都受真菌的侵袭和危害。受害最大的是栽培植物,往往造成减产和品质降低。
动物和人的寄生真菌 真菌能侵染各种动物和人,引起病害。使高等动物受害的绝大多数为半知菌,只有少数是接合菌和子囊菌。这些病原真菌都是兼性寄生菌,大多可以从土壤中分离出来。从人体分离出来的真菌约120种,其中半数是常见的腐生菌。至今还没有发现专性寄生于人的真菌。
真菌寄生真菌 从另一种真菌获取养料的真菌。已知有30多种壶菌寄生于其他壶菌和卵菌上。有些毛霉目的真菌常寄生于毛霉目的其他成员上,并具有高度的专化性。有些半知菌是卵菌、接合菌、子囊菌、担子菌和其他半知菌的寄生物。这类真菌可用于害菌的生物防治。
共生真菌及其伙伴 同其他生物生活在一起,彼此受益的真菌(见地衣、菌根、虫道真菌)。
起源和演化
真菌的起源和演化众说纷纭,但直到现在还没有一个能被真菌学家们普遍接受的学说。一个学派认为真菌是由藻类演化而来。这些藻类因丧失色素而从自养变成异养,生理的变化引起了形态的改变。他们根据性器官的形态和交配方式进行推测,认为鞭毛菌来自绿藻,接合菌来自接合藻,子囊菌来自红藻,担子菌来自子囊菌。另一个学派认为除卵菌来自藻类外,其余的真菌来自原始鞭毛生物。鞭毛生物水生,具有一至数根鞭毛,有的含叶绿素或红色素,有的含其他色素或无色素。含叶绿素的演化为藻,无色素的演化为菌。20世纪中叶以前,有关真菌系统发育的设想、方案和系统树,除生态习性和营养方式外,主要是以形态学为基础。现在人们普遍认识到多数活有机体的基础生物化学是相同的,而其差异如细胞壁的组分、DNA和RNA的含量、DNA中的G+C克分子百分含量、氨基酸的排列顺序、血清反应以及某些代谢产物的含量与合成途径等,则有利于确定它们之间的亲缘关系。例如,根据血清反应,推测藻状菌是管藻目中靠近无隔藻的两个早期分支,后来又衍生出许多支系,直至今天的炭角菌、羊肚菌、锈菌、鸟巢菌、牛肝菌,鹅膏菌等类群。再如,根据赖氨酸的生物合成途径,推测壶菌可能是从与眼虫藻有关的鞭毛生物演化而来。壶菌丧失鞭毛以后,由于菌丝发达而演化成接合菌,从接合菌再演化为子囊菌和担子菌。所以,真菌的演化途径可能是:与眼虫藻有关的鞭毛生物→壶菌→接合菌→子囊菌→担子菌。这是真菌演化的主轴,相当于绿色植物中的:绿藻→轮藻→苔藓→维管束植物。过去认为卵菌是在产生高等真菌的演化线上的一个早期分支,现在却认为它属于另一个演化线,大概起源于与金藻有关的无色素类型,因而被排除在真菌以外。
分 类
真菌分类系统很多,现仅根据英国真菌学家G.C.安斯沃思等人(1971,1973)制订的系统,将真菌界分为2门、5亚门、20纲(见真菌界)。
意 义
真菌是一项丰富的自然资源。人和动物每年消耗大量的真菌菌体和子实体(见食用菌);真菌也是重要的药材(见药用真菌)。
真菌的某些代谢产物在工业上具有广泛用途,如乙醇、柠檬酸、甘油、酶制剂、甾醇、脂肪、塑料、促生素、维生素等。而且这些东西都能进行大规模的生产(见有机酸发酵微生物、酶制剂微生物、维生素发酵微生物、甾体化合物的微生物转化作用)。
在农业、林业和畜牧业中,真菌既有有害的一面,又有有利的一面。真菌能引起植物多种病害,从而造成巨大的经济损失。例如,1845年欧洲由于马铃薯晚疫病的流行摧毁了5/6的马铃薯,中国由于1950年的小麦锈病和1974年的稻瘟病而使小麦和水稻各减产 120亿斤。但是,菌根能对多种木本和草本植物提供必不可少的营养。在真菌的腐解作用中,它使许多重要化学元素得以再循环。真菌直接或间接地影响着地球生物圈的物质循环和能量转换。
参考书目
C.J.亚历克索普洛斯、C.W.米姆斯著,余永年、宋大康等译:《真菌学概论》,农业出版社,北京,1983。(C.J.Alexopoulos,C.W.Mims,John Wiley & Sons,New York,1979.)
J.H.Burnett,Fundamentals of Mycology, 2nd ed.,Edward Arnold Ltd., New York,1976.