snake
reptile
Introduction
also called serpent
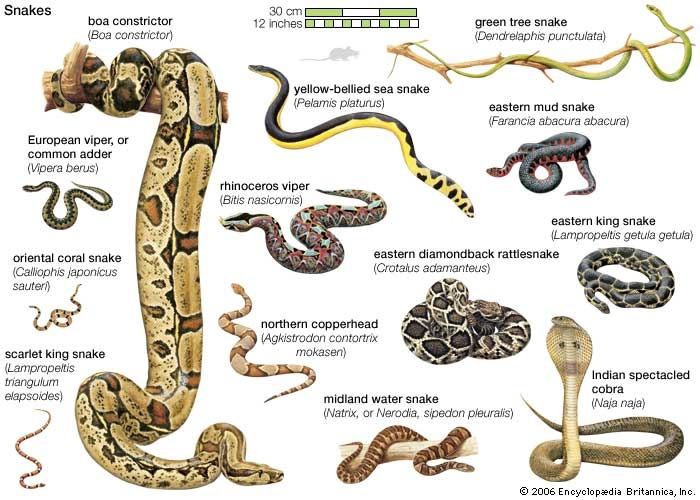 any of about 2,900 species of reptiles distinguished by their limbless condition and greatly elongated body and tail. Classified with lizards in the order Squamata, snakes represent a lizard that, over the course of evolution, has undergone structural reduction, simplification, and loss as well as specialization. All snakes lack external limbs, but not all legless reptiles are snakes. Certain burrowing lizards may have only front or hind limbs or be completely legless. Unlike lizards, snakes lack movable eyelids, which results in a continuous and often disconcerting stare. Snakes also lack external ear openings. Internally, they have lost the urinary bladder. The visceral organs are elongated, with reduction of the left member in relation to the right; the left lung is greatly reduced or even lost entirely. However, snakes possess increased numbers of vertebrae and have developed two novelties among vertebrates: a tracheal lung in the neck region and a venom-conducting system for subduing prey.
any of about 2,900 species of reptiles distinguished by their limbless condition and greatly elongated body and tail. Classified with lizards in the order Squamata, snakes represent a lizard that, over the course of evolution, has undergone structural reduction, simplification, and loss as well as specialization. All snakes lack external limbs, but not all legless reptiles are snakes. Certain burrowing lizards may have only front or hind limbs or be completely legless. Unlike lizards, snakes lack movable eyelids, which results in a continuous and often disconcerting stare. Snakes also lack external ear openings. Internally, they have lost the urinary bladder. The visceral organs are elongated, with reduction of the left member in relation to the right; the left lung is greatly reduced or even lost entirely. However, snakes possess increased numbers of vertebrae and have developed two novelties among vertebrates: a tracheal lung in the neck region and a venom-conducting system for subduing prey.Snakes and man
Snakes are a misunderstood and often maligned group of animals, primarily out of ignorance about their true nature and position in the natural world. All snakes are predators, but venomous snakes have given an inaccurate reputation to the entire group, as most people cannot tell the dangerous from the harmless. Only a small percentage (fewer than 300 species) are venomous, and of those only about half are capable of inflicting a lethal bite. Although snakebite mortality worldwide is estimated at 30,000–40,000 people per year, the majority of deaths (25,000–35,000) occur in Southeast Asia, owing principally to poor medical treatment, malnutrition of victims, and a large number of venomous species. Although there are 8,000 venomous snakebites per year in the United States, the average number of annual fatalities is only a dozen or so per year—fewer than are attributed to bee stings and lightning strikes. In Mexico, 10 times as many people die annually from bee stings as from snakebites.
 Snakes can control the amount of venom they inject and may bite aggressively for food or defensively for protection. Snakes have a limited amount of venom available at any given time and do not want to waste it on nonprey organisms. As a result, about 40 percent of bites suffered by humans are defensive in nature and “dry” (without envenomation). Statistics show that the vast majority of snakebites occur while either catching and handling captive snakes or trying to molest or kill wild ones. In either case, the snake is only defending itself. rattlesnakes, for example, are venomous, and large ones are quite dangerous owing to the amount of venom they can inject. However, most are shy and retreating, and none will attack a person unmolested. When approached or molested, they will coil up and rattle as a warning to be left alone, striking only as a last resort. Most cases of reputed snake attack are based upon encroachment by a person into the snake's territory, which makes it feel trapped or cornered, or provocation of a snake during the breeding season. Even in these scenarios, only two snakes have a reputation as dangerous aggressors: the black mamba (Dendroaspis polylepis) of Africa and the king cobra (Ophiophagus hannah) of Southeast Asia. Nevertheless, snakes are inoffensive under the vast majority of circumstances. People are rarely indifferent about them, generally exhibiting emotions that range from religious awe and superstitious dread to repulsion and uncontrollable fear. It is interesting to note that, although most people profess to fear or hate snakes, one of the most visited areas of any zoo is the snake house—proof that snakes are mysterious and fascinating, even if they are loathed. Given their exquisite colours, patterns, and graceful movements as they crawl, swim, or climb, some snakes can be considered among the most beautiful animals.
Snakes can control the amount of venom they inject and may bite aggressively for food or defensively for protection. Snakes have a limited amount of venom available at any given time and do not want to waste it on nonprey organisms. As a result, about 40 percent of bites suffered by humans are defensive in nature and “dry” (without envenomation). Statistics show that the vast majority of snakebites occur while either catching and handling captive snakes or trying to molest or kill wild ones. In either case, the snake is only defending itself. rattlesnakes, for example, are venomous, and large ones are quite dangerous owing to the amount of venom they can inject. However, most are shy and retreating, and none will attack a person unmolested. When approached or molested, they will coil up and rattle as a warning to be left alone, striking only as a last resort. Most cases of reputed snake attack are based upon encroachment by a person into the snake's territory, which makes it feel trapped or cornered, or provocation of a snake during the breeding season. Even in these scenarios, only two snakes have a reputation as dangerous aggressors: the black mamba (Dendroaspis polylepis) of Africa and the king cobra (Ophiophagus hannah) of Southeast Asia. Nevertheless, snakes are inoffensive under the vast majority of circumstances. People are rarely indifferent about them, generally exhibiting emotions that range from religious awe and superstitious dread to repulsion and uncontrollable fear. It is interesting to note that, although most people profess to fear or hate snakes, one of the most visited areas of any zoo is the snake house—proof that snakes are mysterious and fascinating, even if they are loathed. Given their exquisite colours, patterns, and graceful movements as they crawl, swim, or climb, some snakes can be considered among the most beautiful animals.Nearly every culture since prehistoric times (including various present-day cultures) has worshipped, revered, or feared snakes. Serpent worship is one of the earliest forms of veneration, with some carvings dating to 10,000 BC. Although Satan is depicted as a serpent in the biblical account of the Creation, snakes are revered by most societies. Owing to ignorance, a vast global compendium of superstitions (superstition) and mythologies about snakes has sprung up. Many stem from the snake's biological peculiarities: its ability to shed the skin is associated with immortality; the ever-open eyes represent omniscience; their propensity for sudden appearance and disappearance allies snakes with magic and ghosts; a phallic resemblance embodies procreative powers; and the ability to kill with a single bite engenders fear of any snakelike creature.
 The hides of six snake species (especially pythons and wart snakes) are commonly bought and sold in the skin trade. The number of rattlesnakes used for their skins is minor in comparison. Hundreds of thousands of live snakes are collected for sale in the international pet trade. Nearly 100,000 ball pythons and 30,000 boa constrictors are imported annually into the United States. The removal of such enormous numbers from the wild threatens the survival of these species, and many snake populations are in decline owing to capture and habitat destruction.
The hides of six snake species (especially pythons and wart snakes) are commonly bought and sold in the skin trade. The number of rattlesnakes used for their skins is minor in comparison. Hundreds of thousands of live snakes are collected for sale in the international pet trade. Nearly 100,000 ball pythons and 30,000 boa constrictors are imported annually into the United States. The removal of such enormous numbers from the wild threatens the survival of these species, and many snake populations are in decline owing to capture and habitat destruction.Natural history
Most snakes do not spend much of their time doing anything but resting. A snake's primary activity is concerned with either thermoregulation or finding live food, which often involves passive waiting rather than active searching. The thermoregulation problem varies with latitude and altitude. The actions and reactions of a snake in temperate North America are distinct from those of one living in the American tropical lowlands but are similar to those of another living at higher altitudes in the Andes of Ecuador. No matter where they live, snakes are subjected to pressures from the living (biotic) parts of the environment as well as from the physical, nonliving (abiotic) parts. But the amount or degree of challenge to the snake from different segments of the environment changes drastically depending upon the region it inhabits. An individual living in the hot, humid tropics of Africa, with comparatively constant temperatures close to optimum throughout the year and ample moisture from both rainfall and the surroundings, faces environmental problems that are overwhelmingly biotic, involving competition with other members of its own species for food, the challenge from other species of snakes and perhaps other vertebrates for possession of the ecological niche, and constant pressure of the predators that find it a tasty morsel. On the other hand, the common adder, or European viper (Vipera berus), living north of the Arctic Circle in Europe, is the only snake present in the area and lives practically unchallenged in its niche. However, its survival is challenged continually by its physical environment, and death from overheating, freezing, or dehydration is a repetitive threat. These differences between animals from different parts of the world are reflected in their life histories, and it is neither possible nor legitimate to speak of the “life history of the snake” unless one speaks of only a single region or species.
Dormant periods
In the tropics, life continues at approximately the same activity level throughout the year. The only break in the rhythm comes in the dry season—and this only when the dry season is not just a period of slightly less rainfall. At such times, snakes may enter a short period of dormancy, which is at least in part a consequence of the effect that the dry season has upon their prey. This dormant period is similar to hibernation in winter by temperate-area snakes, although little is known about physiological changes that may or may not take place in tropical dormancy. At higher latitudes and altitudes, during periods of maximum stress (which for most snakes are the cold months), the animals must seek out a place where they can be completely inactive and nonreactive, where their inability to respond to the stimulus of danger is compensated for by the absence of danger, and where the surrounding extremes of low temperature and low humidity remain within tolerable limits. Such places are few and far between, and good hibernacula (dens used for hibernation) are recognized over generations and are utilized year after year, with snakes of several different species often sharing a den. It is likely that snakes, like sea turtles, can migrate by using celestial or geomagnetic cues. Scent trails, most commonly laid down by females during the breeding season, are also used. Many of the changes that occur in the individual snake after arriving at the hibernaculum are direct results of its dependence upon the environment. As the body cools, the heartbeat and respiration slow almost to a stop, and there is no muscular activity, little digestion, and no defecation. Physiological changes that are not correlated with or responsive to the surroundings also take place, but not to a degree comparable to those occurring in a hibernating mammal, and there is no “alarm system” to stir the snake into activity if a tolerance limit is passed. In such a case, the snake simply dies.
At the end of the cold season, the snake is totally dependent upon the changes in its surroundings to bring it back to activity; it cannot rouse itself. The stimuli are felt by all almost simultaneously, and snakes emerge by the dozens or even by the hundreds from some denning places. In some species, copulation takes place immediately after enough of the sun's rays have been absorbed to permit the development of an interest in the surroundings; in others, copulation (sexual intercourse) is the final act before entering hibernation, and the sperm remain dormant in the hibernating female. Fertilization of the egg can take place immediately after copulation, but, in at least some species, the female can store the sperm for several years, using them to fertilize successive batches of eggs.
Interactions between individuals
Snakes in both tropical and temperate regions tend to be solitary in their habits. The denning and mating aggregations are, for the most part, the only social events of the season. Sea snakes (sea snake) (subfamily Hydrophiinae) differ in this respect, sometimes being seen traveling in large troops, which seems to indicate an urge to aggregate. Female sea snakes also congregate in large numbers in seawall caves at parturition time, but this may have no social significance, since it seems to be a consequence of availability of a safe place for the young to be born rather than aggregational behaviour per se. There is some tendency for females of certain species in temperate areas to use a single site for egg deposition. Hunting of food is strictly an individual act for snakes; there are no known instances of cooperative hunting, as seen in some mammal and bird species. Hiding places and basking sites are occasionally shared; this again is a consequence of availability, and in the tropics, where hiding places abound, it is rare to find more than one snake at a time under a log or a rock. Except for these few weak instances, there is no development of social behaviour in snake populations—no establishment of social hierarchies, no territoriality, and perhaps no dominance. While combat dance (see below) certainly establishes a dominant individual temporarily, there is no indication that awareness of this dominance is retained by either snake. A dominance that must be reestablished at every encounter does not contribute to a social structure.
reproduction
Mating
The occurrence of mating immediately after emergence from hibernation allows snakes to take advantage of the fact that the females are accessible, concentrated, and receptive. The males are equally concentrated, so pair formation and copulation (sexual intercourse) are a simple matter. Males of some species have nuptial tubercles on various parts of the body, used to stroke or massage the female and, presumably, to arouse her sexually. Even when obvious tubercles are absent, the male uses a rubbing technique to stimulate the female, and in some species a muscle ripple moving along the male's body will provide a lateral caress. There are many descriptions in the literature of courtship dances done by snakes, in which the bodies are entwined and as much as one-third lifted off the ground, the coils ebbing and flowing with silent grace. Unfortunately, in many of these reports, the snakes were not captured and sexed, and the observer simply assumed that a male and female were involved. The combat dance engaged in by two males is believed to be a competitive behaviour for the acquisition of females during the breeding season. As in the courtship dance, the front of the bodies entwine and are raised higher and higher off the ground until finally one snake overthrows the other. It has been suggested that the combat dance is essentially a homosexual encounter, with each male attempting to copulate with the other. In any event, copulation is achieved after a comparatively brief courtship through the insertion of a hemipenis in the female's cloaca (a common urogenital chamber, lying just anterior to the anus). The hemipenis is one of a pair of mirror-image intromittent organs lying in the base of the male's tail, posterior to the anus, and strictly reserved for mating, for the urinary passages empty directly into the cloaca of the male. Either hemipenis can be used in copulation and must be everted through a process of turning itself inside out. This is achieved primarily by engorgement of the organ with blood.
The everted organ is heavily armed with spines, spinules (minute spines), flounces, calyxes, and other ornaments, all of which appear to play a role in ensuring that the male is securely attached to the female for the entire period until the sperm have been deposited. The sperm pass along a deep groove in the hemipenis, which, although open along one margin when examined in a dead snake, clearly forms a tubular passage as a result of the pressures of the engorged margins of the groove. After release, the sperm may immediately move up the oviducts and fertilize eggs just released from the ovary, or they may be stored by the female and released later to achieve fertilization.
egg formation and laying
Once fertilization has occurred, the egg begins to accumulate additional layers from the shell glands in the oviduct. In some species, this continues until a firm yet pliable leathery shell has been formed, permeable to both gases and liquids but capable of retaining much of its liquid content unless in a very dry place. The female then deposits the entire clutch of eggs in a protected damp, warm, and usually dark place, often along with clutches from other females of the same species, for the same stimuli that lead snakes to congregate for hibernation also take them to the same places for egg laying. Many species immediately abandon the eggs; some remain with the clutch and certainly appear to be protecting them from external danger; and a very few actually assume the role of a brood hen, maintaining a body temperature measurably higher than the surroundings and presumably assisting in incubation. In certain species, additional layers of membranous material are deposited around the embryo, but the calcareous (calcium-containing) shell does not form. Instead, the embryo is retained in the oviduct and continues its development there. This is termed ovoviviparous development (viviparity), since it is simply an egg retained in the oviduct, in contrast to viviparous, the condition seen in mammals, where the fetus develops in the uterus and establishes a placental connection with the uterine wall to permit exchange of materials with the maternal circulation. But, while an umbilical connection does not develop, there is considerable evidence of an exchange of materials between mother and fetus across their contiguous, highly vascularized membranous surfaces.
Regardless of the devices used to provide it with protection, the snake fetus is always brought to term before the onslaught of environmental conditions that could result in its death. The embryonic turtle can sleep away its first winter in the egg and hatch the following spring none the worse for the experience, but there is as yet no evidence that snakes can do the same. The contrast may result from the fact that the female turtle can scoop out a hole deep enough for freezing temperatures not to affect her brood, but the female snake is restricted, both by her limblessness and by the nature of the egg itself, to egg laying on or near the surface, where below-freezing temperatures are unavoidable. In the tropics, evidence is scanty, but it would appear that there is an endogenous (i.e., controlled from within) rhythm there as well, since young are not produced throughout the year.
Early development and growth
The young snake, whether from an egg or born alive, comes equipped with a sharp cutting device on its upper lip, the egg tooth. It slashes its way out of the rubbery eggshell with this tooth or, in the case of the live-born, cuts its way out of the soft membranes and is instantly competent to cope with its surroundings. Almost invariably, the first act of a newborn snake is to extend its tongue and taste the surroundings, conveying to the Jacobson's organ (a sensitive region in the roof of the mouth) chemical information perhaps more significant than the visual cues picked up by the pair of very inexperienced eyes. Young snakes begin to feed immediately after hatching, displaying considerable ability in the capture and consumption of prey. Venomous (venom) snakes are born with functional venom glands and fangs and are capable of immediate utilization of their most formidable weapons. Some of the viperid snakes are born with a bright green tail tip (contrasting strongly with the rest of the body colour), which they are capable of waving and shaking in a way that attracts the attention of possible prey. Within a very short time after birth, the first sloughing of the skin takes place, and the egg tooth is shed at about the same time.
The rate of growth is correlated with availability of food and temperatures high enough to permit full metabolic activity. When all factors are optimal, snakes grow surprisingly fast. A brood of California rosy boas (Charina trivirgata) doubled their length in a nine-month period, growing to only a few inches shorter than their mother, an adult close to maximum length for the species. It has been suggested that all snakes grow rapidly until they reach sexual maturity, after which time growth slows but very seldom stops completely. Snakes have indeterminate growth, which means there is no terminal point in time or size for growth in their lifetime, but they can continue to increase in length until they die. Sexual maturity is reached in about two years by many snakes. In the larger species, sexual maturity comes later, after four or five years or more.
In terms of total length, the reticulated pythons (Python reticulatus)—which inhabit Southeast Asia, Indonesia, and the Philippines—are the largest, growing to more than 7 metres (23 feet) as adults. In contrast, adults of the species Leptotyphlos carlae, a type of worm snake found on the island of Barbados, average only 10 cm (4 inches) long.
molt
A regularly recurrent event during the activity period of all snakes is the shedding, or molting, of the skin. Dormant individuals do not shed, but quite often this is one of the first events to take place after the end of dormancy. The integument of all animals represents the primary buffer between internal structures and the environment, and it is constantly subject to wear, tear, and other damage. The first line of defense against damage, especially when the skin is completely broken, is the formation of a blood clot or a scab, cellular reorganization, and scar formation. The second line of defense is the constant production of new cells in the deeper layers of the skin to replace cells lost or worn away from the surface. In snakes, the replacement procedure has been modified to a considerable degree. The replacement cells are not constantly produced independently of one another but grow on the same cycle and cohere into a complete unit. When this unit is functional, the old skin lying external to it becomes a threat to continued good health. At this point, the snake's eyes become a milky blue, an indication of a physiological loosening of the skin that forms the eye cap. This loosening is duplicated all over the body, although not so obviously. Shortly, the eyes clear, and the snake rubs loose the skin around the mouth and nose and crawls out of it completely, leaving a new, functional skin resplendent in fresh, bright colours. The rattlesnake sheds its skin in the same fashion as all other snakes, but the process is highly modified at the tail tip, where successive layers of keratinized, hardened epidermis are interlocked, or nested, to form the rattle, a device used to ward off large mammals. The use of the rattle is reasonably successful with buffalo, cattle, or horses but spectacularly unsuccessful with man because, through this advertising, the rattling snake precipitates either its death or its capture but seldom its escape.
locomotion
The snake has overcome the handicap of absence of limbs by developing several different methods of locomotion, some of which are seen in other limbless animals, others being unique. The first method, called serpentine locomotion, is shared with almost all legless animals, such as some lizards, the caecilians, earthworms, and others. This is the way most snakes move and has been seen by any zoo visitor. The body assumes a series of S-shaped horizontal loops, and each loop pushes against any resistance it can find in the environment, such as rocks, branches, twigs, dust, sand, or pebbles. The environment almost always provides sufficient resistance to make movement possible, and many snake species never use any other method of locomotion. Such species, when placed on a surface providing no resistance, such as smooth glass, are unable to move, whipping and thrashing around without progress. Snakes, like fishes and eels, swim by lateral undulation, which is essentially identical to serpentine locomotion. The sea snakes, however, possess a distinct anatomy in the form of a flattened, oarlike tail.
 Other methods of terrestrial movement also involve at least some resistance by the environment but usually less than the first. One of these is known as “concertina” locomotion, because the snake in action resembles the opening and closing of an accordion or a concertina. First the tail and the posteriormost part of the body are securely anchored, and then the head and the rest of the body are extended as far forward as possible from that secure base. At the maximum extension, the head and the forepart of the body are anchored and the posterior part drawn up as close as possible in the accordion-like folds. A second cycle follows the first, and the snake progresses. Tree snakes (tree snake), such as Imantodes and Oxybelis, modify this technique to move from branch to branch and have a strongly compressed body that permits surprising lengths of it to be stiffened and extended, using a modified I-beam effect for rigidity. The third method is called “caterpillar” or “rectilinear” locomotion, because the body moves in a straight line, using a flow of muscle contractions along the sides that looks like a caterpillar in motion. The body musculature is used for sequential lifting, anchoring, and pushing against individual ventral scales, which results in an inching along. It is used by large, heavy-bodied snakes, such as the boas and some of the vipers.
Other methods of terrestrial movement also involve at least some resistance by the environment but usually less than the first. One of these is known as “concertina” locomotion, because the snake in action resembles the opening and closing of an accordion or a concertina. First the tail and the posteriormost part of the body are securely anchored, and then the head and the rest of the body are extended as far forward as possible from that secure base. At the maximum extension, the head and the forepart of the body are anchored and the posterior part drawn up as close as possible in the accordion-like folds. A second cycle follows the first, and the snake progresses. Tree snakes (tree snake), such as Imantodes and Oxybelis, modify this technique to move from branch to branch and have a strongly compressed body that permits surprising lengths of it to be stiffened and extended, using a modified I-beam effect for rigidity. The third method is called “caterpillar” or “rectilinear” locomotion, because the body moves in a straight line, using a flow of muscle contractions along the sides that looks like a caterpillar in motion. The body musculature is used for sequential lifting, anchoring, and pushing against individual ventral scales, which results in an inching along. It is used by large, heavy-bodied snakes, such as the boas and some of the vipers.The fourth method is the least dependent on friction with the surface and is called sidewinding. This mode characterizes snakes living in the desert (though some non-desert dwellers also use it), where the sand simply gives way under any kind of push. The sidewinder does not progress forward when in motion but actually goes sideways. The snake, lying extended on the sand, lifts the anterior part of the body, moves it several centimetres to the side, and rests that part on the sand, maintaining the rest of the body as a lifted loop. This loop is then progressively shifted along the body to the end of the tail, at which time the entire snake has moved to the side from its previous position. By the time the first loop has reached the end of the tail, a new loop has already lifted the head and started down the body, and the snake looks like a coiled spring rolling across the sand. There is no rolling involved, however, since the ventral surface is always in contact with a substrate and usually leaves its impression behind in the sand, like a footprint. Sidewinding allows minimal contact of the body with hot sand and thereby allows the snake to be active at a time when surface travel would be lethal.
Form and function
 The most characteristic aspect of the snake form is the elongate body and tail and the absence of limbs. There is no snake in which the limb remnants still retain a function in locomotion, but complete or reduced elements of the pelvis and femur remain in many snake families, including the boa and python families. The body is usually slender, although there are some comparatively short and thick-bodied species. The body shape is correlated with activity level, with the slender species moving about all the time and the heavy forms leading a sedentary life. The pit vipers, for example, while not always long, are often big. It seems likely that these snakes evolved in the direction of heaviness only after the development of a heat-sensitive depression, the loreal pit, located between the eye and the nostril, and the venom apparatus, which enabled them to stay in one place and wait for their prey, rather than engaging in a continuous active search for food. Similarly, some of the largest nonvenomous snakes (boas, anacondas, and pythons) have labial pits that function in the same way as the loreal pit of the vipers, so they too can be sedentary and grow fat. Arboreal snakes are the most elongated and slender of all, with the tail (the region posterior to the anus) approaching half the length of the entire body. The body is often strongly compressed laterally, which permits greater rigidity of the body frame while crawling from branch to branch. Burrowing (fossorial) snakes are seldom large, and the true burrowers, the Typhlopidae and Leptotyphlopidae, living all their lives like earthworms, are the tiniest snakes of all. The burrowers have almost no tail, although some of them retain a spiny tail tip, which probably serves the animal as an anchoring point when crawling through the soil. The tail of sea snakes is flattened to form an oar, used to scull through the water. Sea snakes are almost totally helpless on land, locomoting only with the utmost difficulty.
The most characteristic aspect of the snake form is the elongate body and tail and the absence of limbs. There is no snake in which the limb remnants still retain a function in locomotion, but complete or reduced elements of the pelvis and femur remain in many snake families, including the boa and python families. The body is usually slender, although there are some comparatively short and thick-bodied species. The body shape is correlated with activity level, with the slender species moving about all the time and the heavy forms leading a sedentary life. The pit vipers, for example, while not always long, are often big. It seems likely that these snakes evolved in the direction of heaviness only after the development of a heat-sensitive depression, the loreal pit, located between the eye and the nostril, and the venom apparatus, which enabled them to stay in one place and wait for their prey, rather than engaging in a continuous active search for food. Similarly, some of the largest nonvenomous snakes (boas, anacondas, and pythons) have labial pits that function in the same way as the loreal pit of the vipers, so they too can be sedentary and grow fat. Arboreal snakes are the most elongated and slender of all, with the tail (the region posterior to the anus) approaching half the length of the entire body. The body is often strongly compressed laterally, which permits greater rigidity of the body frame while crawling from branch to branch. Burrowing (fossorial) snakes are seldom large, and the true burrowers, the Typhlopidae and Leptotyphlopidae, living all their lives like earthworms, are the tiniest snakes of all. The burrowers have almost no tail, although some of them retain a spiny tail tip, which probably serves the animal as an anchoring point when crawling through the soil. The tail of sea snakes is flattened to form an oar, used to scull through the water. Sea snakes are almost totally helpless on land, locomoting only with the utmost difficulty.Vertebrae
The vertebral column of snakes is highly elongated and has more vertebrae than any other living animal—up to 600 in the Australian python (Morelia oenpelliensis). Since there are no limb girdles associated with the skeleton, there are no good delimiters of regions, but snakes are generally regarded as having only two kinds of vertebrae: body (precaudal) and tail (caudal). There are 100–450 vertebrae in the body and 10–205 vertebrae in the tail. A pair of ribs is associated with each body vertebra except for a few immediately behind the head. By definition, there are no ribs on the tail vertebrae. Each vertebra articulates with its neighbour at five different points: first, at the contact point between the main, central bodies of the bones (centra), which is a ball-and-socket joint; then at two projections (prezygapophyses and postzygapophyses) from the centra, with articulating surfaces that lie above and below; and finally the zygosphenes and zygantra, found almost exclusively in snakes, the zygosphene being a projecting shelf on the upper part of the vertebra and the zygantrum being a pocket into which the zygosphene fits and within which it can swivel. These five points permit lateral and vertical rotation while preventing almost entirely any twisting of the vertebral column, thus achieving both flexibility and rigidity. The vertebra may bear on its ventral surface a long posteriorly directed projection called a hypapophysis. The presence or absence of this structure on the vertebrae of the posterior third of the body has been of considerable importance in snake classification because large groups of species show this as a common characteristic. In the egg-eating snakes (subfamily Dasypeltinae), the hypapophyses of a series of vertebrae a short distance behind the head have developed anteriorly directed tips that have a distinct coating of an enamel-like substance. These serve as eggshell breakers, projecting through a gap in the dorsal intestinal wall, where they can rip into a swallowed egg when the snake constricts the muscles of the body. The crushed shell is regurgitated, and the contents of the egg pass on to the stomach. The vertebrae of the tail tip in the rattlesnake are highly modified to form a “shaker” for the hollow rattle segments.
The skin
 Snakes are covered with scales (scale), which are cornified folds in the epidermal layers of the skin. These scales are usually arranged in rows along the body, the numbers and arrangement of which are characteristic of the species. The scales may be large and shield-shaped, in which case the number of rows is low (from 10 to 30), or they may be very small, rounded, and occasionally with the centre raised, in which case the number of rows can be as high as 180. A single scale may be very smooth and shiny (as in the rainbow snakes), have a raised ridge (keel) along its centre, be heavily striated, or even have a raised spine in the centre, as in the Javanese wart snake. The scales in some species have sensory structures on the posterior margins called apical pits, and all scales have various micro-ornamentations, consisting of hairlike projections, holes, spinules (small spines), and other specializations visible only through an electron microscope. The scales on the ventral surface of the body are modified into broad plates in the majority of species and are used in locomotion. The ventral scales of sea snakes (Hydrophiinae), worm snakes, and blind snakes are small, as on the dorsal surface, and there are several other species, including boas and pythons, that have partly enlarged ventral scales.
Snakes are covered with scales (scale), which are cornified folds in the epidermal layers of the skin. These scales are usually arranged in rows along the body, the numbers and arrangement of which are characteristic of the species. The scales may be large and shield-shaped, in which case the number of rows is low (from 10 to 30), or they may be very small, rounded, and occasionally with the centre raised, in which case the number of rows can be as high as 180. A single scale may be very smooth and shiny (as in the rainbow snakes), have a raised ridge (keel) along its centre, be heavily striated, or even have a raised spine in the centre, as in the Javanese wart snake. The scales in some species have sensory structures on the posterior margins called apical pits, and all scales have various micro-ornamentations, consisting of hairlike projections, holes, spinules (small spines), and other specializations visible only through an electron microscope. The scales on the ventral surface of the body are modified into broad plates in the majority of species and are used in locomotion. The ventral scales of sea snakes (Hydrophiinae), worm snakes, and blind snakes are small, as on the dorsal surface, and there are several other species, including boas and pythons, that have partly enlarged ventral scales. coloration
 The colours and colour patterns seen in snakes are often bright and occasionally spectacular. Snake colours are produced in two ways, either by pigment deposited in the skin or by differential diffraction of light as a consequence of the physical properties of the skin itself. When seen on a unicolour or uniform background, most snakes are obvious, and their colour patterns seem bold and prominent. When the animals are placed in their natural habitat, however, the significance of the colour patterns becomes obvious. The many lines running at sharp angles to the elongated lines of the body, the triangles or rectangles of colour, the blotches, spots, bands, or lozenges—all become highly disruptive to the eye, and the snake disappears into its surroundings. Blotched or spotted snakes tend to be sedentary and heavy-bodied, while striped and the occasional unicolour snakes are usually active species. In both cases, the coloration is protective, since a coiled sedentary snake has its body outline completely obscured by the overlapping patterns, while the stripes on a crawling snake eliminate the sensation of motion until they suddenly narrow at the tip of the tail and the snake disappears.
The colours and colour patterns seen in snakes are often bright and occasionally spectacular. Snake colours are produced in two ways, either by pigment deposited in the skin or by differential diffraction of light as a consequence of the physical properties of the skin itself. When seen on a unicolour or uniform background, most snakes are obvious, and their colour patterns seem bold and prominent. When the animals are placed in their natural habitat, however, the significance of the colour patterns becomes obvious. The many lines running at sharp angles to the elongated lines of the body, the triangles or rectangles of colour, the blotches, spots, bands, or lozenges—all become highly disruptive to the eye, and the snake disappears into its surroundings. Blotched or spotted snakes tend to be sedentary and heavy-bodied, while striped and the occasional unicolour snakes are usually active species. In both cases, the coloration is protective, since a coiled sedentary snake has its body outline completely obscured by the overlapping patterns, while the stripes on a crawling snake eliminate the sensation of motion until they suddenly narrow at the tip of the tail and the snake disappears. Although in most snakes the colours are such that they help the animal to hide, there are some species that seem to be advertising their presence rather than trying to hide it. Their patterns are aposematic (aposematic mechanism), or warning, in nature, and they let a possible enemy or predator know that it runs some risk in an encounter with the snake. The warning is effective, of course, only if the intruder is knowledgeable concerning its significance and can take heed. This implies a teaching and learning sequence, with the dangerous snake as “teacher” and the predator as “student.” For this reason, it has been suggested that the bright colours of the highly (and often fatally) venomous coral snakes did not evolve as warnings of the snakes' own poison but as mimics of some other venomous species, less dangerous but still able to teach the predator the significance of the warning coloration. There is no evidence that avoidance of aposematic species is instinctive; on the contrary, naive predators readily attempt to take aposematic forms. A predator that dies in its first encounter with a dangerous species cannot act as a selective force favouring the coloration of that species. There are quite a few mildly poisonous rear-fanged snakes, brightly banded in red, black, and yellow (colours found in the coral snakes), that can make a predator suffer a sufficiently painful lesson that it will avoid contact with all similarly coloured snakes, including the fatally venomous coral snakes and the completely harmless milk snakes (Lampropeltis) and scarlet snake. (For a complete discussion of the evolution of mimicry, see mimicry: The evolution of mimicry (mimicry).)
Although in most snakes the colours are such that they help the animal to hide, there are some species that seem to be advertising their presence rather than trying to hide it. Their patterns are aposematic (aposematic mechanism), or warning, in nature, and they let a possible enemy or predator know that it runs some risk in an encounter with the snake. The warning is effective, of course, only if the intruder is knowledgeable concerning its significance and can take heed. This implies a teaching and learning sequence, with the dangerous snake as “teacher” and the predator as “student.” For this reason, it has been suggested that the bright colours of the highly (and often fatally) venomous coral snakes did not evolve as warnings of the snakes' own poison but as mimics of some other venomous species, less dangerous but still able to teach the predator the significance of the warning coloration. There is no evidence that avoidance of aposematic species is instinctive; on the contrary, naive predators readily attempt to take aposematic forms. A predator that dies in its first encounter with a dangerous species cannot act as a selective force favouring the coloration of that species. There are quite a few mildly poisonous rear-fanged snakes, brightly banded in red, black, and yellow (colours found in the coral snakes), that can make a predator suffer a sufficiently painful lesson that it will avoid contact with all similarly coloured snakes, including the fatally venomous coral snakes and the completely harmless milk snakes (Lampropeltis) and scarlet snake. (For a complete discussion of the evolution of mimicry, see mimicry: The evolution of mimicry (mimicry).) skull and sense organs
Snakes rely on several senses to inform them of their surroundings. The pits, found in the region between the nostril and the eye in the pit vipers (the viperid subfamily Crotalinae) and in the scales of the lip line in some boas and pythons, are sensitive to very slight changes in temperature. These snakes feed almost exclusively on animals, such as birds and mammals, that maintain a constant body temperature and can therefore be located by the snake through the reception of the heat of the warm body. The heat lost by even a small rodent is sufficient to alert a waiting viper and enable it to direct a fast strike at the animal as it passes by. Death follows rapidly, and the snake follows the dying animal at a leisurely pace, perhaps in full awareness that it will not go far. The boids use the same technique for detecting warm prey, but after striking they retain the grip, killing by constriction.
The eye (eye, human) of the snake is lidless and covered by a transparent cap of epidermis, which is shed with the rest of the skin at each molt. Animals active during the day usually have round pupils, while the nocturnal species have a vertical or slit pupil that opens up in the dark, as does that of a cat, but closes more effectively in bright light, protecting the sensitive, dark-adapted retina. The eye has been almost completely lost in the burrowing families, in which it is visible only as a black spot and may be covered with a scale or flesh. Arboreal snakes often have a bulging, laterally placed eye, which permits them to see activities directly below as well as above and around them. The structure of the eye in snakes indicates that their lizard ancestor was probably a burrower and that all aboveground activity by snakes is a secondary invasion from an ancestral life underground.
The reception of sound (sound reception) is entirely by bone conduction within the skull. The snake has no external ear, but it still retains a few vestiges of the internal ear, which are connected to other skull bones in such a way as to permit transmission of some earth-borne, and perhaps a few aerial, sound waves of low frequency.
 The skull of snakes is characterized by mobility. It is light, with a reduced number of bones, and there are hinge joints at several levels that permit slight rotation or movement of one segment upon another. The only compact unit is the central braincase, with all other skull bones little more than attachments held in place by ligaments and muscles. In most snakes, the six bones of the upper jaw (left and right of the maxilla, palatine, and pterygoid) can move forward and back as well as sideways. The flexibility of the skull and the jawbones (jaw) attached to it is a major compensation for the loss of limbs and allows a snake to swallow prey that is several times the diameter of its head.
The skull of snakes is characterized by mobility. It is light, with a reduced number of bones, and there are hinge joints at several levels that permit slight rotation or movement of one segment upon another. The only compact unit is the central braincase, with all other skull bones little more than attachments held in place by ligaments and muscles. In most snakes, the six bones of the upper jaw (left and right of the maxilla, palatine, and pterygoid) can move forward and back as well as sideways. The flexibility of the skull and the jawbones (jaw) attached to it is a major compensation for the loss of limbs and allows a snake to swallow prey that is several times the diameter of its head.To swallow prey, the skulls of snakes move in a variety of ways, ranging from a lizardlike “inertial-feeding” mode, whereby the upper and lower jaws are both engaged forward over the prey items, to a “walk feeding,” whereby each of the four jaw arches moves independently of the others. Inertial feeding is found in the primitive blind snakes and thread snakes (infraorder Scolecophidia). In shieldtail snakes and their relatives (superfamily Anilioidea), a solid skull and limited movement of the jawbones allow “snout-shifting,” where one entire side of the head is moved forward in relation to the other side, which clamps the prey. Boas and pythons (superfamily Booidea) are more advanced and utilize a “walk-feeding” method in which one jaw arch is disengaged and moved forward, while the three other arches hold fast. The “walk” progresses by moving the jaw arches in order: upper left, upper right, lower left, lower right, and so on.
In most snakes, the upper jaw is connected to the lower jaw by a joint that acts as a pivot point, and, when eating, all toothed bones on one side of the mouth move forward as a unit. In the slug- and snail-eating snakes (the colubrid subfamilies Dipsadinae and Pareatinae), the connection between the upper jaw and the quadrate is lost, and there are four independent units rather than two. This jaw modification permits them to hold their food with three jaws while the fourth is advanced. The maxillary bone (the main bone of the upper jaw) of most snakes is elongated, with many teeth, but, in the Viperidae (Old World vipers, New World rattlesnakes, and other pit vipers), only one functional fang remains, on a short, blunt, rotatable maxillary. The position usually occupied by the maxillary has been taken by the pterygoid bone. In the Elapidae (cobras and relatives), the maxillary bears a single fang in a fixed position, sometimes followed by a few smaller, solid teeth. In several different evolutionary lines of snakes, the posterior one or two teeth on the maxillary have enlarged and changed, usually in the direction of developing a groove or canal on the anterior edge to conduct a flow of venom. These are the rear-fanged snakes, usually (although not always) nonlethal to man.
Snake teeth are usually long, slightly recurved, and needle-sharp. This facilitates swallowing and prevents loss of food, because the only direction in which a food item, which may be alive when swallowed, can go to escape the teeth is down the throat. Modifications in food habits have often been accompanied by changes in tooth structure or the loss of teeth. The egg-eating snakes (egg-eating snake), for example, have only a few peglike teeth left. The burrowing blind snakes have very reduced dentition and often have lost the teeth of one jaw entirely.
urogenital system
The urogenital system in snakes is not very distinctive from that of other vertebrates. The testes and ovaries tend to be staggered as a consequence of the elongation of the body, with the right usually lying anterior to the left. Snakes do not have a urinary bladder, and kidney wastes are excreted in a solid state as uric acid. As mentioned in the section Mating, the male snake has two separate intromittent organs, the hemipenes. This structure is not homologous with the penis of mammals but seems to represent a completely different solution to the problem of internal fertilization. It is a saclike structure that must be turned inside out to be inserted in the cloaca of the female and can be removed only by turning it back inside, because to draw it out directly would damage the female considerably. The hemipenis is extremely variable in its overall appearance and structure; the cloaca of the female is often similarly constructed, which thus prevents cross-fertilization by males of related species.
Specializations for securing food
The most essential and time-consuming activity for a snake during nondormant periods, regardless of its habitat, is the pursuit, capture, and digestion of food. Most snakes consume one prey item and then rest before eating again. There are many morphological and behavioral modifications to be observed in snakes that facilitate food gathering. Some of these changes are widespread and found in practically every kind of snake. Jacobson's organ, for example, is located in the roof of the mouth and is capable of detecting minute quantities of various chemical substances when they are picked up externally on the delicate double-tipped tongue and thrust into the organ for analysis. It is so significant and useful that it has never been lost by any snake species. While important primarily in trailing and recognition of prey, the Jacobson's organ is also used in detection of enemies, in trailing other snakes of the same species, and in courtship.
Some snakes have specialized salivary glands (salivary gland) that elaborate a potent venom, along with either grooved or tubular teeth to permit internal injection of the venom. This device for rapid immobilization of prey has proved successful whenever it has occurred, and several different lines of evolution can be detected both in the type of injector tooth and in the chemical composition or mode of action of the venoms. The types of injector tooth include the long tubular fang of the viperids, the short hollow fang on a fixed maxillary seen in the elapids, and the several kinds of grooved tooth on the posterior end of the maxillary. In the latter case, there is little doubt but that the development of grooving has taken place several different times in different places. As for the venom, it is true that the terms neurotoxic, hemotoxic, and cardiotoxic, once used to describe the different effects, are clearly misleading and are too simple for accurate statements concerning venom composition. Nevertheless, it is still accurate to say that some components of venom cause changes in red blood cells, coagulation defects, and blood-vessel injury, while others produce deleterious changes in sensory and motor functions and in respiration, and still others have a direct effect on the heart. Some venom kills very rapidly, such as that of the golden fer-de-lance (Bothrops insularis) of Queimada Island, off the Brazilian coast, which would lose as prey most of the birds it bites if they could fly very far away. Other venoms kill more slowly, and the snake bites, retires, and waits, finally trailing the bitten prey, using the tongue and the Jacobson's organ until it finds the already stiffening body. Some snakes, in particular the rear-fanged species, bite, chew, and hold on, eventually bringing the hindmost maxillary teeth into play, which permits the injection of toxin.
Other feeding specializations are not so widespread among species, and some are restricted to a single group. Nearly all boids and many colubrids utilize a constriction method for killing their food. The prey is struck and held by the teeth, and a series of body coils are rapidly thrown around it. These coils tighten until respiration is impossible and suffocation results, but very seldom are bones crushed or broken. The snail-eating snakes have a series of modifications of the teeth, the toothed bones, and the lower jaw that permit insertion of the lower jaw into a shell to pull out the snail's body. One genus of sea snakes (sea snake), Microcephalophis, has a tiny head and a long neck with the same diameter as the head, which can be inserted deeply into very narrow holes inhabited by its prey. An Asian water snake, Erpeton tentaculatus, has a sizable pair of tentacles on its snout, the purpose of which is uncertain. There is a great correlation between the difficulty in catching a particular kind of prey and the development of morphological and behavioral devices to help solve the problem. The blind snake living in a termite nest needs no more than its tongue and Jacobson's organ to permit it to recognize the soft-bodied, defenseless termites, so its eyes and most of its teeth have been lost. But the rattlesnake, seeking the most elusive prey of all in agile, aware, and sensorially acute small mammals, has a full armament of equipment, including the facial pit, the venom gland, the rotatable maxillary bone, the tubular fang, and two functional eyes.
Evolution
The origin of snakes has been debated since the mid-19th century. On the basis of unique shared derived features (such as paired copulatory organs and the Jacobsen's organ), it is clear that snakes and lizards are close relatives. However, it is not known whether snakes evolved from (1) a prelizard group, (2) a common ancestor of both lizards and snakes, or (3) a lizard group. The last hypothesis is currently in favour, but the specific group (living or extinct) is uncertain. Concerning habitat type, it has been thought that snakes evolved from (1) an aquatic ancestor (such as mosasauroids (mosasaur)), (2) a burrowing ancestor (such as the amphisbaenian worm lizards or the blind lizards), or (3) a secretive ancestor. Each of these is consistent with the morphological changes observed in modern snakes as compared with lizards (loss of limbs, lengthening of body, absence of movable eyelids and external ear opening). The discovery of three marine snake fossils with hind limbs (Hassiophis, Pachyrhachis, Podophis) from the Cretaceous Period (144 million to 65 million years ago) of the Middle East has sparked renewed controversy over the origin of snakes. The interpretation and analysis of these fossils vary among researchers. Some classify them as the most primitive serpents in a sister group to all snakes, whereas others suggest they are a more advanced group related to boas and pythons. The latter interpretation, if correct, would mean either that the hind limbs were re-evolved in these snakes or that the snakes represent an independent evolutionary line that never lost the hind limbs. This debate may be resolved only with the discovery of additional fossils and further analysis.
 The evolutionary history of snakes can be characterized as the transition from the lizardlike “inertial-feeding” mode of the primitive Scolecophidia ( worm snakes and blind snakes), whereby the upper and lower jaws are both engaged forward over the prey items, to “walk feeding.” Although the fossil record of snakes is poor and fragmentary compared with other vertebrates, much is known about the evolution of living and extinct groups. The oldest fossils are found in South America, Africa, and Madagascar from the time when these continents were joined as Gondwanaland (Gondwana). The earliest undisputed snake (Lapparentophis) was terrestrial. Found in Africa, it dates to 120 million years ago, but it can be assumed that snakes originated farther back in time, probably during the Late Jurassic (Jurassic Period) Epoch (159 million to 144 million years ago). The initial dichotomy among living snakes occurred between the specialized, burrowing Scolecophidia (worm snakes and blind snakes) and the generalized Alethinophidia (ancient and modern snakes). Given present distribution, it is likely that the Scolecophidia originated in Gondwanaland; the fossil record clearly indicates a Gondwana origin for the superfamilies Anilioidea and Booidea. Some of the more advanced snakes (families Viperidae, Elapidae, and Colubridae) originated in the ancient continent of Laurasia, their earliest fossils being from North America and Europe.
The evolutionary history of snakes can be characterized as the transition from the lizardlike “inertial-feeding” mode of the primitive Scolecophidia ( worm snakes and blind snakes), whereby the upper and lower jaws are both engaged forward over the prey items, to “walk feeding.” Although the fossil record of snakes is poor and fragmentary compared with other vertebrates, much is known about the evolution of living and extinct groups. The oldest fossils are found in South America, Africa, and Madagascar from the time when these continents were joined as Gondwanaland (Gondwana). The earliest undisputed snake (Lapparentophis) was terrestrial. Found in Africa, it dates to 120 million years ago, but it can be assumed that snakes originated farther back in time, probably during the Late Jurassic (Jurassic Period) Epoch (159 million to 144 million years ago). The initial dichotomy among living snakes occurred between the specialized, burrowing Scolecophidia (worm snakes and blind snakes) and the generalized Alethinophidia (ancient and modern snakes). Given present distribution, it is likely that the Scolecophidia originated in Gondwanaland; the fossil record clearly indicates a Gondwana origin for the superfamilies Anilioidea and Booidea. Some of the more advanced snakes (families Viperidae, Elapidae, and Colubridae) originated in the ancient continent of Laurasia, their earliest fossils being from North America and Europe.The burrowing Anilioidea of South America and Southeast Asia is the most primitive group of alethinophidians. The Aniliidae of the Amazon probably most closely represent the ancestral snake condition. They are moderate-sized, with a lizardlike skull, barely enlarged ventral shields, and a vestigial left lung. Fossil anilioids are known from the Cretaceous of South America to the Eocene Epoch (54.8 million to 33.7 million years ago) of Europe. The anilioids gave rise to the booids, a primitive but more generalized group of boas and pythons. The fossil record of the extinct terrestrial Madtsoiidae extends from about 80 million years ago to the Pleistocene Epoch (1,800,000 to 10,000 years ago) of Europe, Africa, Madagascar, South America, and Australia. The two most primitive living booid families, Xenopeltidae and Loxocemidae, are small ground-dwelling snakes. The Boidae and Pythonidae are the first snakes to have moved aboveground and diversified into terrestrial, arboreal, and aquatic habitats. Their success was probably due to increased size and changes in the feeding apparatus (loosening of the skull's jaw bones, increased dentition on the four jaw arches, expandable skin between scales) that permitted them to subdue large warm-blooded mammals and birds by constriction. Boas and pythons are large, heavy, and slow-moving and include the largest living serpents, with some reaching 6 to 10 metres (20 to 33 feet) in length. During the Late Cretaceous, a land bridge connecting South America to North America allowed anilioid and booid snakes to invade the northern supercontinent of Laurasia from Gondwanaland. These two groups then spread from North America eastward to Europe and possibly westward into Asia. The fossil record indicates that the height of booid evolution and diversity occurred in the Eocene, when they dominated the world's snake fauna, after which time they diminished in numbers. Between the end of the Cretaceous and the middle of the Eocene, the largest terrestrial vertebrate in the world was a booid snake (Titanoboa cerrejonensis). Known from a single fossilized vertebra, Titanoboa probably weighed 1,135 kilograms (about 2,500 pounds) and reached a length of 13 metres (about 43 feet).
The most advanced snake group, the superfamily Colubroidea, evolved during the Eocene in Laurasia. These snakes were smaller and faster and possessed more flexible jaws than the booids, and several groups developed venom glands and fangs for subduing prey. The fossil record indicates that colubroids outcompeted the booids, which today number fewer than 80 species, as opposed to some 2,200 colubroids. The Oligocene Epoch (33.7 million to 23.8 million years ago) was a transitional period, with the booids and anilioids diminishing in numbers and the colubroids increasing. The Miocene Epoch (23.8 million to 5.3 million years ago) was a period of great expansion of colubroids in North America, Europe, and Asia. The first venomous snakes evolved in the Early Miocene, with the Viperidae appearing in North America, Europe, and Asia and the Elapidae in North America and Europe. It was during the Miocene that Africa was invaded by the Colubridae, Elapidae, and Viperidae from Europe. The diversification and expansion of the Colubridae as the dominant snake group began in the Miocene and has continued to the present. During Australia's long separation and isolation from Gondwanaland and other continents, only four families existed there, three of which survive today (Typhlopidae, Pythonidae, and Elapidae) and one of which became extinct in the Pleistocene (Madtsoiidae).
Annotated classification
The classification presented here is a synthesis of three separate works by G. Underwood, H.G. Dowling, and S.B. McDowell.
Suborder Serpentes (snakes (snake))
2,900 species in 2 infraorders of 460 genera. In addition to the living families listed below, 10 extinct snake families are known: (1) the marine Pachyophiidae of Early to Late Cretaceous of Europe and Asia, (2) the terrestrial Lapparentophiidae from the Early Cretaceous of Africa and Europe, (3) the marine Simoliophiidae from the Late Cretaceous of Europe, Asia, and Africa, (4) the terrestrial Madtsoiidae of the Late Cretaceous to Pleistocene of Europe, Africa, Madagascar, South America, and Australia, (5) the terrestrial Dinilysiidae of the Late Cretaceous of South America, (6) the semiaquatic and marine Palaeophiidae of the Late Cretaceous to Eocene of Europe and Asia, (7) the aquatic Nigerophiidae of the Late Cretaceous to Eocene of Europe, Asia, and Africa, (8) the aquatic Vialovophiidae of the Paleocene to Eocene of North America and Asia, (9) the aquatic Anomalophiidae of the Eocene of Europe, and (10) the aquatic Russellophiidae of the Late Cretaceous to Eocene of Europe and Africa.
Infraorder Scolecophidia
375 species in 3 families of 12 genera. Small wormlike snakes with a short blunt tail that resembles the head. Pelvic vestiges usually present. Mouth small. Eyes reduced or invisible; when present, beneath a large head shield. Retina with only single rods. Left lung absent. One pair of thymus glands. Liver segmented. Left oviduct usually absent. Rectal cecum present. Cutaneous sebaceous glands. Fossorial with a specialized diet of eggs, pupae, larvae, and adult ants and termites, which are ingested in large quantities. Lays 1–60 eggs. Fossils from the Paleocene to Pleistocene of North America, Europe, and Africa.
Superfamily Typhlopoidea (blind snake)
Family Typhlopidae ( blind snakes)
255 species in 6 genera. Size small, 15–50 cm, but a few species reach 75–90 cm. Pelvic vestiges present. Tracheal lung present. 1 species (Ramphotyphlops braminus) parthenogenetic (parthenogenesis).
Family Anomalepididae (dawn blind snakes)
15 species in 4 genera from Central America to northern South America. Size small, 15–40 cm. Pelvic vestiges absent. Tracheal lung present. Possible fossil from the Paleocene of Texas.
Superfamily Leptotyphlopoidea
Family Leptotyphlopidae ( worm snakes or thread snakes)
105 species in 2 genera of the southwestern United States to South America, Africa, and southwestern Asia. Size very small, 7–35 cm. Pelvic vestiges present.
Infraorder Alethinophidia (typical, or advanced, snakes)
2,500 species in 16 families of about 450 genera. Size small to very large. Mouth large. Eye covered with a clear spectacle or brille, conforming to shape of eye. Sebaceous glands absent. Two pairs of thymus glands. Liver unsegmented. Rectal cecum absent. Midventral scales transversely enlarged, usually at least as wide as body except in some primitive groups. The first five superfamilies listed below are more primitive and thus referred to as “lower snakes,” Colubroidea being “higher snakes.”
Superfamily Anilioidea
60 species in 12 genera. Small tropical fossorial or secretive snakes. Four fossil genera with 7 species from the Cretaceous to Pliocene of North and South America and Europe.
Family Uropeltidae (shieldtail snakes (shieldtail snake))
45 species in 9 genera from southern India and Sri Lanka. Size small, 25–50 cm. Pelvic vestiges absent. Left lung absent. Bears living young.
Family Cylindrophiidae (pipe snakes)
8 species in 1 genus. Size moderate, 0.4–1 metre. Pelvic vestiges present. Left lung present. Bears living young.
Family Anomochilidae (stump heads)
2 species in 1 genus. 1 species from peninsular Malaysia, the other from Borneo. Size small, 35 cm. Pelvic vestiges present. Left lung and rectal cecum absent. Lays eggs.
Family Aniliidae (coral pipe snake)
1 species (Anilius scytale) from the Amazon basin. Size moderate, 0.7–1 metre. Pelvic vestige present. Small eye beneath a head shield. Bears living young.
Superfamily Booidea
Approximately 80 species in 20 genera found worldwide. Left lung large, 50–75 percent of the right lung length. Approximately 50 fossil genera with more than 70 species from the Cretaceous to Pleistocene.
Family Boidae ( boas)
Approximately 45 species in 9 genera from the New World tropics, Madagascar, and the southwestern Pacific. Size moderate to very large, 1–8 metres. Pelvic vestiges present. Bears living young.
Family Pythonidae ( pythons)
Approximately 30 species in 8 genera of the Old World tropics. Size large to very large, 2–10 metres. Pelvic vestiges present. Lays eggs.
Family Xenopeltidae ( sunbeam snakes)
2 species in 1 genus from Southeast Asia and Indonesia. Size moderate, 0.8–1.1 metres. Semifossorial. Pelvic vestiges absent. Teeth hinged. Lays eggs.
Family Loxocemidae (New World python)
1 species (Loxocemus bicolor) from Mexico to Costa Rica. Size moderate, 0.8–1.4 metres. Fossorial. Pelvic vestiges present. Lays eggs.
Superfamily Tropidophioidea
Family Tropidophiidae (dwarf boas or wood snakes)
35 species in 4 genera from Mexico to Ecuador and the West Indies. Size small, 30–60 cm. Terrestrial. Pelvic vestiges present. Tracheal lung present, left lung absent. Bears living young. Four fossil genera with 35 species from the Eocene to Oligocene of North America and Europe.
Family Xenophidiidae (strange snakes)
2 species in 1 genus of Malaysia. Size small, 30 cm. Terrestrial. Pelvic vestiges absent. Hinged teeth. Lays eggs.
Superfamily Bolyerioidea
Family Bolyeriidae (split-jawed boas)
1 remaining species, 1 recently extinct, both from Mauritius. Maxilla divided into two elements, a condition unique among vertebrates. Pelvic vestiges absent. Lays eggs.
Superfamily Acrochordoidea
Family Acrochordidae (wart snake) (wart snakes)
3 species in 1 genus from Southeast Asia and Australia. Size moderate to large, 1–2.7 metres. Marine. Left lung absent, tracheal lung present. Bears living young. One fossil species from the Miocene of Pakistan.
Superfamily Colubroidea (“higher snakes”)
More than 2,300 species in 410 genera. Some groups have a tubular fang at the anterior end or grooved fangs at the posterior end of the upper jaw. Right common carotid artery absent or reduced.
Family Colubridae (colubrid) (“harmless” and rear-fanged snakes)
1,700 species in approximately 300 genera found worldwide except for most of Australia. Size small to very large, 0.3–3 metres. Includes the majority of harmless snakes in the world. Seventeen fossil genera from the Eocene to Pliocene of North and South America, Europe, and Africa.
Subfamily Colubrinae ( king snakes, racers, bull snakes, and rat snakes)
700 species in 100 genera from North America, Eurasia, and the East Indies. Size small to very large, 0.5–3 metres. Majority are constrictors feeding on mammals and birds. Lays eggs.
Subfamily Xenodontinae ( hognose snakes, ring-necked snakes, and rainbow snakes)
600 species in 85 genera found in Mexico, Central and South America, and the West Indies. Size small to large, 0.5–2 metres. Grooved rear fangs. Lays eggs.
Subfamily Natricinae ( garter snakes and water snakes)
250 species in 35 genera from North America, Eurasia, and the East Indies. Size small, 0.2–2 metres. Semiaquatic or aquatic. Most feed on amphibians and fish. Lays eggs and bears living young.
Subfamily Lamprophiinae ( house snakes)
220 species in 45 genera of Africa and Madagascar. Size small to large, 0.4–2.6 metres. Lays eggs.
Subfamilies Pareatinae and Dipsadinae (slug- and snail-eating snakes)
70 species in 6 genera. Size small, 30–90 cm. Semiarboreal, specialized for eating snails and slugs. Lays eggs. Dipsadinae of Mexico, Central America, and South America with 3 genera and 50 species; Pareatinae of southern Asia and the East Indies with 3 genera and 20 species.
Subfamilies Xenodermatinae and Nothopsinae
40 species in 11 genera. Size small, 30–80 cm. Terrestrial and semiaquatic. Vertebrae with dorsal and lateral expansions. Tracheal lung present, no left lung. Bears living young. Xenodermatinae of Southeast Asia with 4 genera with 20 species; Nothopsinae of Central and South America with 7 genera and 20 species.
Subfamily Homalopsinae (rear-fanged water snakes)
35 species in 10 genera from Pakistan and China to Australia. Size small, 0.5–1 metre. Aquatic (marine, brackish and fresh water). Fish- and frog-eating except for one genus (Fordonia) which feeds upon crabs. Grooved rear fangs. Valvular nostrils. Tracheal lung present. Bears living young.
Subfamily Dasypeltinae (egg-eating snakes)
7 species in 2 genera, 6 African, 1 Asian. Size moderate, 0.5–1 metre. Semiarboreal with specialized diet of bird eggs. Dentition reduced to several small teeth. Anterior vertebral projections that crush eggshells when swallowed. Lays eggs.
Family Elapidae (elapid) ( cobras, mambas, coral snakes, taipans, kraits, and sea snakes)
315 species in 60 genera found worldwide except Europe, Madagascar, and Antarctica. Size small to very large, 0.7–6 metres. Maxilla with a short, tubular, venom-conducting fang. Fossils from the Miocene of North America, Europe, and Africa.
Subfamily Elapinae
150 species in 20 genera found in the New World, Africa, Asia, and Southeast Asia. Eye with round pupil. Lays eggs.
Subfamily Oxyuraninae.
110 species in 28 genera from Australia and New Guinea. Smooth scales. Eye with round pupil. Terrestrial and fossorial. Lays eggs.
Subfamily Hydrophiinae (sea snakes)
55 species in 16 genera of the tropical western Pacific and Indian oceans; 1 species (Pelamis platurus) ranging to the Pacific coast of the New World. Size moderate, 1–2 metres. Marine with tail and part of the body compressed vertically for swimming. Tracheal lung present, left lung absent. Bears living young.
Subfamily Laticaudinae (sea kraits)
6 species in 1 genus of Southeast Asia and southwest Pacific. Tail compressed vertically for swimming.
Family Viperidae ( vipers and adders)
260 species in 35 genera. Movable maxillary bones, each with a single long tubular venom-injecting fang that is folded against roof of mouth when closed. Tracheal lung present. Eye with vertical pupil. Keeled scales. Fossils from the Miocene to Pleistocene of North America, Europe, and Africa.
Subfamily Viperinae (true vipers)
90 species in 15 genera of the Old World except Madagascar, New Guinea, and Australia. Size moderate to large, 0.6–1.5 metres. Terrestrial. Lacking loreal pit. Bears living young.
Subfamily Crotalinae (pit vipers)
170 species in 20 genera of the New World and eastern Asia. Size moderate to large, 1–3.6 metres. Heat-sensing loreal pit between nostril and eye. Bears living young except for a few Asian egg-laying species.
Family Atractaspididae (stiletto vipers, burrowing asps, and aparallactines)
65 species in 12 genera of Africa and adjacent southwestern Asia. Size small, 0.3–1.1 metres. Fossorial or secretive. Maxilla with grooved fangs on either anterior or posterior end of maxilla. Venomous, though only one genus (Atractaspis) is dangerous to humans. Smooth scales. Eye with round pupil. Lays eggs.
Additional Reading
General works
Harry W. Greene, Snakes: The Evolution of Mystery in Nature (1997), surveys snake diversity and biology and is complemented by stunning photography. John Coborn, The Atlas of Snakes of the World (1991), discusses the care of snakes and snakebites in addition to describing each family and genus. Carl H. Ernst and George R. Zug, Snakes in Question: The Smithsonian Answer Book (1996), provides answers to many of the questions most frequently asked about snakes. Tony Phelps, Poisonous Snakes, rev. ed (1989), covers the rear-fanged colubrids, the elapids, and the vipers. Ramona and Desmond Morris, Men and Snakes (1965), thoroughly reviews man's relationships with snakes, including culture, mythology, medicine, and exploitation. The Ultimate Guide: Snakes (1997), a video produced for the Discovery Channel by Partridge Films, portrays snakes in the wild and also includes computer graphics and interviews with researchers.
Advanced works
Roy W. McDiarmid, Jonathan A. Campbell, and T'Shaka A. Touré, Snake Species of the World: A Taxonomic and Geographic Reference, vol. 1 (1999– ), constitutes the first of a comprehensive series. Richard A. Seigel, Joseph T. Collins, and Susan S. Novak, Snakes: Ecology and Evolutionary Biology (1987), is suitable for amateur naturalists as well as researchers. F. Harvey Pough (ed.), Herpetology, 2nd ed. (2001), is a college-level textbook.
- U.S. Stock Market Prices, Table
- U.S. Stock Market Prices, Table 1
- U.S. Supreme Court justices
- Ussuri River
- Ussuriysk
- Ustaša
- Ust-Ilimsk
- Ustinov, Dmitry Fedorovich
- Ustinov, Sir Peter
- Ust-Kamenogorsk
- Ust-Ordyn Buryat
- Ust-Ordynsky
- U.S. Trade Balance with Major Trading Partners
- Ustyurt Plateau
- usufruct
- Usuki
- Usulután
- Usumacinta River
- usury
- USX Corporation
- Uta
- Utagawa Kunisada
- Utagawa Kuniyoshi
- Utagawa Toyokuni
- Utah