棘皮动物门(化石)(卷名:生物学)
Echinodermata(fossil)
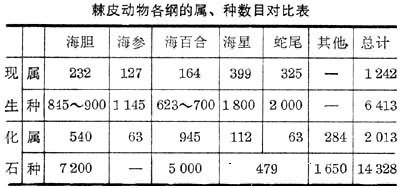
棘皮动物是海生无脊椎动物,以其复杂的水管系统、发达的骨板(除海参外)、五辐射对称的身体和表皮具各类棘刺而区别于其他动物门。包括大量已绝灭的形状各异的种类。棘皮动物化石种类极多,有些种类的数量也很多。目前已知现生棘皮动物有5纲1242属6413种,化石棘皮动物 21纲2013属14328种(见表)。
分类
棘皮动物曾以其生活方式被分为有柄或固着亚门和无柄或游走亚门。前者包括海百合纲和已绝灭的海林檎纲、海蕾纲、海座星纲;后者包括海胆纲、海参纲和海星亚纲。随着对棘皮动物研究的日益深入,发现了不少新的化石种类。同时认识到生活方式不能成为分类的依据。
近来根据棘皮动物水管系统生长的特点将棘皮动物分为海胆、海扁果、海百合、海星4亚门21纲9亚纲。
海百合亚门 体呈五辐射对称,多数具萼、茎和腕,由一系列骨板围成的萼部多呈球形、锥形或柱形,与腕共同构成冠部,其口孔向上。通常营固着或浮游生活。共约1200属,生活于早寒武世(距今约6亿年)-现代,除海百合纲外,均在古生代(距今6~2.8亿年)先后绝灭。
始海百合纲 具萼、茎和腕,萼部由 20~500块骨板相间排列,口孔位于口板上,具肛孔、水孔及生殖孔。早寒武世-早志留世,32属,产于欧洲、北美、北非、西伯利亚。
海林檎纲 具萼及腕,茎有或无,萼部由13~200块骨板不对称排列,萼板上具板孔,形成复杂的沟孔系列,沟孔散布在整个萼上的为双孔目 (Diploporita),集合成一个特殊孔菱区的为孔菱目(Rhombifera)。口孔位于腹面中央,肛孔位于口板上方覆有肛板,水孔覆有筛板。早奥陶世-晚泥盆世,102属,产于北美、欧洲、亚洲。因其形状似林檎果(北方称沙果,南方称花红)而得名。
拟海百合纲 萼部椭球形由30~175块骨板组成,排列不规则。两侧对称。具茎及腕2~4个,腕可分叉。中泥盆世,15属,产于北美、苏联爱沙尼亚。
垫海蕾纲 萼呈倒梨形,有茎无腕,萼部由20块骨板规则排列为底板、辐板、三角板和口板四圈。间辐位置上有形状不定的下三角板,步带内有呈双列状规则排列的等大盖板。中奥陶世,1属,产于北美。
拟海蕾纲 萼呈梨形、有茎,有腕羽,萼板多;在反口面除底板、辐板各5块外,尚有5对双腕板和许多间腕板,在口面有花瓣状步带板及间辐板各 5块。步带为盖板所覆盖。早、中奥陶世,3属,产于北美及苏联爱沙尼亚。
海蕾纲 萼梨形或钟形,多数有茎,具腕羽,体腔不扩展入腕羽内部。底板3块,辐板5块,辐板呈叉状,叉口间嵌入步带。间辐板小,三角形。志留纪一二叠纪,95属,世界性分布。
鳞海林檎纲 萼部倒锥形,无茎、具双列式腕,具大量呈叠瓦状排列的小骨板,口面骨板具缝孔,肛锥位于口面边缘。早寒武世,1属,发现于北美。
海百合纲 萼部花冠状,有茎、具腕,萼部骨板呈环状规则排列,具内底板者为双环海百合,仅有底板者为单环海百合,其上为辐板,辐板间有间辐板。腕可分叉或呈羽枝状,腕内为体腔的扩展部分。海百合类的茎由一系钙质茎环连接而成,茎环断面呈圆、椭圆或多边形,其中央有圆形、五角形、方形等茎孔。其下分为游离海百合 (Inadunata)、可曲海百合(Flexibilia)、圆顶海百合(Cameratal)、关节海百合(Articulata)4个亚纲,前 3个限于古生代,后者出现于三叠纪并延续至今,其中一些进化为无茎种类。中寒武世-现代,1009属,世界性分布。
海扁果亚门 萼部多背腹扁平,形状不规则,具长短不等的茎尾,多具腕或腕突。不呈五辐射对称,有些可呈两侧对称状。可能平卧海底,并可向茎尾方向移动(图1)。曾被称为海果核类 (Carpoid)。是棘皮动物中较特化的一类,在古生代已全部绝灭。
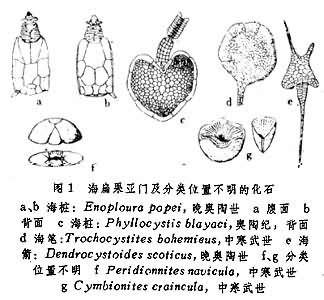
海桩纲 萼扁平,由数列大小不等的骨片组成,腹面略凹而背面略凸,不对称。茎尾粗短。具腕突,口孔位于腕基部萼的内面,肛门位于口的对端。中寒武世至中泥盆世,32属,分布于西欧、北美、南美、澳大利亚和非洲南部(图1a、b、c)。
海笔纲 萼为不规则勺状,由不规则骨板构成,内层骨板小而数目多,外层骨板大而数目少。具茎尾。无腕,口与肛门彼此靠近位于萼的同一极。中寒武世,4属,分布于西欧及北非(图1d)。
海箭纲 萼呈不对称的箭簇形,近两侧对称,由大量不规则骨板组成。具单个的细腕。茎尾长。口孔位于腕基与肛门之间,肛门位于萼基部的一侧。中寒武世至早泥盆世,13属,产于欧洲、北美、澳大利亚(图1e)。
一些古生物学家认为海桩纲显示了明显的脊索动物特征而建立了一个新的亚门──钙索动物亚门(Calci-chordata)。其下分为角首目(Cornuta)与帽首目 (Mi-trata), 前者见于西欧、北非和美国的中寒武世到晚奥陶世地层;后者见于西欧、北美、南美、澳大利亚和非洲南部的早奥陶世至中泥盆世地层。其身体的两部分被称为头和尾或体部和尾部。而脊索动物另 3个亚门的祖先均与帽首目有密切的关系。有人指出头索亚门可能与捷克波希米亚地区中奥陶统的Lagynocystis有关,尾索亚门可能与产于上述地层的Balanocystites有关,而脊椎动物亚门则与产于法国和波希米亚中、晚奥陶世和德国早泥盆世的 Mitrocystites有关。它可能是脊椎动物祖先类型中的一个旁支,因真正的脊椎动物可能在晚寒武世即已出现。
海星亚门 海星类体呈明显的五辐射对称的星形,平扁,可以腕在海底行走。整个亚门只有唯一的星形纲(Stelleroidea),但据其身体结构上的特征分为 3个亚纲。最早出现于早奥陶世,至今仍有现生代表。
体海星亚纲 体扁,腕宽而短,中央盘呈钝五角形,与腕区别不明显。腕具双列步带板,步带枝平行状自步带伸向前外方,直达边缘板。口孔在腹面,呈大五角形,其边缘具口角板。早奥陶世-现代,8属,分布于欧洲、美洲及澳大利亚。
蛇尾亚纲 体五角星形,圆形中央盘与细长的腕间区别明显,动物的脏器不伸入腕中,腕可多次分枝,由多列骨板互相关节,活动自如。口孔位于腹面,无肛门,筛板位于腹面。早奥陶世-现代,338属,世界性分布。
海星亚纲 体扁,五角星形,中央盘与腕区别不明显,脏器伸入腕内,腕较宽。有些种类亦可分枝。腕由多排骨板紧密排列而成。口五角形,位于腹面中央,肛孔位于背面中央,筛板位于肛孔附近。早奥陶世-现代,430属,世界性分布。
海胆亚门 壳多呈球形或半球形,无茎无腕,可自由活动(图2),其水管经向生长。最早出现于早寒武世,除海胆纲和海参纲外,均已在古生代绝灭。
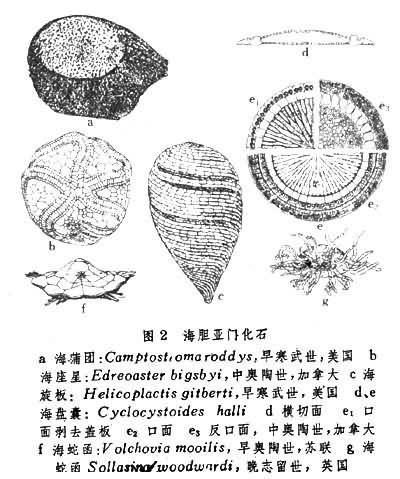
海旋板纲 壳纺锤形或陀螺形,由长方形或菱形小骨板呈螺旋状旋绕排列。骨板互不缝合,外被坚硬皮膜,壳可能具胀缩和弯曲的能力。步带,亦呈旋绕形,口及肛孔位于壳之两端。早寒武世,3属,产于北美(图2c)。
海座星纲 壳球形,腹面具柔软皮膜盘司固着。骨板排列规则,背腹面间具边缘板。步带自口部伸出,向外延伸并向左右扭曲,口及肛孔均位于壳的背面。早寒武世至中石岩世,35属,分布于北美及欧洲(图2b)。
海蛇函纲 壳盔形,腹面平、背面呈穹状。口位于腹面中央,有五步带由口部向外辐射,每一步带具 8对大而特殊的步足。肛孔位于背部侧方,背方具多角状骨板,分为5个间辐区。早奥陶世-早石炭世,6属,分布于北美和欧洲(图2f、g)。
海蒲团纲 壳饼形,具原始的方解石骨板,体四周具放射肋。在未发现其骨板前曾被置于腔肠动物门。早寒武世,1属,产于北美(图2a)。
海盘囊纲 壳盘形,口面向上,反口面向下。口面具中央盘,有五条步带向四周伸出,整个被骨板覆盖。口孔位于口面中央,肛孔位于口边近边缘处。中奥陶世-中泥盆世,8属,产于北美及欧洲(图2d、c)。
海参纲 体筒形,不具硬壳。小骨板散于体壁内,体内有锚状骨针及窗孔状小骨板。可能自中寒武世即已出现,可靠的化石发现于中奥陶世,现代仍有不少,200属,世界性分布。但所发现的化石除极少数外,均以零散骨针、骨板为代表,描述了一些形态属及形态种。
海胆纲 壳球形、半球形或心形等,由极多骨板呈叠瓦状排列(软壳)或相互联结(硬壳)构成胆壳,壳外具各种海胆刺及叉棘。壳上可分出明显的步带及间步带区。口面位于腹面中央或偏前方;肛孔位于反口面中央或偏后方的间步带区,偶有移至口面者。目前分为两个亚纲,围海胆亚纲(Perischoechinoidea)壳多柔软,不具外鳃,步带板2~20列,间步带板1~14列。其中沟头帕海胆目(Bothriocidaroidea)、棘囊海胆目(Echinocystitoidea)和古海胆目(Palaechinoida)有约 40属均已在古生代绝灭,而头帕海胆目(Cidaroida)除 10余属发现于古生代外,其余均繁盛于中生代至现代。真海胆亚纲(Eu-echinoidea)具外鳃,步带及间步带各具2列骨板。约700余属,分为4超目,约20目 80余科。真海胆类可能起源于头帕海胆目。海胆纲共约772属,最早的化石记录见于晚奥陶世,世界性分布。
系统发生及进化
棘皮动物有两侧对称的担轮幼虫期,显示其成年个体的辐射对称可能是次生的。一般认为棘皮动物起源于具两侧对称的祖先。有人认为可能是一种具3对体腔囊的身体柔软的两侧对称型祖先,更多人认为可能是一种具3对体腔囊和具5条与体腔相通的生在口部四周的触手的五触手型祖先。它们首先改营固着生活,而导致身体结构上发生辐射对称,不过后来有些又转而营自由生活,但辐射对称特征却保留下来。一些化石棘皮动物类别也不具辐射对称也足以证明。棘皮动物的卵裂方式和具三胚层、真体腔、后口等特征都与脊索动物极为接近。但棘皮动物没有头和高级神经中枢以及具担轮幼虫阶段,因而有人认为它们与星虫门(Sipunculida)、帚虫门(Phoronida)、腕足动物(Brachiopoda)等有密切关系。
近年以棘皮动物中的海桩类建立钙索动物门的意见又再次被提出,暂不论多数古生物学者,还未接受这一假说,但也使得人们注意到脊索动物终究还是与棘皮动物极为接近的。
许多棘皮动物化石发现于早寒武世,而且构造复杂,按照传统的渐进论观点,认为在此以前棘皮动物已经经历了极漫长的进化过程。按照当前的间断平衡理论则它们的出现可能是在一定的环境条件中形成的,而它们的很快绝灭或种类很少、分布局限正说明了这一问题。
中国的棘皮动物化石
相对说来,中国的棘皮动物化石数量、种类均较少,发现最多而普遍的是海百合茎。在云南、贵州、陕西等省的奥陶纪地层中产有中国海林檎(Sinocystis),在黑龙江中泥盆统及贵州中石炭统产海蕾化石。在贵州、河北、四川等地发现海百合化石。海胆化石在中国发现较多,在湖南、广西、江西等地的早石炭世地层中发现瓜海胆(Melenechinus)、古海胆(Palechinus)、拉文古胆(Lovenechinus)和头帕海胆类化石;在西藏、新疆、台湾、南海等地发现中生代及第三纪海胆化石。自60年代初期以来,蛇尾及海星化石陆续发现于贵州、广东、陕西、新疆等地。最近在广西桂林附近还找到了保存极好的海百合化石。海参骨针化石亦偶有报道。
此外还有一些具辐射对称或两侧对称特征,又具有骨针或骨片的化石被归入棘皮动物门,如澳大利亚早中寒武世的海豆类(Cyamoidea)及海穹类(Cycloidea)被建立一个新的亚门──海独亚门。有人将无茎无腕,口孔为筛板所覆盖的海扁果类建立为栉海林檎纲 ( Stenocys-toidea)。对于这些类别的分类位置及归属各家意见不一。