生态系统(卷名:生物学)
ecosystem
一定空间中的生物群落与其环境组成的系统,其中各成员借助能流和物质循环形成一个有组织的功能复合体。“系统”一词有两层基本含义:一是它由一些相互依赖、相互作用的部分所组成;二是这些部分按照一定的规律组织在一起,从而使这个整体具备了统一的功能特征。把生物与环境视为一个统一整体的概念非常古老,但根据生物学研究而明确提出这种观念则只有 100多年历史。德国学者K.A.默比乌斯1877年提出“生物群落”概念时,最初便有环境的含意。美国学者S.A.福布斯1887年提出“微宇宙”。“生态系统”一词是1935年由英国生态学家A.G.坦斯利提出的,与此概念相近的还有苏联学者B.H.苏卡乔夫1940年提出的“生物地理群落”。
生态系统概念一经提出,很快为生态学家普遍接受。但由于生态系统的研究规模大并且要求综合研究手段,这在当时的物质技术条件下是很难满足的,所以在最初的20多年间研究的范围比较小。直到60年代,人口、资源和污染等问题受到广泛重视之后,生态系统概念才开始在人类实践活动中发挥出越来越大的作用。当今,无论小面积的害虫防治、山林管理,还是大范围的农业规划、水土保持,甚至全国性的国土整治等问题,都需要从生态系统的观点来考虑。生态系统理论已成为现代环境科学的基石。
结构 一般由3方面描述,即营养结构、空间结构和时间结构。
营养结构 生产者主要是绿色植物,能利用简单的无机物质制造复杂的有机物;消费者主要是动物,以植物或其他动物为食,又分为一级消费者(植食动物)、二级消费者(肉食动物)、三级消费者(顶级肉食动物);分解者主要是微生物,将动植物死体及排遗物中的复杂的有机物分解还原为简单的无机物,供生产者重新利用,它们又称为还原者。从消费食物的角度看,它们也属于广义的消费者。
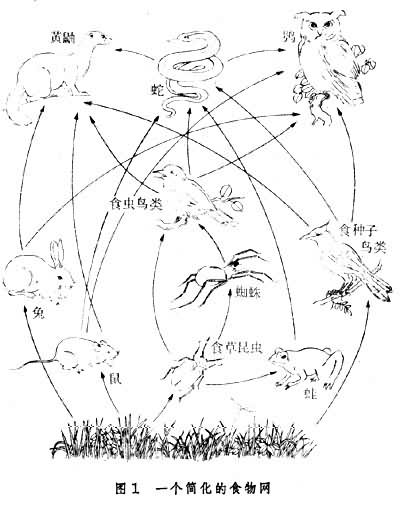
生物之间一物以它物为食的循环递进的连锁关系称为食物链。例如,草原上可以见到牧草→植食动物→肉食动物→大型肉食动物的链锁,这种从植食动物吃活植物体开始的食物链,称为牧食食物链。在森林中,每年新生长的枝叶,只有不到1/10被植食动物消耗,大部分都落在地面被微生物分解。这种从微生物分解动植物死体有机物开始的食物链,称为碎屑食物链。当然,这种食物链不限于森林中,一切动植物尸体及排遗物都要经此途径分解为腐殖质或简单的无机物质。自然界中的许多食物链彼此交织形成复杂的网络状营养结构,称为食物网。
在营养结构中,生产者是系统中其他生物的营养来源,属第一营养级;作为一级消费者的植食动物属第二营养级,依此类推。但实际上对具体物种进行营养分级只能是相对的。例如一个杂食动物可能同时占若干营养级,微生物可能分解生产者和各级消费者的死体和排遗物,所以通常也只笼统称为“次级”消费者而不再细分。
空间结构 草原、森林、湖泊、海洋等自然生态系统在空间上可区分出两个营养层次:上面的吸收阳光进行光合作用的自养层和下面的异养层。自养层,其中生产者的生产过程占优势,在水域中就是日光能透入的光亮层,在陆地上就是植被的株冠。异养层,以动植物死体及排遗物的分解过程为主,在水域中就是无光带和水底的沉积带,在陆地上就是落叶层和土壤的上部。
自然生态系统还有另一种垂直分层,往往是以植物生活型为标准划分的。森林生态系统尤其是热带雨林,具有复杂的垂直分层结构,如乔木层、灌木层、草被层、地被层等。而草原和荒漠生态系统的分层结构就比较简单。
时间结构 由于日光、温度和其他自然因素的昼夜周期变化,生态系统中大部分生物成员的活动表现出昼夜的周期性。例如有的动物白天活动,有的夜间活动,还有的在晨昏活动,这使生物群落的面貌在一昼夜中有明显改变。同时,季节周期性也是大多数生物群落的特征。一年四季,生态系统的结构发生明显而有规律的变化。例如夏季的食物网就与冬季的食物网明显不同。
分类 ∩以根据物质和能量能否跨越其边界而划分为3种基本类型:物质和能量都不能跨越边界的系统称为隔离系统;物质不能跨越边界而能量可以出入的系统称为封闭系统;物质和能量都能自由出入的系统则称为开放系统。如果把整个地球的生物圈看作一个生态系统,则系统中经常有能量出入,但除陨石和火箭等形式外几乎没有物质流动,因此,可将其看成是封闭系统。地球上一切局部的生态系统,不仅在邻近系统间有物质交换,通过大气和河流,物质还可以在全球范围内移动,因此都属于开放系统。
生态系统的范围有大有小。最大的是生物圈,包括地球上的一切生物;小的如一块草地、一个池塘等,甚至还有人把海洋、湖泊中的一滴水也看作一个生态系统。很多密切关联的小系统又可以组合成一个大系统。有时因研究需要,还可以把任意划定的一个范围当作一个系统,研究有关的生态现象。一般说来,一定的生物群落总生存在一定的自然环境内,其中的生态关系也具有相应的特点。因此人们常按生境和植被来划分各类生态系统。例如生物圈中有陆地生态系统、海洋生态系统和淡水生态系统;陆地生态系统中又进一步分为森林、草原、荒漠、冻原等生态系统;森林生态系统又有针叶林、阔叶林、混交林,等等。这样划分的自然生态系统之间常常没有截然的分界线,彼此重叠的区域称为群落交错区。同类系统内部的自然条件、物种组成以及生态关系都是比较相近的,系统与外界间的能流和物质交换有时还有一定的间断性。除自然系统外,还有人工生态系统,如农田、城市、宇宙飞船的密闭舱等。人工系统的边界一般比较明确。由于当今的人类活动几乎影响到生物圈的每个角落,因此自然系统与人工系统之间往往也不能划一条绝对的界线,只是二者受人类影响的程度有所不同而已。有一类人工生态系统是模拟自然系统制造的实验系统,完全由人工控制,具有作为生产者、消费者和分解者的生物,如人工气候室、小宇宙、恒化恒浊装置等。这些系统一般较小,常称做微生态系统。
功能 指系统水平的生命活动,也就是生态系统的整体代谢。一般包括能流、物质循环、稳态调节与信息流三个方面。
能流 生态系统中的食物链实际反映了系统中的能的流动。在森林中,绿色植物以日光为能源,吸收空气中的CO2及土壤中的养分,制造构成自身的生命物质。植食动物吞吃绿色植物,肉食动物又捕杀植食动物,它们都利用食物中的能量维持自身的生命活动。一切动植物死亡后都在一定的条件下被微生物分解。微生物依靠分解所释放的能为生,为解过程又把各种复杂的有机物质还原为绿色植物能重新利用的无机物质。在这个过程中,太阳能是一切生命活动的初始能源,它以化学键的形式在食物链中传递,并在逐级分解过程中释放出来,最后以热的形式散失。这是一个能的单向流动过程。研究能在生态系统中的动态规律,研究它如何从一种形式转变为另一种形式,以及它在流动过程中的数量变化和利用效率等,是生态系统的功能研究的主要内容。
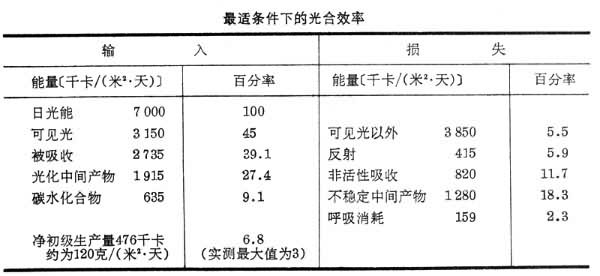
在生态系统中,能由光能经化学能到热能不断地转换形式,但总量不变。因此可以用数学等式来表示。按照热力学第二定律,当能量由一种形式转换为另一种形式时,总有一部分能量转化为热能的而耗散。在生态系统中,植物接收大量光能,但平均只有不到2%的能量转换为生命物质中的化学能;其余的或者反射掉,或者转换为热能而散失。曾有人列举最适条件下的光合效率(见表)。在热带的或温带仲夏的无云天,太阳辐射能的最大输入可达 7000千卡/(米2·天)。但只有39.1%的入射光被吸收,而形成碳水化合物的只有9.1%,相当被吸收光能的23.2%,这就是总初级生产量,再减除植物的呼吸消耗,则净初级生产量只有入射光的6.8%,相当被吸收光的17.4%,约合120克/(米2·天)。这是最大光合效率的估计值,实际测定的最大值只有54克/(米2·天),只占入射光的3.1%,相当被吸收光的7.9%。一般实测值均远低于最高估计值,这说明净初级生产量还有其他限制因素:在陆地有水、湿度、营养物质和植食动物等;在水域,则与水的透明度有关。
植物提供的净初级生产量成为消费者的食源,而前一级消费者又成为后一级的食源。但因种种情况,牧食食物链中传递的能量逐级减少,一部分为各级生物呼吸消耗和用于生长或生殖,一部分转入碎屑食物链为消费者消耗和利用,还有一部分未得利用而暂时堆积地面。
应指出,未被植物固定的能量以及在能流过程中以热能形式逐级散失的能量并不全是浪费。生物需要一定的环境温度才能生存;生态系统的物质循环需要水流、气流来带动。这两者都要由热能来提供。
无论整个生态系统还是某一个营养级,都有一定的现存生物量(见生物生产力)。如果能量输入和输出相平衡,现存生物量稳定不变;如果输入大于输出,则生物量增加,造成积累;如果输出大于输入,则生物量减少,造成亏损。这种动态变化,一般用生态系统的总初级生产量(P)和总群落呼吸量(R)的比值来表示,即:P/R(图2)。
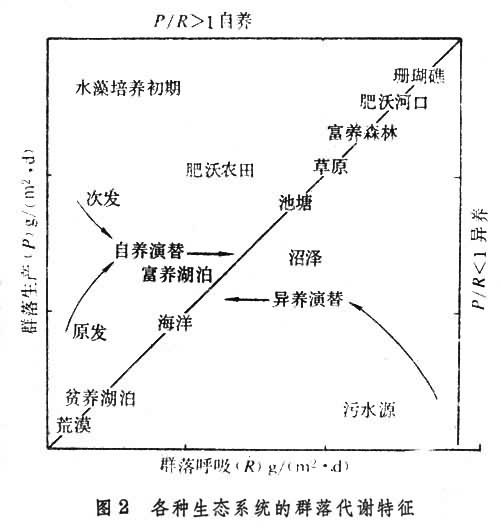
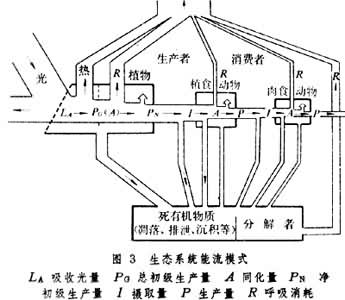
图2表明,一切达到成熟阶段的生态系统,P/R稳定在1上下。在一般生态系统的发展过程中,大都是生产大于消耗P/R>1,表现为群落自养演替;但在污水分解自净过程中,消耗却大于生产P/R<1,表现为群落异养演替。生态系统的能流过程可用图3说明。
物质循环与源自太阳的单向能流不同,生物代谢所需的物质只能取自周围环境,这些物质通过生物的排遗或死体分解又送还到自然界,在有限的范围内反复使用而形成循环。虽然有些气态或水溶性物质可随气流、水流传到远处,但就全球范围来讲物质只能是循环的。化学物质在生物体与环境间的循环称为生物地球化学循环。
现已知道,自然界92种化学元素中有30多种是生命所必需的。其中较为重要且需量较大的有氧、氢、碳、氮、磷、硫等几种。
地球表面的一切物质均以分子状态存在,有气态、液态和固态3种形式。一般说来,地面生物都存在于大气之中,大气可以在大范围内自由流动,因此气态分子最便于生物摄取。水也是以气态飘至内陆的,但降雨后液态水却只能往低处流,很多水溶性物质只能随河流单向入海。固态分子活动性最小,很多要溶成液态后才能被生物吸收。
人们常把物质的循环过程描绘成由一个“库”流动到另一个“库”。例如在碳循环中,气态CO2可看作是一个库,化石燃料中的固态碳又是一个库。对于生物来讲,物质的库首先必须是便于摄取的,例如在地面,气态库是最方便的;其次,库的储量必须足够大才能保证稳定的供应,在这方面,气态库也比较优越,因为大气的流动能使局部的消耗迅速得到补充;第三,库中的物质必须能为生物所利用。
参与物质循环的各种化合物可能含有大小不同的能量。有些化合物在转化过程中放出可为生物利用的能量,但反向的转化却必须依靠外界提供能量。例如,碳水化合物和CO2同为碳的化合物。前者在生物体内分解为CO2和H2O时,可以释放能量供生物利用;CO2在植物体内合成为碳水化合物时却需要阳光提供能量。含氮化合物虽然不是一般动植物的能源,却可能为某些土壤微生物提供能量。生物遗体的分解产物以及动物排泄的含氮废物主要是氮氢化合物,还有氮肥中的铵盐,这些负三价氮的化合物 (N娛、NH3、N嬃)在土壤中很快被硝化细菌氧化为正三价的亚硝酸盐(NO2)或正五价的硝酸盐(N幦)。硝化细菌就依靠氧化过程中释放的能量为生。植物根系吸收硝酸盐后,还必须借助体内呼吸作用提供的能量再将其逐步还原为氨基形式,才能将它用于合成蛋白质。空气中富含的零价氮(N2)也要由固氮菌依靠其他能源将其转变为NH3的形式后, 才能被植物利用。因此,在碳、氮、硫等涉及能级转换的物质循环过程中,都有若干生物环节。
在上述氮的自然界循环过程中处处有微生物的参与。不仅如此,珊瑚礁和热带雨林可能都是因为有以生物为媒介的高效物质循环过程才表现出极高的生产力。在珊瑚礁中,可能是内生藻类为珊瑚虫提供能量并在两者之间形成物质循环;在热带雨林中,根际共生真菌可能直接消化落叶层和土壤腐殖质中的有机物质并经菌丝提供给根系。否则土壤中的可溶性物质将被大雨淋溶丢失。
稳态调节与信息流 稳态调节概念来自个体水平的生理学。例如哺乳动物体内的温度、渗透压、pH值以及各种电解质和营养物的浓度都保持在一个稳定的范围内,这是在其自身神经体液系统调节下,随时抵消各种偏差而实现的。生态系统虽然没有与此类似的调节机制,但也具有一定的抵御环境压力保持平衡状态的能力。例如水体受到轻度污染时能通过理化及生物作用自净,森林经受一定程度的火灾或砍伐后能通过自我更新复原。特别是成熟的生态系统,每年的能量收支大致相等,营养物质循环近于“封闭式”,流失极少,系统能相当长久地保持一定的外观和结构,这些都是稳态调节的结果。
生态系统的稳态调节是通过负反馈机制实现的。例如,种群的个体数量增加并达到一定程度时,食物必然相对短缺,内部竞争随之加剧,从而导致出生率的下降和死亡率的上升,迫使种群的个体重新接近稳定的水平。反之,种群的个体数量减少并达到一定程度时,食物会相对充裕,内部竞争也会减少,这就为个体数量的恢复创造了条件。在这些过程中,数量的增减作为一种信息回输到系统中去,所以这称为“反馈”;反馈的结果与原趋势相反,所以又称为“负反馈”。这种通过信息传输的调节过程也常称为生态系统的信息流。
生态系统的自我调节能力是有一定限度的,超过这一限度将导致系统结构和功能的不可恢复的破坏。例如水体污染超过自净能力,可导致水生生物大量死亡;森林大面积砍伐会造成水土流失、岩石裸露;草原过度放牧能引起荒漠化等等。随着人类社会的发展,这些问题日益严重。为了合理地开发、利用和改造自然资源,有必要深入研究生态系统的调节能力及其机制。
生态系统的发育 生态系统也有一个从幼年期到成熟期的发育过程。表现为:现存生物量增大,物种增多,营养结构和空间结构趋于复杂;P/R趋近于1,物质循环接近“封闭式”,流失极少;系统稳定性增高等。美国生态学家E.P.奥德姆于1969年总结了生态系统发展过程中重要的结构和功能的变化规律,主要内容是:
① 能量学变化:为了便于比较,这里的生物量都折算为能量值。P为总初级生产量,即总光合量;R为群落呼吸量,即维持系统运转的能量消耗;E为单位能流量。在发展期中,一般是P大于R(P/R>1),因此现存生物量B逐渐积累增加(自养演替)。只有在个别情况如有机废物污染水系时,R大于P(P/R<1),使现存生物量逐步减少(异养演替)。但两者演变的趋势都是P/R趋向于1,即系统能量收支相抵,净生产量减至最低,这是成熟期的最重要标志。因现存生物量B的增大,所以生产量的相对数值P/B减少,而单位能流所能维持的现存生物量B/E增加。
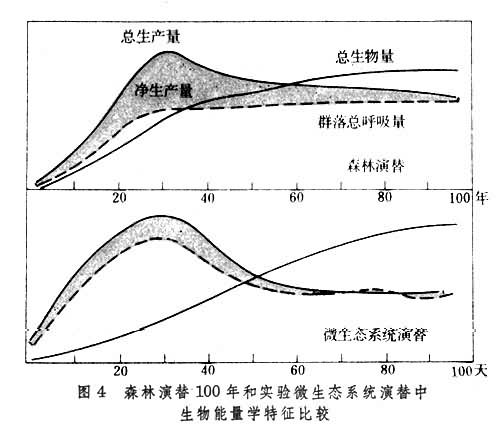
发展期中,食物链较简单,常为直线状,主要为牧食食物链。到成熟期,食物链多形成复杂网状结构,大部分能量通过碎屑食物链。
② 群落结构及物种生活史:随着系统的发育,物种多样性常趋向增高,某一物种或小类群占优势的情形减少。但至顶极期多样性指数也有下降的情况。一方面,生物个体增大,生活史延长并趋向复杂以及竞争加强等因素,都可能减少物种的多样性;另一方面,群落生物量的增加和结构的复杂化又可能提供更多的生态位,从而有利于多样性的增高。物种多样性指标的最终数值取决于上述两种倾向的对比。
近来的研究发现,在系统发育过程中,生物排泄到环境中的代谢产物种类大量增加(生化多样性)。这些产物不一定是废物,可能对邻近生物的生长和繁殖具有抑制或促进作用,从而调节生态系统的组成和发育(见生物种间化学交互作用)。
③ 营养物质循环:在系统发育过程中,物质循环趋向“封闭”,物质主要经过生物途径,因而很少丢失。
④ 选择压力:由生物进化的角度来看,在系统发育早期,繁殖多、生长快的物种较易生存;至晚期,竞争性强的物种占据优势。整个发育过程中,由r选择转向K选择,以量取胜的对策让位于以质取胜和自我调节的策略(见生态策略)。
⑤ 稳态:成熟期的一个特点是物种间的联系更加密切,共生、捕食、寄生等关系错综复杂,系统也更加稳定。物质在系统内部循环很少丢失。群体结构高度有序,即信息量较高。
总之,生态系统的发育过程遵循着一定的程序。这种规律究竟取决于生物群落的内在因素还是环境因素,或者是两者交互作用的结果,目前还没有一致的看法。有人把生物群落视为超级个体,强调生物因素。但个体发育蓝图载于基因结构之中,群体并无相应结构。另一极端则视群落为物种的随机聚合,每个物种单独受环境制约,物种间交互作用决定着整体功能。看来还需要更多的研究才能得出较成熟的结论。
生态系统发育的理论有着重要的现实意义。自然生态系统的发展趋势是最大限度地维持大生物量、多生物种和复杂结构,在成熟系统中能量主要用于维持呼吸消耗,所以净生产量很小。而人工生态系统(如农林业生产系统)则力求提供最高生产量,同时使积聚在系统中的生物量维持在最小数值,即追求高K/B比值。世界上流行的做法是,保持系统处于演替早期并采用单种栽培。这样做的结果牺牲了系统的稳定性,例如单种栽培特别易受虫害。更重要的是,人类需要新鲜的空气、清洁的用水、适宜的气候等等,这一切都要求地面上保留足够的生产力不高的植被。所以,人类生活不仅需要充足的衣食供应,还需要美好的环境,能够满足这些要求的是包括农田以及森林、湖泊、海滨等多种自然生态系统在内的组合体。
参考书目
E.P.Odum,Fundamentals of Ecology,3rd ed.,W.B.Saunders Co., Philadelphia, 1971.
E.J.Kormondy, Concept of Ecology, 2nd ed.,Prentice-Hall,Inc., Englewood,Cliffs,New Jersey,1976.
R.H.Whittaker,Communities and Ecosystems,2nd ed.,Macmillan Pub. Co., New York,1975.