着床(卷名:生物学)
embedding,implantation,nidation
━━━━━━━━━━━━━━━━━━━━━━━━
着床的准备
着床的类型
中央或表面着床
偏心着床
完全侵入着床
无延缓着床和延缓着床
着床的步骤
定位
粘着
穿入
着床机制的研究
同步化和蜕膜反应
子宫分泌物的作用
卵巢激素的影响
胚泡的作用
其他因素
结语━━━━━━━━━━━━━━━━━━━━━━━━
亦名“植入”,指胎生哺乳类动物的早期胚胎和母体子宫壁结合,从而建立母子间结构上的联系以实现物质交换的过程。着床后的胚胎摄取母体血液营养继续发育。着床是母子双方有准备,相互配合的结合过程;是基因型不同和发育阶段不同的两代个体间的一种既紧密靠近,又保持独立的结合。
着床是胎生哺乳动物生殖过程中一个重要环节。对着床的了解,既可为哺乳动物胚胎移植开路,也可为抗着床措施提供线索;从理论上推测,抗着床措施应该是比较理想的避孕方法。
着床的准备
真兽类动物的受精卵在输卵管中缓慢运行(3~5天),经过连续多次分裂之后形成桑椹胚进入子宫腔,继续发育成为囊胚(有些种类是在囊胚期进入子宫腔)。真兽类囊胚特名为胚泡(图1)。
 胚泡外围绕着透明带,依靠吸取子宫腺分泌的子宫乳继续发育。随着胚泡的长大,透明带胀大变薄被撑破而脱落(或被溶解);胚胎从包膜中脱出。胚胎外壁的滋胚层细胞裸露,直接与子宫内膜接触。由于子宫“着床点”的吸引,或者再加上胚泡主动的选择,胚泡在一定位置附着于内膜,称为定位。
胚泡外围绕着透明带,依靠吸取子宫腺分泌的子宫乳继续发育。随着胚泡的长大,透明带胀大变薄被撑破而脱落(或被溶解);胚胎从包膜中脱出。胚胎外壁的滋胚层细胞裸露,直接与子宫内膜接触。由于子宫“着床点”的吸引,或者再加上胚泡主动的选择,胚泡在一定位置附着于内膜,称为定位。着床的类型
按照着床时胚泡在宫腔内的位置、滋胚层细胞和内膜上皮细胞接触及结合的程度,常把着床分为:
中央或表面着床 着床时胚泡体积扩大,充满子宫腔一段空间,滋胚层大面积地与内膜上皮接触。属于此型的动物很多,如有蹄类、食肉类、兔形类、一些灵长类(跗猴、狐猴、宽鼻猴等)、贫齿类和翼手类等。兔在着床初期属于表面型,但以后内膜上皮分解,胚泡又埋入内部。
偏心着床 胚泡体积较小,子宫腔相当大,胚泡深嵌在宫腔一侧的上皮陷窝之中。滋胚层细胞的许多微绒毛突起插进内膜上皮细胞的间隙。以后上皮消融,胚泡被包围在基质之中。此型包括许多啮齿类如大鼠、小鼠等。
完全侵入着床 有些动物的胚泡附着到子宫内膜之后,侵蚀内膜上皮细胞,然后胚泡穿过上皮基膜,完全离开宫腔,钻入内膜基质之中所以胚胎是在子宫壁内生长发育的。属于此型的有某些蝙蝠、食虫类的刺猬以及灵长类的狭鼻猴、猩猩和人等。
无延缓着床和延缓着床 动物的胚泡进入宫腔之后,不一定就着床而继续发育。有些种类的胚泡需经历一段休眠期后才进行着床,因此按有无滞留分为无延缓着床与延缓着床。各种动物的胚泡从到达子宫到开始着床的时间不同,虽然如此,附表中所列的大都是所谓无延缓着床,即胚泡进入宫腔后不久即开始着床而继续发育。
此外,还有延缓着床的情况,可分为兼性和专性延缓着床两类型。属前一类型的动物一般不出现延缓着床现象。但在母兽处于泌乳、应急、饥饿等情况时,进入宫腔的胚泡可处于停滞状态,一直要等到仔兽断乳或条件好转,胚泡才又恢复着床。大鼠、小鼠、沙鼠等属此类型。
属于专性延缓着床的,胚泡进入宫腔后有相当长的一段时间是游离的,并转入休眠状态。经过几个星期甚至几个月,才恢复活动而出现着床行为,胚胎继续发育下去。延缓着床期间卵巢黄体合成甾体激素的活力处于很低水平,直到临近着床才又升高。一般认为延缓着床是受环境因子,如光照、温度、食物等影响。属于此类型的动物有麋鹿、獾、熊、貂、黄鼬、海豹、袋鼠等。
着床的步骤
以胚胎在子宫壁内发育的侵入型着床(鼠、兔)为例,胚泡经转运,定向,透明带溶解之后开始着床,着床步骤有定位、粘着、穿入等(图2)。
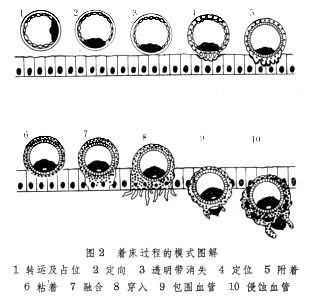
定位 胚泡进入子宫腔后,在一定位置靠近内膜。由于胚泡本身所发生的机械压力和子宫平滑肌的蠕动作用的相互配合,能将胚泡安置在子宫角腔中一个适当的位置。如果将塑料或葡聚糖做成的小珠代替胚泡灌入假孕鼠子宫腔,也会出现类似正常妊娠的均匀分布情况。着床位点的选择可能决定于胚泡和内膜的某种特性。
粘着 胚泡在内膜上附着之后进一步贴紧粘着,除非损伤胚泡,就无法使它与内膜分开。粘着的直接原因是由于滋胚层细胞表面的糖蛋白受到酶解作用产生粘性。在粘着之后,母、子细胞接触面出现微绒毛交错,或者由内膜上皮细胞的膜上长出突起去连接胚泡的滋胚层细胞。此后滋胚层细胞和内膜上皮细胞的接触面上形成桥粒和连接复合体等专门的固着结构。

穿入 已粘着的胚泡其滋胚层细胞分泌某种物质(可能是蛋白酶) 使相邻的内膜上皮的细胞之间出现缝隙。胚泡与内膜溶合后挤进缝隙而钻入内膜的基质之中。有人认为胚泡进入内膜向内迁移,是受到微血管散发物质的吸引,直至最后停靠在微血管的边缘。例如在家兔,滋胚层细胞似乎对准一处微血管作为前进的目标,穿过上皮组织,最后破坏微血管壁的内皮细胞;在狗、猫等动物的滋胚层细胞则对准子宫腺的口部,沿着腺管穿入。
胚泡的滋胚层细胞所接触的内膜上皮逐渐改组。改组有 3种方式:①上皮细胞移位。比较常见,在胚泡穿入前,上皮细胞从基底膜上松散开来,并出现大面积的移动,让滋胚层细胞经过基底膜面钻入基质。在此过程中常有个别上皮细胞脱落下来,破碎的残片通常被滋胚层细胞所吞噬。②细胞融合。滋胚层细胞与上皮细胞接触后,发生融合,两种细胞的胞质合并,上皮细胞失掉它原有的特性。如家兔胚泡的着床。③侵入。两类细胞皆保持完整无伤,只是滋胚层细胞挤进上皮组织。最初有滋胚层细胞的突起插入上皮细胞间的缝隙,继而插入部分扩展,一直深入上皮组织的基底膜,贯穿该膜而进入基质内部,以后形成血窦绒毛型、结缔绒毛型或内皮绒毛型胎盘动物的着床,胚泡都深埋在内膜基质中。
着床机制的研究
同步化和蜕膜反应 着床时胚泡和子宫都处于生长发育的动态过程中,只有双方同时达到适宜的成熟程度(同步化),着床才能成功。所谓适宜,在胚胎方面指从桑椹胚成长为胚泡,并从透明带膜中孵出;在子宫方面指其内膜从不敏感状态转入敏感状态能够接收胚泡着床。任何一方配合不当就会导致着床失败。
在卵子受精之后,子宫内膜在有关激素调节下发生明显的变化。在垂体分泌的黄体生成素和促黄体素(LH与LTH),使卵巢中的排卵卵泡发育成为妊娠黄体,并促使黄体分泌孕酮。孕酮激发子宫腺进入分泌状态。在黄体时相卵巢又分泌少量雌激素。在孕酮和雌激素的协同作用下,子宫内膜增生,同时转入敏感状态。这时的内膜,若其表面受到胚泡着床的刺激或者在实验条件下受到机械性创伤或接触到油滴、气泡等,子宫内膜在着床点区域便出现水肿或增生肥大、细胞分裂相增多、基质内微血管舒张、血管的通透性升高、细胞的代谢水平升高,酶活性加强等。更为重要的变化是基质细胞的蜕膜化。基质细胞本是小型梭状的成纤维细胞,转变后成为具有上皮细胞性质的大型细胞──蜕膜细胞;后者在相连的细胞膜上出现接合点。这一变化首先发生在胚泡着床的部位并包围胚泡,不久扩散到全部子宫内膜。这种在妊娠期高度发育与特化了的子宫内膜称蜕膜,形成蜕膜的一系列变化称蜕膜反应。蜕膜细胞常有多核现象,细胞质中集聚许多糖原颗粒和脂肪滴,还含有许多溶酶体。
人类在月经周期的黄体时相内,子宫内膜出现类似蜕膜反应的转化,称为“前蜕膜”。若卵子不受精,前蜕膜则在行经时脱落而随血流排出;如果受精,则在孕卵着床时(前蜕膜)转化为蜕膜。蜕膜细胞能为发育的胚胎提供营养物质,并限制滋胚层细胞的侵蚀,因而起到保护母体的作用。人类蜕膜细胞还分泌促乳激素,它有促进胚胎生长的作用,同时也是维持妊娠的重要激素。在无蜕膜的动物如猪、羊、牛等偶蹄类,子宫内膜在胚泡着床时不出现蜕膜反应,在分娩时,子宫内膜也不剥离脱落(见胎盘)。
子宫分泌物的作用 根据假孕期的大鼠子宫灌注组胺能引起蜕膜反应,而灌注组胺的拮抗剂则降低胚泡的着床数和蜕膜反应的比率。有人认为子宫壁中的肥大细胞受到雌激素高峰的激发释放组胺,从而引起蜕膜反应。但肥大细胞产生组胺没有专一性,其所引起的反应遍及整条子宫角。现已发现胚泡还能产生雌激素,局部刺激子宫壁的肥大细胞释放组胺,组胺能引起蜕膜反应,这对着床区蜕膜反应的发生提供了一个合理的解释。
子宫分泌液中所含的蛋白酶对于消化着床前胚泡外围的透明带起主要作用。其中一种酶还能改变胚泡表面糖蛋白的结构,产生粘性,使胚泡粘附于内膜。该酶的分泌是由雌激素调节的。妊娠子宫还有其他几种分泌物,如子宫珠蛋白、子宫糖蛋白等对于即将着床的胚泡发生何种作用还知道不多。
卵巢激素的影响 卵巢激素对着床的调节,各类动物并不完全一致。在妊娠初期切除卵巢,多数动物不能实现着床,需要及时注射孕酮;或者注射孕酮外还要增加雌激素。可知此种型式的着床要求孕激素与雌激素相对的平衡。可是在豚鼠切除卵巢后不需补充激素,胚泡仍能着床。
正常妊娠的大鼠子宫,在第 3天内膜上皮发生大量的细胞分裂,在第四天出现一个雌激素高峰,频繁的细胞分裂使增殖细胞由上皮转移到基质之中,即在此时刻发生着床现象。雌激素促进分裂细胞 DNA的合成以及一些酶的合成;后者消除子宫内膜对胚泡的抵制,并促使胚泡的透明带溶解与脱落,所以雌激素被视为着床的起始因子。如果在这时期给予过量的雌激素,却又会破坏着床。这种情况在鼠类的实验和人类的避孕实践中已为大量数据所肯定。
胚泡的作用 胚泡对子宫内膜乃至整个母体发出许多信号,以协调着床的步伐,其作用可以从两方面讲。
机械刺激 如把玻璃珠、蜡珠或琼脂糖珠灌注到假孕大鼠的子宫腔中,可以引发一系列着床的变化。可见胚泡对子宫内膜的局部压力是促使内膜发生蜕膜反应的一个因素。
在某些特殊情况下,胚泡也能在子宫以外非内膜位置,例如在输卵管中,输卵管的浆膜、系膜等部位着床发育,称为“宫外孕”。其所引起的周围结构的变化大致和着床相似。胚泡能在宫外着床发育,说明子宫内膜并非着床的唯一必要条件;而胚泡才是决定着床的主要方面。
胚泡分泌物和代谢物质的作用 胚泡分泌的激素和排出的代谢物,分量虽然极少,但在局部可发生很大的作用。例如代谢旺盛的胚泡释出较多的二氧化碳必定会转移到子宫内膜的细胞中,然后进入微血管,靠母体血液循环将它排除。这样就在胚泡附近形成一个二氧化碳的梯度场,由于碳酸酐酶的干预,涉及碳酸盐-碳酸氢盐之间的平衡,可能影响到这些细胞内环境中酸度值的变化,从而促使子宫内膜基质细胞发生蜕膜反应。二氧化碳还可使滋胚层细胞表面与内膜上皮细胞表面的糖蛋白的粘性升高,从而有助于胚泡的粘着。
在离体条件下,家兔和猪的胚泡着床前仍能利用标记的前体合成几种甾体,其中包括雌激素和孕激素。在活体子宫中,家兔、猪和牛的胚泡也能合成甾体。
与着床有关的甾体激素的作用是局部的,而不是全身性的。例如子宫内膜局部微血管的通透性的升高,范围很小,这是胚泡分泌雌激素的结果。雌激素能降低前列腺素 PGF2α的合成,由于 PGF2α有溶黄体作用,因此雌激素就有促黄体的效果。黄体能继续分泌孕酮而促进着床。
人类胚泡的滋胚层在很早阶段,甚至在着床前(排卵后第 6、7天)就分泌绒毛膜促性腺激素(hCG);它在胚泡液中的浓度比释放在母体血液中的要高出一倍。它能促使排卵后的黄体转变成为妊娠黄体,分泌孕酮等妊娠所需要的激素。
其他因素 母体的卵巢和垂体是着床的一个重要条件。摘除卵巢,依靠注射孕激素或再加上雌激素的补偿,胚泡仍能着床。在受精大鼠的妊娠早期,摘除垂体,则胚泡不能着床;但若补充促乳素,则可使胚泡处于延缓着床状态;如能再给予雌激素,就能着床。下丘脑是比垂体更高级的神经内分泌调节的结构,下丘脑分泌黄体生成素释放激素(LH-RH),促使垂体前叶分泌LH和FSH。但已发现大剂量使用LH-RH时有抗生育作用,其中包括抗着床效果。大剂量LH-RH促使雌二醇水平升高并抑制羟甾脱氢酶活性,直接导致子宫失去敏感性,因而胚泡脱落。
外界环境也可影响着床。例如新受孕的某一品系的雌鼠若是与异品系的雄鼠或其排泄物接触,则妊娠中止,即所谓布鲁斯效应。现知这是由于异品系气味经过嗅觉通路传入中枢,导致下丘脑增强促乳素抑制激素的释放,因而垂体前叶促乳素分泌降低,引起黄体分泌孕酮不足,妨碍胚泡着床。正因如此,若能给予接触异种气味的初期孕鼠以促乳素,则可以挽回着床而生育。
结 语
着床是胎生胚胎发育早期阶段的一个重要环节。在着床之前,胚泡在宫腔内游离,它是新个体,有遭排斥脱落危险。经过附着、穿入内膜(属于血窦绒毛型胎盘者)之后,则母子两方面紧密结合,因而更有利于胚胎的生长发育。对于着床的理解,近60年来人们从比较胚胎学、生殖内分泌学、实验形态学以及生物化学和分子生物学等各方面积累了相当丰富的知识。但是着床的机制究竟如何仍待探索。在受精卵发育成为胚泡的同时,子宫及内膜也同步化地经历一系列的变化,逐步成为可以接纳胚泡继续发育的基地。脑垂体、卵巢、子宫与胚泡之间相互影响,关系相当复杂,有促进、调节、制约,存在一定时空顺序。值得注意的是胚泡有主动的一面。由胚泡发出的微量信息,在局部范围可引起子宫内膜的变化;甚至在宫外位置也能转变附近组织原来的属性。还有一点是子宫的发育必需先经一系列雌激素和孕激素协调的刺激后,才能接受机械刺激。许多有关着床的知识是从鼠类和兔类的实验研究推测出来的。上述仅是一般化模式,主要说明了血窦绒毛型情况;至于上皮绒毛型等其他着床类型则未必有这样复杂。近20年来,由于人类计划生育研究的促进,试管婴儿的开展,家畜胚胎移植及转基因动物等研究的兴起,一方面要利用已有的着床知识,另一方面也为着床的研究创造了有利的条件。
参考书目
曲漱蕙等:《动物胚胎学》,人民教育出版社,北京,1980。
K.Yoshinaza et al., Implantation of the Ovum,Havard University Press, Cambridge, 1976.