行动(卷名:生物学)
locomotion
生物在空间的整体移动行为,例如动物的步行、奔跑、游泳、飞翔等。行动指主动的移动,不包括被气流、水流或其他生物带动所造成的空间移动(如寄生虫随寄主在空间的移动)。但有时主动和被动的界限不明确,如某些蜘蛛放出一段蛛丝后,乘丝随风转移到其他地方,或蝗虫主动起飞后却被强大气流带至远方,两者都有一个准备阶段是主动行为,而具体的转移却是被动的。除阿米巴之类生物无法说出它们的哪一部分负责行动之外,大多数能行动的生物都演化出专门的行动结构,包括细菌的鞭毛直到高等动物的足、鳍、翼等行动器官。多种细菌和原生动物都能行动,但在多细胞生物中,行动主要见于动物。除某些营固定附着生活的低等动物外,大部分动物都有特化的行动器官,它们的取食、生殖等基本生活活动也都要通过行动才能实现。就行动的范围、速度、单程的距离以及定向的准确性而言,以鱼类、哺乳类(包括鲸等海兽)和鸟类最为突出,因此,它们分别在水陆空3界成为优势生物。
介质与基底 大多数生物生活在水介质或空气介质里。水的密度较高,一部分比重与水相近的生物经常浮在水中,而比重高于水的就只能在水底上行动。在空气中,只有微生物可经常飘浮。所谓飞行生物只是它们的主要行动在空中,一方面它们要在飞行器官发育成熟后才能飞行,而且仅仅维持浮在空中也需要耗能,所以它们平时休息、产卵、育幼等等活动都要在地面上进行。因此除了浮在水中的动物外,大多数动物都要在基底(陆地及水底)上行动。此外还有少数比重轻于水的水生生物漂浮在气水界面上,另有一些地下动物生活于地下穴道中,一些底栖动物隐藏于泥沙中,不过后两者也常活动在地面或水底之上。
适应行动的需要,较高等的动物都呈两侧对称的体型,就行动方向而言有头尾之分,相对于重力方向则分为上下。而且较高等的动物都演化出专供行动的附肢如足、鳍、翼等,在基底上行动的动物借附肢支持和带动身体。附肢使动物的行动更加迅速和灵活,但也带来稳定性的问题,动物必须维持正确的姿势才能行动。浮在介质中行动的动物还存在一个浮力问题。无论是水生生物还是飞行动物的比重都比一般陆生动物轻。有的水生动物的骨骼无钙盐沉积(软骨鱼)或由几丁质构成(乌贼),有的水生动物体液中的较重离子如Mg2+及SO厈等为较轻离子所代替。有的水生动物体内富含油脂等增加浮力的物质,例如鲨鱼的肝特别大,肝中含有一种较大多数油脂为轻的鲨鱼脂。另一些水生动物则在体内特殊部位中储气,乌贼骨中全是微小的气室,其中充满气体,很多硬骨鱼则具有气鳔。全部软骨鱼及许多硬骨鱼是靠运动来产生升力。在飞行动物中,昆虫体小而轻,鸟类的钙质骨像乌贼骨一样具气室,但鸟的比重仍远大于空气。它们必须借助飞翼的运动保持足够升力。有时它们可以利用一定的地形和气候条件下的上升气流升空,但这只能作为一种辅助手段。此外,处于多变的水、气流中的水生及飞行动物还必须保持良好的平衡,因此他们要依靠种种平衡器官,如果蝇由后翼演化成的平衡棒和脊椎动物的前庭器官。
动物向前行进,一方面要靠躯干或附肢与基底或介质相互作用以产生推进力,另一方面还要克服介质对前进的阻力。地面坚实,以足蹬地推进的效率最高,但在地下钻行阻力也最大。在水中前进阻力要比在土中小得多,但划水前进所获得的动力也要低得多。单细胞生物如草履虫可靠其表面的纤毛拨水前进,这是因为水有一定的粘性,微小的纤毛拨水类似以足蹬地,大量纤毛产生的推进力足以推动草履虫前进。但对于重量大得多的鱼类来说,水的粘性便远远不够,而要依靠水的惯性力,片状的鱼鳍把大量水拨向后方从而将本身反推向前方。水生动物的比重与水相近,故它们无需消耗多少能量以产生升力。但要飞翔,飞行动物便要既克服阻力又同时产生升力。空气的阻力最小,但因其密度过小故必须向后拨动大量空气才能产生足够的推进力。飞机的螺旋桨产生推力带动机身前进,在空气中前进的机翼则提供升力。动物的飞翼把两者结合在一起,同时产生推力和升力。此外,快速行进的水生动物和飞行动物都具有流线形的体型,以减少介质的阻力。
运动的机制 主要有3种:变形运动、鞭毛和纤毛运动、肌肉运动。前两者主要见于微小生物,后者见于大型动物。
变形运动 是一种细胞运动,只有单细胞生物能采用这种行动方式,例如原生动物阿米巴类和粘菌。行动中细胞不断变形,先向前伸出伪足,继而后面的细胞质流入伪足使伪足扩大,逐渐变成生物的主体,继续向前行进时再伸出新的伪足。变形运动属于借基底行动的方式。伪足必须固着于外界物体上才能带动整体前进。变形运动的速度异常缓慢。
鞭毛运动和纤毛运动 部分细菌有鞭毛,但与真核生物的不同,较细且短,全靠基部的力量旋转运动。真核生物中,原生动物的鞭毛纲和纤毛纲分别利用鞭毛和纤毛来行动,某些小型扁形动物和软体动物也用纤毛在水底滑行。鞭毛和纤毛的构造基本一致,毛内有11根微管,中心两根,周围包绕着9根。这种9+2的格局普遍存在于动物界;绝大多数的动物精子以及脊椎动物的各种纤毛上皮细胞也如此。鞭毛与纤毛的不同只在于它们的运动方式:鞭毛位于动物的头侧,鞭毛各部分顺序摆动形成一个自前而后的波浪,使动物体沿鞭毛长轴方向前进;纤毛动物则体覆大量纤毛,每根纤毛的动作如单臂划水,各纤毛互相配合使水沿体表向后移动,因此动物前进的方向与纤毛呈直角正交。鞭毛和纤毛是特化的行动器官,它可使动物在液体介质中灵活行动,速度较变形运动快得多。
肌肉收缩运动 是一切大型生物的主要运动机制(见彩图)。在行动时,身体的某一点(支点)固着于基底(如足固着于地面)或推动周围介质(如鳍或翼推动水或空气),通过肌肉收缩使身体的其他部分(动点)与这个支点间发生相对运动。 条形肌肉的两端必须联结在坚实的组织上,这种组织要能承受肌肉的收缩力并将力传给支点和动点。这种组织在高等无脊椎动物是外骨骼,在脊椎动物则为内骨骼。再一种情况是环状肌肉组成的管腔中充满液体,肌肉收缩加力在液体上,通过液体传力。这主要见于没有骨骼的低等无脊椎动物,如蚯蚓躯体的伸缩,海星管足的移动,水母、扇贝和章鱼的后向喷水等都属于这种情况。跳蛛后肢无伸肌,但它的血压却异常地高(400毫米汞柱),高压血液突然涌入后肢内造成快速的伸直运动,可使微小的跳蛛跃出10厘米,其行动的动力间接来自心脏肌肉的收缩。



高等动物行动时,足、鳍、翼都是通过颉颃肌的交替收缩而作屈伸动作。每个单独屈伸动作的动能有一部分转化为肌腱等组织的弹性位能,在下次反向运动时再转回成为动能。昆虫的外骨骼富有弹性,蚊、蝇、蜂的双翅和垂直及水平两组肌肉分别联在外骨骼的不同点上,两者不直接相联。这些肌肉具备一个特点:被牵张时可引起迅速收缩。垂直肌收缩即引起胸部变形,造成双翅上扬和垂直肌本身的张力突然丧失,但胸廓变形却牵张水平肌而引起它的迅速收缩,这个收缩又引起胸廓的反向变形,造成双翅下扇和水平肌本身的张力丧失。这样,外骨骼和肌肉就形成一个振荡器,动能和弹性能交互转化,肌肉提供的能量则用以克服气动阻力维持振荡不致衰减。因此搧翅的频率只决定于振荡器本身,每秒钟百余以至近千下,而肌肉接受的神经冲动频率一秒不过数十。因而这些昆虫的振翅动作的起动是受神经支配的,但高频振荡却主要是由振荡器的机械作用来维持的。
肌肉纤维中含有大量平行排列的蛋白质微丝。微丝有两种,粗微丝由肌浆球蛋白构成,细微丝含肌动蛋白等。粗细微丝交错搭接,当两者交互滑行使肌纤维缩短乃形成肌肉收缩。 鞭毛和纤毛内的9+2微管其化学成分与肌肉不同,但有证据说明,它们也是靠微管间滑行来运动。只是因微管长度固定,故管间滑行只能造成一定的侧弯运动,从而造成波浪式运动或划水式运动。变形运动的分子机理还不清楚,但一切真核细胞中都发现有肌动蛋白和肌浆球蛋白,说明这一切运动机制可能都有相近的分子基础。
行动的类型 不同的物种为了适应相同的生态条件发展出类似的器官,例如细菌和原生动物的鞭毛、昆虫翅和鸟翼等等。这些功能上相近但非同源的器官称为同功器官。因为在进化阶梯上处于不同地位的生物,面对同样的功能要求,只能是改造利用自己原有的结构。所以这些同功器官结构不同,效能也各异。
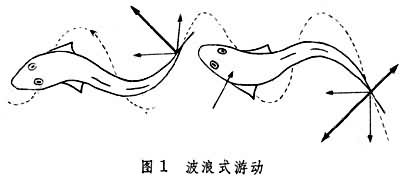
游动 上面介绍过几种微小水生生物的行动方式:变形运动和鞭(纤)毛运动。但在动荡的水体中,它们基本是随水流被动地漂移。在水中能主动游动甚至逆流而行的生物,只能依靠肌肉收缩,并主要采取4种行动方式。①波浪式游动:鳗鲡、海蛇等细长型动物在游动时,身体左右移动形成一个由头向尾传播的波浪形运动,其作用类似摇橹驶船(图1)。这种方式速度不高。②水翼式游动:大型快速游泳的动物如金枪鱼等均采用这种方式,它们左右摆动其尾鳍推水前进,作用也像摇橹。尾鳍产生推力的原理类似机翼,属于小翼类型。因鱼尾面积大且摆动有力,故速度颇快。至于胸、肤、背、臀等鳍的功能主要是维持平衡。海兽如鲸的尾叶呈水平位,上下摆动产生强大推力。海龟的鳍状前肢和企鹅的鳍状翼的工作原理也是如此。③喷水式游动:水母、章鱼均利用向后喷水前进。扇贝平时栖居水底,行动时猛然关闭双壳将水外挤。因其断面轮廓像机翼断面,且喷水方向偏下,故扇贝向斜上方漂去。④划水式游动:大多数水禽后肢带蹼,在水面游动时左右交替划动,潜水时则双侧一同蹬水。蛙类和淡水龟也都用带蹼后肢划水。
地面行动 除了某些小昆虫因体轻可利用水的表面张力在水面上行走之外,绝大多数陆生动物和底栖动物都是在地面上行动。底栖动物与陆生动物不同处在于,水的阻力大,行动不能很快,但水浮力却减轻了体重负荷,因而虾、蟹在水中行动比陆上还快些。有些小扁形动物和小软体动物也可以利用腹面纤毛在底面滑行。这些小动物在腹面还分泌一层粘液将身体垫离底面以容纤毛运动。所有地面行动可分为蠕行和走行两个基本类型。
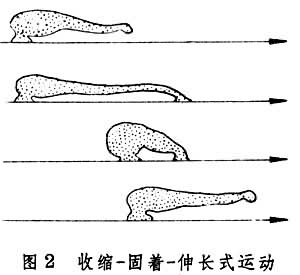
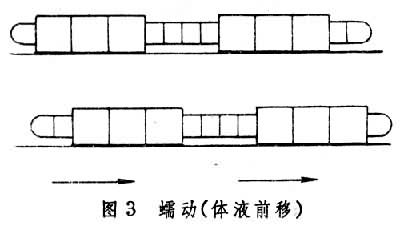
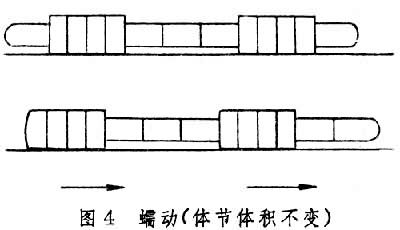
蠕行见于多种管型动物如蠕虫和蛇,它们借躯体的伸缩、屈伸或摇摆向前行进,行进时部分身体紧贴地面。蛭的身体前后两端各有一个吸盘,如果开始时用后吸盘固着地面,便先向前伸长躯体并使前吸盘附着地面,然后以前吸盘为固着点,躯体收缩带动后吸盘前移,然后再以后吸盘为固着点重复上述行动。这种以一点固着地面,或收缩后面部分,或伸长前面部分的前进方式常被称为“收缩-固着-伸长”式行动(图2)。这种行进方式只见于体壁柔软的蠕虫,因无骨骼限制,故可借助纵肌和环肌变换体形,体腔内液体起传力作用。再如Polyphysia属的海生蠕虫的体腔并不分隔为小室,故体腔液容易在体腔内流动。行动时身体一部分胀大固着地面,位于膨胀部前面的身体部分伸长,膨胀部后面的身体部分收缩,结果体液前移使胀大部分也前移(图3)。在这种运动中,收缩波也向前传播且速度超过整体前进速度。但更常见的是蚯蚓式的蠕动(图4)。因为蚯蚓身体被隔膜分为多个体节,体液只能在每个体节内流动,每个体节的体积恒定,纵肌收缩时体节变得短而粗,环形肌收缩时体节变得长而细。短而粗的部分压强较大乃固着地面,细而长的部分抬离地面。固着部分前面的体节伸长变细,后面的体节缩短转粗,在收缩波向后传播的同时整体却在前进。蛇有脊椎骨,故身体长度无法改变,它躯体一侧的纵肌收缩,该侧躯体便侧弯,而对侧舒张,一张一弛形成波浪,波浪式的运动从头到尾传递。当收缩的躯体遇地面上可凭依的物体时,它便借侧推力前进。
走行指依靠附肢在地面行动,见于节肢动物和陆生脊椎动物。附肢具关节,有助于灵活运动,而附肢将身体抬离地面才使快速运动成为可能。这样,附肢不仅是行动器官,还要支持身体;不论如何行动,身体重心须保持在附肢支持的范围内。对小昆虫来说,微风也可能将其吹倒,故其附肢伸展面积远超过身体投影面积。附肢数多,有助于稳定,但不利发展速度。昆虫6足,蜘蛛8足,而多足纲常达几十足。陆生脊椎动物多为4足,而当其快速奔跑时,保持着地的足数常不超过2足,甚至有4足腾空的阶段。开阔草原上的大型植食动物如斑马,长途跋涉于水草之间,遇敌只能靠奔跑逃生。它们附肢的远端细长,以特化的趾端(蹄)着地。减少远端重量有助增加前后摆动的速度;肌肉集中在近端也是牺牲力量而求速度的表现。某些大型肉食动物也相应加快了速度,不过它们以爪着地,爪有利捕食或攀缘。猎豹在奔跑时,背部也有屈伸运动,伸时有助增加步长。借助躯干运动来增长步长的还有蝾螈(两栖类)和蜥蜴(爬行类),它们的四肢位于躯体两侧,因此躯干向一侧侧弯叶可大大地加大另一侧附肢间的距离。
双足行动稳定性差,但却有一定速度,如某些蜥蜴迅速奔跑时两只前足离地,身体几乎直立。鸟类和人均为双足行动,前肢可从事其他活动(飞行和劳动);两者的足掌较大,增加了稳定性。在鸟类,双足行动常只是一种辅助方式,但某些走禽如鸵鸟的奔跑速度可达每小时60公里。跳行也应属于双足行动的一类,蛙、袋鼠、兔和跳鼠都具4足,但后肢长度几近前肢的两倍,跳跃动力全来自后肢。跳跃时后肢同时蹬地,继之为腾空阶段,落地时以前足(兔)或仍以后足(袋鼠)着地。跳跃距离主要决定于起跳速度和前肢角度。蚤(无脊椎动物)的跳跃距离可达身长的50倍,这要求极高的初速(远超过蚤肌肉收缩的速度)。事实上,蚤的后肢基部有一块弹性组织,借助肌肉运动预先使其变形,跳跃时储存的弹性位能突然释放出来,将蚤体弹射出去。
地下行动 泥沙阻力较小,而在土壤中行动却需要很大的力量和时间。蠕虫主要靠蠕动方式将细长的头节插入土隙,继之头节膨胀,扩大土隙,固定后再拖曳后面各节前进。两栖类和爬行类也以躯干钻行,但哺乳类却靠附肢掘穴。终生栖息地下的鼹鼠以强大的前肢掏土,将土压实在两侧壁上。至地面寻食的草原犬鼠则自身下将土排至穴外。
攀援 节肢动物体小而轻,故不需特殊结构以助攀援,但营攀援生活的脊椎动物都发展出各种有利附着抓握的器官。树蛙4趾端扩大如盘状,蜥蜴则具爪或鳞。啄木鸟以尾羽协助利爪,3点支撑附立树面。鼯鼠亦具利爪,且因其后足可转向抓握故可以头向下的方式奔跑。猿猴则发展出能抓握的长趾,有的还具长尾盘绕树枝以稳定身体,长臂猿则可在树间悠荡,特称臂行。
飞行 滑翔可以认为是飞行的雏型。有少数可以滑翔的蛙、蛇、蜥蜴、鼯鼠,它们生活在树丛中,常自高处下跃,滑行至低处或地面。它们在滑行时伸展四肢以增加空气阻力。例如,有的蜥蜴具长肋,展开后呈扇状;有的鼯鼠前后肢间有肉趾;飞蛇在滑行时身体强直,肋骨向外伸展到极限,以获得最大的体表面积,并收缩其腹部正中部分形成一个向下的凹面,以增加浮力。飞鱼的行动也近于滑翔,当在水内加速窜出水面时,翼状胸鳍和臀鳍相继外展,借风浮起,可滑行数十米。
真正的扑翼飞行只见于昆虫、鸟类和哺乳类(蝙蝠)。昆虫翅为几丁质,借褶皱结构及翅脉增加刚性。昆虫体轻不耐风力,但动作异常灵活。鸟翼系前肢演化而来,肢骨管状中空储气,故既坚固又轻,翼本身由羽毛构成。蝙蝠指骨极长,指骨末端至肱骨、体侧、后肢及尾间连以薄而柔韧的皮膜。扑翼飞行的关键在于,下扇时翼面积增大产生足够的前进推力和升力,但上扬时必须改变翼与气流的角度或缩小翼面积以避免产生反方向的力量抵消原有效果。蜂鸟还可悬停采蜜,这时鸟身直立,双翼前后翻飞只产生升力,这种飞翔耗能较大。此外,鸟类也常利用气流滑翔。例如山坡常有沿坡上升的气流,日照下地表也常有热流上升,鹰隼等猛禽即借此热流在空中翱翔觅食,这称为静力性翱翔。另一种是动力性翱翔,见于海鸟。在大洋中经常有平行海面的气流且越高风速越大,海鸟借风力螺旋前进。海鸟顺风下滑逐渐增速,接近海面时再上转,借已积累的动能逆较弱的海面风上升,随着上升所遇风速越来越大,原积累的动能也逐渐消失,至30~40米高再重复下滑,这样每次可前移一段距离。在整个过程中也适当辅以扑翼来补偿能量消耗(图5)。

行动的进化 真核生物研究得比较多。从分子水平上来讲,肌动蛋白和肌浆球蛋白是一切真核生物运动的基础。细胞质的定向流动及相变造成变形运动。运动蛋白集束成特定结构乃有鞭毛和纤毛运动,纤毛行动较为进化,因为各纤毛的运动必须协调配合。多细胞生物沿着3条路线向前发展。营光合作用的植物固着地面,以叶面固定光能,以根系吸取液态营养。靠吸收获取营养的真菌,或腐生或寄生也营固着生活。这两者的生殖细胞借外力传播,完成有性生殖。只有到了动物,才逐渐发展出完备的运动器官,觅食、求偶等基本生活活动全在行动中完成。
动物中出现特化的运动组织肌肉,而一切具有一定行动速度的动物都采取了长管状的体型,主要感觉器官和进食器官集中于前部。早期无脊椎动物的体壁柔软但缺乏坚实的支持组织,体壁上的纵肌和环肌只能作用在体液上,因此有人称体液为“液压骨骼”,动物靠“收缩-固着-伸长”的方式前进。坚实骨骼的出现是一个巨大进化,但骨骼限制了躯干的伸缩,动物转而以附肢行动。带关节的附肢将躯干抬离地面,加快了行动的速度和灵活性。目前在陆地上的动物有两支:无脊椎动物中的一部分昆虫和脊椎动物中的哺乳类。而活跃于天空的主要是飞行昆虫和脊椎动物中的鸟类。
就行动的速度和单程行动的持续时间及距离而言,鸟类达到最高峰,单位距离所需能量也以鸟类最低,但却不能说鸟类便是最进化的物种。事实上,不同的介质中采用不同的行动方式无非是各种动物在地球上开辟新生态位的过程。飞翔万里的候鸟不过是占据了其他动物难以利用的某些高纬度的季节波动较大的资源。动物不断地行动,目的不外觅食、求偶、避敌、趋温等。当这些目的易于满足时,还可能出现退化性变化,如某些寄生虫丧失了行动能力。再如海岛及高山上的昆虫易被大风吹失,故它们不复飞翔。另一方面,鸟类中有善游的企鹅和善跑的鸵鸟,两者也丧失了飞翔能力;哺乳动物中鲸和蝙蝠却分别发展出极高的游泳和飞翔的能力。这种转变使它们能利用一些同类动物不能利用的生态位。
完全变态的昆虫在发育的不同阶段采取不同的行动方式。终日取食的幼虫在植物上爬行,四出求偶的成虫则在空中飞翔。在行动的进化上可以见到各种高度发达的同功器官,无论哪种器官,只要能适应环境,都表明了进化的成功。飞行可说是最复杂的行动方式,动作的协调、姿式的稳定和准确的定向能力等等都要求神经系统高度发达。昆虫和鸟类的飞翔行为很多是本能的,所以幼体出生后便能独立生活。还有人认为,正因为飞行动物的神经结构都用于从事复杂的定型活动,所以它们未能演化出像高等哺乳动物那样灵活多变的智力行为。