遗传密码(卷名:生物学)
genetic code
决定蛋白质中氨基酸顺序的核苷酸顺序,由3个连续的核苷酸组成的密码子所构成。由于脱氧核糖核酸(DNA)双链中一般只有一条单链(称为有义链或编码链)被转录为信使核糖核酸(mRNA),而另一条单链(称为反义链)则不被转录,所以即使对于以双链DNA作为遗传物质的生物来讲,密码也用核糖核酸(RNA)中的核苷酸顺序而不用DNA中的脱氧核苷酸顺序表示。
在转移核糖核酸(tRNA)分子中有一组与mRNA中的密码子配对的三联体,称为反密码子。每种tRNA携带一种特定的氨基酸,在遗传密码的解读中起着关键性的作用。
简史 1961年英国分子生物学家F.H.C.克里克等在大肠杆菌(Escherichia coli)噬菌体T4中用遗传学方法证明密码子由三个连续的核苷酸所组成。他们用原黄素作为诱变剂处理噬菌体T4的野生型,从中获得噬菌斑较野生型为大的快速溶菌突变型 rⅡ。这种突变型可用原黄素再度处理而成为具有野生型表型的回复体。将回复体和原来的野生型杂交,发现子代中出现一些rⅡ突变型,说明这些回复体实际上是基因内抑制的结果(见抑制基因)。由于原黄素诱发的是移码突变,也即是由于个别核苷酸的增加(+)或缺失(-)而导致的突变,因此可以把rⅡ突变写作+(也可写作-),把基因内抑制突变写作-(或写作+),于是由于基因内抑制而出现的回复体便可写作+-(或-+)。通过基因重组还可以得到种种组合如++、--、---等(表1)。不管密码子由几个核苷酸组成,只要有一个核苷酸的增加(或减少)都会使这一位置以后的密码意义发生错误而成为突变型;而另一个核苷酸的减少(或增加)则可以校正后一核苷酸位置以后的码组,从而使表型恢复为野生型。从表1 的实验结果可以看到在除了 +-以外的各种组合中只有 +++或---或------等组合的表型是野生型。这一实验结果只能解释为密码子是由三个(或三的倍数)核苷酸组成。

美国生物化学家M.W.尼伦伯格等从1961年开始用生物化学方法进行解码研究。他们用人工合成的多聚核苷酸作为信使,用来代替大肠杆菌无细胞系统中原来的内源mRNA进行离体的肽链合成实验,发现以多聚U(U表示尿嘧啶核苷酸)作为信使则只有标记的苯丙氨酸参入而合成多聚苯丙氨酸;以多聚A(A表示腺嘌呤核苷酸)作为信使则产物是多聚赖氨酸。这些实验结果表明苯丙氨酸的密码子可能是UUU,赖氨酸的密码子可能是AAA。与此同时,美国生物化学家H.G.科拉纳用共聚的多核苷酸如 UGUGUGUG……(G表示鸟嘌呤核苷酸)作为人工信使进行类似的实验,发现产物是缬氨酸和半胱氨酸的多聚体,说明缬氨酸的密码子可能是GUG,但也可能是UGU,半胱氨酸的密码子可能是UGU,也可能是GUG。以后的实验确证缬氨酸的密码子是GUG,半胱氨酸的密码子是UGU。
1964年尼伦伯格等人进行人工合成的三核苷酸和氨基酰-tRNA、核糖体三者的结合试验,证明三核苷酸已经具备信使的作用。 例如三核苷酸是UUU时只能和苯丙氨酰-tRNA结合;三核苷酸是AAA时只能和赖氨酰-tRNA结合。二核苷酸则没有作用。
通过上述的种种实验,遗传密码已于1966年全部阐明(表2)。
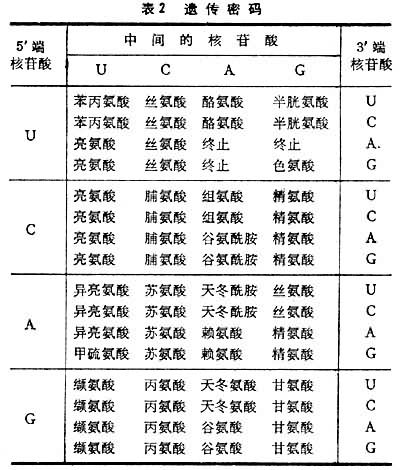
密码表中的左边是密码子5′端的第一个核苷酸,中间是密码子的第二个核苷酸,右边是密码子的第三个核苷酸。例如苯丙氨酸的密码子是UUU或UUC(C表示胞嘧啶核苷酸)。表2中所列的64个密码子编码18种氨基酸和两种酰胺。至于胱氨酸、羟脯氨酸、羟赖氨酸等氨基酸则都是在肽链合成后再行加工而成的。64个密码子中还包括三个不编码任何氨基酸的终止密码子,它们是UAA、UAG、UGA。 这种由三个连续的核苷酸组成的密码称为三联体密码。
特性 无逗号 密码子与密码子之间没有逗号。除了相当于多肽链羧基末端的终止密码子作为翻译结束的讯号之外,密码子与密码子之间没有任何不编码的核苷酸。翻译从一个特定的起始密码子AUG(极少数为GUG)开始,然后按连续的码组沿mRNA多核苷酸链由5′到3′方向进行。直到终止讯号 UAA、UAG或UGA处翻译便自然停止。
不重叠性 在多核苷酸链上任何两个相邻的密码子不共用任何核苷酸。例如在一条由九个核苷酸所组成的多核苷酸链CAGCAGCAG中,如果每一个三联体CAG分别决定肽链中邻接的一个氨基酸则称为不重叠;如果CAG决定第一个氨基酸,AGC决定第二个氨基酸,GCA又决定第三个氨基酸等等则称为重叠。在核苷酸顺序中迄今为止还没有发现过这样的重叠现象。
简并性 64个密码子中除甲硫氨酸和色氨酸外,一种氨基酸往往可由几种密码子代表。例如UUU和UUC都代表苯丙氨酸;又如代表丝氨酸、脯氨酸的密码子各有四种之多。这种密码的同义性称为简并性,同义的密码便称为简并密码。
1966 年克里克提出摆动假说来解释简并性。 认为mRNA上的密码子与氨基酰 -tRNA上的反密码子之间的碱基配对除了A对U和C对G的标准方式外, 反密码子5′端的碱基与密码子 3′端的碱基配对时也可以不按标准方式,例如反密码子上的U既可以与密码子上的A配对,也可以与G配对。又如酵母菌的丙氨酰-tRNA上的反密码子是5'IGC3',丙氨酸的三种密码GCU、GCC和 GCA中的第三位核苷酸U、C和A都可以和I(次黄嘌呤)配对。
统一性 表2中所列的密码子大多是用大肠杆菌作为研究对象得来的。以后陆续发现不论是烟草花叶病毒外壳蛋白或大肠杆菌的色氨酸合成酶或人血红蛋白等的突变型中的氨基酸置换都可以用同样的密码子来解释。在离体的系统中大肠杆菌抽提液可以把兔血红蛋白的mRNA作为模板合成兔血红蛋白。此外还有人将兔血红蛋白的mRNA注射到蛙卵中,蛙卵中也出现了兔血红蛋白。70年代以来用遗传工程的手段还可以将真核生物的基因连接在基因载体上并引入细菌,使细菌产生相应的蛋白质(见重组体DNA技术)。以上事实都足以证明密码的统一性。可是1979年以后对线粒体的密码子的深入研究结果表明遗传密码的统一性并不适用于线粒体。例如在人和酵母的线粒体中UGA都不是终止密码子而是色氨酸密码子;在人的线粒体中 AUA不是异亮氨酸密码子而是甲硫氨酸的密码子。遗传密码的演变是分子进化研究中令人感到兴趣的一个问题。