tropical rainforest
Introduction
also spelled tropical rain forest
luxuriant forest, generally composed of broad-leaved trees and found in wet tropical uplands and lowlands around the Equator.
 Rainforests are vegetation types dominated by broad-leaved trees (tree) that form a dense upper canopy (layer of foliage) and contain a diverse array of vegetation. Contrary to common thinking, not all rainforests occur in places with high, constant rainfall; for example, in the so-called “dry rainforests” of northeastern Australia the climate is punctuated by a dry season, which reduces the annual precipitation. Nor are all forests in areas that receive large amounts of rainfall true rainforests; the conifer-dominated forests in the extremely wet coastal areas of the American Pacific Northwest are temperate evergreen forest ecosystems. Therefore, to avoid conveying misleading climatic information, the term rainforest is now preferred over rain forest.
Rainforests are vegetation types dominated by broad-leaved trees (tree) that form a dense upper canopy (layer of foliage) and contain a diverse array of vegetation. Contrary to common thinking, not all rainforests occur in places with high, constant rainfall; for example, in the so-called “dry rainforests” of northeastern Australia the climate is punctuated by a dry season, which reduces the annual precipitation. Nor are all forests in areas that receive large amounts of rainfall true rainforests; the conifer-dominated forests in the extremely wet coastal areas of the American Pacific Northwest are temperate evergreen forest ecosystems. Therefore, to avoid conveying misleading climatic information, the term rainforest is now preferred over rain forest.This section covers only the richest of rainforests—the tropical rainforests of the ever-wet tropics.
Origin
Tropical rainforests represent the oldest major vegetation type still present on the terrestrial Earth. Like all vegetation, however, that of the rainforest continues to evolve and change, so that modern tropical rainforests are not identical with rainforests of the geologic past.
 Tropical rainforests grow mainly in three regions: the Malesian botanical subkingdom, which extends from Myanmar (Burma) to Fiji and includes the whole of Thailand, Malaysia, Indonesia, the Philippines, Papua New Guinea, the Solomon Islands, and Vanuatu and parts of Indochina and tropical Australia; tropical South (South America) and Central America, especially the Amazon basin; and West and Central Africa (Western Africa) (see biogeographic region). Smaller areas of tropical rainforest occur elsewhere in the tropics wherever climate is suitable. The principal areas of tropical deciduous forest (monsoon forest) are in India, the Myanmar–Vietnam–southern coastal China region, and eastern Brazil, with smaller areas in South and Central America north of the Equator, the West Indies, southeastern Africa, and northern Australia.
Tropical rainforests grow mainly in three regions: the Malesian botanical subkingdom, which extends from Myanmar (Burma) to Fiji and includes the whole of Thailand, Malaysia, Indonesia, the Philippines, Papua New Guinea, the Solomon Islands, and Vanuatu and parts of Indochina and tropical Australia; tropical South (South America) and Central America, especially the Amazon basin; and West and Central Africa (Western Africa) (see biogeographic region). Smaller areas of tropical rainforest occur elsewhere in the tropics wherever climate is suitable. The principal areas of tropical deciduous forest (monsoon forest) are in India, the Myanmar–Vietnam–southern coastal China region, and eastern Brazil, with smaller areas in South and Central America north of the Equator, the West Indies, southeastern Africa, and northern Australia.The flowering plants (angiosperms (angiosperm)) first evolved and diversified during the Cretaceous Period about 100 million years ago, during which time global climatic conditions were warmer and wetter than those of the present. The vegetation types that evolved were the first tropical rainforests, which blanketed most of the Earth's land surfaces at that time. Only later—during the middle of the Cenozoic Era, about 40 million years ago—did cooler, drier climates develop, leading to the development across large areas of other vegetation types.
It is no surprise, therefore, to find the greatest diversity of flowering plants today in the tropical rainforests where they first evolved. Of particular interest is the fact that the majority of flowering plants displaying the most primitive characteristics are found in rainforests (especially tropical rainforests) in parts of the Southern Hemisphere, particularly South America, northern Australia and adjacent regions of Southeast Asia, and some larger South Pacific Islands. Of the 13 angiosperm families generally recognized as the most primitive, all but two— Magnoliaceae and Winteraceae—are overwhelmingly tropical in their present distribution. Three families—Illiciaceae, Magnoliaceae, and Schisandraceae—are found predominantly in Northern Hemisphere rainforests. Five families—Amborellaceae, Austrobaileyaceae, Degeneriaceae, Eupomatiaceae, and Himantandraceae—are restricted to rainforests in the tropical Australasian region. Members of the Winteraceae are shared between this latter region and South America, those of the Lactoridaceae grow only on the southeast Pacific islands of Juan Fernández, members of the Canellaceae are shared between South America and Africa, and two families— Annonaceae and Myristicaceae—generally occur in tropical regions. This has led some authorities to suggest that the original cradle of angiosperm evolution might lie in Gondwanaland (Gondwana), a supercontinent of the Southern Hemisphere thought to have existed in the Mesozoic Era (248 million to 65 million years ago) that consisted of Africa, South America, Australia, peninsular India, and Antarctica. An alternative explanation for this geographic pattern is that in the Southern Hemisphere, especially on islands, there are more refugia—i.e., isolated areas whose climates remained unaltered while those of the surrounding areas changed, enabling archaic life-forms to persist.
The first angiosperms are thought to have been massive, woody plants appropriate for a rainforest habitat. Most of the smaller, more delicate plants that are so widespread in the world today evolved later, ultimately from tropical rainforest ancestors. While it is possible that even earlier forms existed that await discovery, the oldest angiosperm fossils—leaves, wood, fruits, and flowers derived from trees—support the view that the earliest angiosperms were rainforest trees. Further evidence comes from the growth forms of the most primitive surviving angiosperms: all 13 of the most primitive angiosperm families consist of woody plants, most of which are large trees.
As the world climate cooled in the middle of the Cenozoic, it also became drier. This is because cooler temperatures led to a reduction in the rate of evaporation of water from, in particular, the surface of the oceans, which led in turn to less cloud formation and less precipitation. The entire hydrologic cycle slowed, and tropical rainforests—which depend on both warmth and consistently high rainfall—became increasingly restricted to equatorial latitudes. Within those regions rainforests were limited further to coastal and hilly areas where abundant rain still fell at all seasons. In the middle latitudes of both hemispheres, belts of atmospheric high pressure developed. Within these belts, especially in continental interiors, deserts formed (see desert: Origin (desert)). In regions lying between the wet tropics and the deserts, climatic zones developed in which rainfall adequate for luxuriant plant growth was experienced for only a part of the year. In these areas new plant forms evolved from tropical rainforest ancestors to cope with seasonally dry weather, forming tropical deciduous forests. In the drier and more fire-prone places, savannas (savanna) and tropical grasslands (grassland) developed.
Retreat of the rainforests was particularly rapid during the period beginning 5,000,000 (Cenozoic Era) years ago leading up to and including the Pleistocene (Pleistocene Epoch) Ice Ages, or glacial intervals, that occurred between 1,600,000 and 10,000 years ago. Climates fluctuated throughout this time, forcing vegetation in all parts of the world to repeatedly migrate, by seed dispersal, to reach areas of suitable climate. Not all plants were able to do this equally well because some had less-effective means of seed dispersal than others. Many extinctions resulted. During the most extreme periods (the glacial maxima, when climates were at their coldest and, in most places, also driest), the range of tropical rainforests shrank to its smallest extent, becoming restricted to relatively small refugia. Alternating intervals of climatic amelioration led to repeated range expansion, most recently from the close of the last glacial period about 10,000 years ago. Today large areas of tropical rainforest, such as Amazonia, have developed as a result of this relatively recent expansion. Within them it is possible to recognize “hot spots” of plant and animal diversity that have been interpreted as glacial refugia.
Tropical rainforests today represent a treasure trove of biological heritage. They not only retain many primitive plant and animal species but also are communities that exhibit unparalleled biodiversity and a great variety of ecological interactions. The tropical rainforest of Africa was the habitat in which the ancestors of humans evolved, and it is where the nearest surviving human relatives—chimpanzees and gorillas—live still. Tropical rainforests supplied a rich variety of food and other resources to indigenous peoples, who, for the most part, exploited this bounty without degrading the vegetation or reducing its range to any significant degree. However, in some regions a long history of forest burning by the inhabitants is thought to have caused extensive replacement of tropical rainforest and tropical deciduous forest with savanna.
Not until the past century, however, has widespread destruction of tropical forests occurred. Regrettably, tropical rainforests and tropical deciduous forests are now being destroyed at a rapid rate in order to provide resources such as timber and to create land that can be used for other purposes, such as cattle grazing. Today tropical forests, more than any other ecosystem, are experiencing habitat alteration and species extinction on a greater scale and at a more rapid pace than at any time in their history—at least since the major extinction event at the end of the Cretaceous Period some 65 million years ago (see ).
environment
The equatorial latitude of tropical rainforests and tropical deciduous forests keeps day length and mean temperature fairly constant throughout the year. The sun rises daily to a near-vertical position at noon, ensuring a high level of incoming radiant energy at all seasons. Although there is no cold season during which plants experience unfavourable temperatures that prohibit growth, there are many local variations in climate that result from topography, and these variations influence and restrict rainforest distribution within the tropics.
Tropical rainforests occur in regions of the tropics where temperatures are always high and where rainfall exceeds about 1,800 to 2,500 mm (about 70 to 100 inches) annually and occurs fairly evenly throughout the year. Similar hot climates in which annual rainfall lies between about 800 and 1,800 mm and in which a pronounced season of low rainfall occurs typically support tropical deciduous forests—i.e., rainforests in which up to about three-quarters of the trees lose their leaves in the dry season. The principal determining climatic factor for the distribution of rainforests in lowland regions of the tropics, therefore, is rainfall, both the total amount and the seasonal variation. Soil, human disturbance, and other factors also can be important controlling influences.
The climate is always hot and wet in most parts of the equatorial belt, but in regions to its north and south seasonal rainfall is experienced. During the summer months of the Northern Hemisphere—June to August—weather systems shift northward, bringing rain to regions in the northern parts of the tropics, as do the monsoon rains of India and Myanmar. Conversely, during the Southern Hemisphere's summer, weather systems move southward, bringing rain from December to February to places such as northern Australia. In these hot, seasonally wet areas grow tropical deciduous forests, such as the teak forests of Myanmar and Thailand. In other locations where conditions are similar but rainfall is not so reliable or burning has been a factor, savannas are found.
Topographic factors influence rainfall and consequently affect rainforest distribution within a region. For example, coastal regions where prevailing winds blow onshore are likely to have a wetter climate than coasts that experience primarily offshore winds. The west coasts of tropical Australia and South America south of the Equator experience offshore winds, and these dry regions can support rainforests only in very small areas. This contrasts with the more extensively rainforest-clad, east-facing coasts of these same continents at the same latitudes. The same phenomenon is apparent on a smaller scale where the orientation of coastlines is parallel to, rather than perpendicular to, wind direction. For example, in the Townsville area of northeastern Australia and in Benin in West Africa, gaps in otherwise fairly continuous tracts of tropical rainforest occur where the prevailing winds blow along the coast rather than across it.
Mean temperatures (temperature) in tropical rainforest regions are between 20 and 29 °C (68 and 84° F), and in no month is the mean temperature below 18 °C (64 °F). Temperatures become critical with increasing altitude; in the wet tropics temperatures fall by about 0.5 °C (0.9 °F) for every 100 metres (328 feet) climbed. Vegetation change across altitudinal gradients tends to be gradual and variable and is interpreted variously by different authorities. For example, in Uganda tropical rainforest grows to an altitude of 1,100 to 1,300 metres and has been described as giving way, via a transition forest zone, to montane rainforest above 1,650 to 1,750 metres, which continues to 2,300 to 3,400 metres. In New Guinea, lowland tropical rainforest reaches 1,000 to 1,200 metres, above which montane rainforests extend, with altitudinal variation, to 3,900 metres. In Peru, lowland rainforest extends upward to 1,200 to 1,500 metres, with transitional forest giving way to montane rainforest above 1,800 to 2,000 metres, which continues to 3,400 to 4,000 metres. These limits are comparable and reflect the similarities of climate in all regions where tropical rainforests occur. Plant species, however, are often quite different among regions.
Although the climate supporting tropical rainforests is perpetually hot, temperatures never reach the high values regularly recorded in drier places to the north and south of the equatorial belt. This is partly due to high levels of cloud cover, which limit the mean number of sunshine hours per day to between four and six. In hilly areas where air masses rise and cool because of the topography, the hours of sunlight may be even fewer. Nevertheless, the heat may seem extreme owing to the high levels of atmospheric humidity, which usually exceed 50 percent by day and approach 100 percent at night. Exacerbating the discomfort is the fact that winds are usually light; mean wind speeds are generally less than 10 km (6.2 miles) per hour and less than 5 km per hour in many areas. Devastating hurricanes (cyclones and typhoons) occur periodically in some coastal regions toward the margins of the equatorial belt, such as in the West Indies and in parts of the western Pacific region. Although relatively infrequent, such storms have an important effect on forest structure and regeneration.
 The climate within any vegetation ( microclimate) is moderated by the presence of plant parts that reduce incoming solar radiation and circulation of air. This is particularly true in tropical rainforests, which are structurally more dense and complex than other vegetation. Within the forest, temperature range and wind speed are reduced and humidity is increased relative to the climate above the tree canopy or in nearby clearings. The amount of rain reaching the ground is also reduced—by as much as 90 percent in some cases—as rainwater is absorbed by epiphytes (plants that grow on the surface of other plants but that derive nutrients and water from the air) and by tree bark or is caught by foliage and evaporates directly back to the atmosphere.
The climate within any vegetation ( microclimate) is moderated by the presence of plant parts that reduce incoming solar radiation and circulation of air. This is particularly true in tropical rainforests, which are structurally more dense and complex than other vegetation. Within the forest, temperature range and wind speed are reduced and humidity is increased relative to the climate above the tree canopy or in nearby clearings. The amount of rain reaching the ground is also reduced—by as much as 90 percent in some cases—as rainwater is absorbed by epiphytes (plants that grow on the surface of other plants but that derive nutrients and water from the air) and by tree bark or is caught by foliage and evaporates directly back to the atmosphere. Soils in tropical rainforests are typically deep but not very fertile, partly because large proportions of some mineral nutrients are bound up at any one time within the vegetation itself rather than free in the soil. The moist, hot climatic conditions lead to deep weathering of rock and the development of deep, typically reddish soil profiles rich in insoluble sesquioxides of iron and aluminum, commonly referred to as tropical red earths. Because precipitation in tropical rainforest regions exceeds evapotranspiration at almost all times, a nearly permanent surplus of water exists in the soil and moves downward through the soil into streams and rivers in valley floors. Through this process nutrients are leached out of the soil, leaving it relatively infertile. Most roots, including those of trees, are concentrated in the uppermost soil layers where nutrients become available from the decomposition of fallen dead leaves and other organic litter. Sandy soils, particularly, become thoroughly leached of nutrients and support stunted rainforests of peculiar composition. A high proportion of plants in this environment have small leaves that contain high levels of toxic or unpalatable substances. A variant of the tropical rainforest, the mangrove forest, is found along estuaries and on sheltered sea coasts in tidally inundated, muddy soils (see boundary ecosystem: Boundary systems between water and land: Coastal systems (boundary ecosystem)).
Soils in tropical rainforests are typically deep but not very fertile, partly because large proportions of some mineral nutrients are bound up at any one time within the vegetation itself rather than free in the soil. The moist, hot climatic conditions lead to deep weathering of rock and the development of deep, typically reddish soil profiles rich in insoluble sesquioxides of iron and aluminum, commonly referred to as tropical red earths. Because precipitation in tropical rainforest regions exceeds evapotranspiration at almost all times, a nearly permanent surplus of water exists in the soil and moves downward through the soil into streams and rivers in valley floors. Through this process nutrients are leached out of the soil, leaving it relatively infertile. Most roots, including those of trees, are concentrated in the uppermost soil layers where nutrients become available from the decomposition of fallen dead leaves and other organic litter. Sandy soils, particularly, become thoroughly leached of nutrients and support stunted rainforests of peculiar composition. A high proportion of plants in this environment have small leaves that contain high levels of toxic or unpalatable substances. A variant of the tropical rainforest, the mangrove forest, is found along estuaries and on sheltered sea coasts in tidally inundated, muddy soils (see boundary ecosystem: Boundary systems between water and land: Coastal systems (boundary ecosystem)).Even within the same area, however, there are likely to be signifiant variations in soil related to topographic position and to bedrock differences, and these variations are reflected in forest composition and structure. For example, as altitude increases—even within the same area and on the same bedrock—soil depth decreases markedly and its organic content increases in association with changes in forest composition and structure.
Biota
Only a minority of plant and animal species in tropical rainforests and tropical deciduous forests have been described formally and named. Therefore, only a rough estimate can be given of the total number of species contained in these ecosystems, as well as the number that are becoming extinct as a result of forest clearance. Nevertheless, it is quite clear that these vegetation types are the most diverse of all, containing more species than any other ecosystem. This is particularly so in regions in which tropical rainforests not only are widespread but also are separated into many small areas by geographic barriers, as in the island-studded Indonesian region (see biogeographic region: The distribution boundaries of flora and fauna (biogeographic region)). In this area different but related species often are found throughout various groups of islands, adding to the total regional diversity. Exceptionally large numbers of species also occur in areas of diverse habitat, such as in topographically or geologically complex regions and in places that are believed to have acted as refugia throughout the climatic fluctuations of the past few million years. According to some informed estimates, more than a hundred species of rainforest fauna and flora become extinct every week as a result of widespread clearing of forests by humans. Insects are believed to constitute the greatest percentage of disappearing species.
Flora (floristic region)
 All major groups of terrestrial organisms are represented abundantly in tropical rainforests. Among the higher plants, angiosperms (angiosperm) are particularly diverse and include many primitive forms and many families not found in the vegetation of other ecosystem types. Many flowering plants are large trees, of which there is an unparalleled diversity. For example, in one area of 23 hectares (57 acres) in Malaysia, 375 different tree species with trunk diameters greater than 91 cm (35.8 inches) have been recorded, and in a 50-hectare area in Panama, 7,614 trees belonging to 186 species had trunk diameters greater than 20 cm. New species of plants—even those as conspicuously large as trees—are found every year. Relatively few gymnosperms (gymnosperm) (conifers (conifer) and their relatives), however, are found in rainforests; instead, they occur more frequently at the drier and cooler extremes of the range of climates in which tropical rainforests grow. Some plant families, such as Arecaceae (palms (palm)), are typically abundant in all tropical rainforest regions, although different species occur from region to region. Other families are more restricted geographically. The family Dipterocarpaceae (dipterocarps) includes many massive trees that are among the most abundant and valuable species in the majority of tropical rainforests in western Malesia (see above Origin (tropical rainforest)); the family, however, is uncommon in New Guinea and Africa and absent from South and Central America and Australia. The Bromeliaceae (bromeliad) (bromeliads), a large family consisting mainly of rainforest epiphytes (epiphyte) and to which the pineapple belongs, is entirely restricted to the New World.
All major groups of terrestrial organisms are represented abundantly in tropical rainforests. Among the higher plants, angiosperms (angiosperm) are particularly diverse and include many primitive forms and many families not found in the vegetation of other ecosystem types. Many flowering plants are large trees, of which there is an unparalleled diversity. For example, in one area of 23 hectares (57 acres) in Malaysia, 375 different tree species with trunk diameters greater than 91 cm (35.8 inches) have been recorded, and in a 50-hectare area in Panama, 7,614 trees belonging to 186 species had trunk diameters greater than 20 cm. New species of plants—even those as conspicuously large as trees—are found every year. Relatively few gymnosperms (gymnosperm) (conifers (conifer) and their relatives), however, are found in rainforests; instead, they occur more frequently at the drier and cooler extremes of the range of climates in which tropical rainforests grow. Some plant families, such as Arecaceae (palms (palm)), are typically abundant in all tropical rainforest regions, although different species occur from region to region. Other families are more restricted geographically. The family Dipterocarpaceae (dipterocarps) includes many massive trees that are among the most abundant and valuable species in the majority of tropical rainforests in western Malesia (see above Origin (tropical rainforest)); the family, however, is uncommon in New Guinea and Africa and absent from South and Central America and Australia. The Bromeliaceae (bromeliad) (bromeliads), a large family consisting mainly of rainforest epiphytes (epiphyte) and to which the pineapple belongs, is entirely restricted to the New World.Tropical rainforests, which contain many different types of trees, seldom are dominated by a single species. A species can predominate, however, if particular soil conditions favour this occurrence or minimal disturbance occurs for several tree generations. Tropical deciduous forests are less diverse and often are dominated by only one or two tree species. The extensive deciduous forests of Myanmar, for example, cover wide areas and are dominated by only one or two tree species— teak (Tectona grandis) and the smaller leguminous tree Xylia xylocarpa. In Thailand and Indochina deciduous forests are dominated by members of the Dipterocarpaceae family, Dipterocarpus tuberculatus, Pentacme suavis, and Shorea obtusa.
Ferns, mosses, liverworts (liverwort), lichens (lichen), and algae are also abundant and diverse, although not as well studied and cataloged as the higher plants. Many are epiphytic and are found attached to the stems and sometimes the leaves of larger plants, especially in the wettest and most humid places. Fungi (fungus) and other saprophytic plants (vegetation growing on dead or decaying matter) are similarly diverse. Some perform a vital role in decomposing dead organic matter on the forest floor and thereby releasing mineral nutrients, which then become available to roots in the surface layers of the soil. Other fungi enter into symbiotic relationships with tree roots (mycorrhizae (mycorrhiza)).
Fauna (faunal region)
Interacting with and dependent upon this vast array of plants are similarly numerous animals. Like the plants, most animal species are limited to only one or a few types of tropical rainforest within an area, with the result that the overall number of species is substantially greater than it is in a single forest type. For example, a study of insects in the canopy of four different types of tropical rainforest in Brazil revealed 1,080 species of beetle, of which 83 percent were found in only one forest type, 14 percent in two, and only 3 percent in three or four types. While the larger, more conspicuous vertebrates (mammals, birds, and to a lesser degree amphibians and reptiles) are well known, only a small minority of the far more diverse invertebrates (particularly insects) have ever been collected, let alone described and named.
As with the plants, some animal groups occur in all tropical rainforest regions. A variety of fruit-eating parrots, pigeons, and seed-eating weevil beetles, for example, can be expected to occur in any tropical rainforest. Other groups are more restricted. Monkeys, while typical of tropical rainforests in both the New and the Old Worlds, are entirely absent from New Guinea and areas to its east and south. Tree kangaroos inhabit tropical rainforest canopies only in Australia and New Guinea, and birds of paradise (bird-of-paradise) are restricted to the same areas.
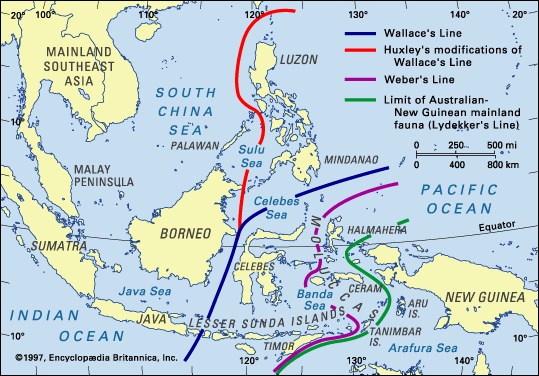 To a large extent these geographic variations in tropical rainforest biota reflect the long-term geologic histories of these ancient ecosystems. This is most clearly demonstrated in the Malesian phytogeographic subkingdom, which has existed as a single entity only since continental movements brought Australia and New Guinea northward into juxtaposition with Southeast Asia about 15 million years ago. Before that time the two parts were separated by a wide expanse of ocean and experienced separate evolution of their biota. Only a relatively small sea gap lies between them today; Java, Bali, and Borneo are on one side, and Timor and New Guinea are on the other, with islands such as Celebes and the Moluccas forming an intermediate region between. The gap is marked by a change in flora and, especially, fauna and is known as Wallace's Line (see biogeographic region: Fauna: Wallacea (biogeographic region)). The contrast is particularly stark with respect to mammals. To the west the rainforests are populated—or were populated until recently—by monkeys, deer, pigs, cats, elephants, and rhinoceroses, while those to the east have marsupial mammals, including opossums, cuscuses (cuscus), dasyurids (dasyurid), tree kangaroos, and bandicoots (bandicoot). Only a few groups such as bats and rodents have migrated across the line to become common in both areas. Similar contrasts, albeit less pronounced, can be seen in many other animal and plant groups across the same divide.
To a large extent these geographic variations in tropical rainforest biota reflect the long-term geologic histories of these ancient ecosystems. This is most clearly demonstrated in the Malesian phytogeographic subkingdom, which has existed as a single entity only since continental movements brought Australia and New Guinea northward into juxtaposition with Southeast Asia about 15 million years ago. Before that time the two parts were separated by a wide expanse of ocean and experienced separate evolution of their biota. Only a relatively small sea gap lies between them today; Java, Bali, and Borneo are on one side, and Timor and New Guinea are on the other, with islands such as Celebes and the Moluccas forming an intermediate region between. The gap is marked by a change in flora and, especially, fauna and is known as Wallace's Line (see biogeographic region: Fauna: Wallacea (biogeographic region)). The contrast is particularly stark with respect to mammals. To the west the rainforests are populated—or were populated until recently—by monkeys, deer, pigs, cats, elephants, and rhinoceroses, while those to the east have marsupial mammals, including opossums, cuscuses (cuscus), dasyurids (dasyurid), tree kangaroos, and bandicoots (bandicoot). Only a few groups such as bats and rodents have migrated across the line to become common in both areas. Similar contrasts, albeit less pronounced, can be seen in many other animal and plant groups across the same divide.Population and community development and structure
Tropical rainforests are distinguished not only by a remarkable richness of biota but also by the complexity of the interrelationships of all the plant and animal inhabitants that have been evolving together throughout many millions of years. As in all ecosystems, but particularly in the complex tropical rainforest community, the removal of one species threatens the survival of others with which it interacts. Some interactions are mentioned below, but many have yet to be revealed.
General structure of the rainforest
Plants with similar stature and life-form can be grouped into categories called synusiae, which make up distinct layers of vegetation. In tropical rainforests the synusiae are more numerous than in other ecosystem types. They include not only mechanically independent forms, whose stems are self-supporting, and saprophytic plants but also mechanically dependent synusiae such as climbers, stranglers, epiphytes, and parasitic plants. An unusual mix of trees of different sizes is found in the tropical rainforest, and those trees form several canopies below the uppermost layer, although they are not always recognizably separate layers. The upper canopy of the tropical rainforest is typically greater than 40 metres above ground.
 The tropical rainforest is structurally very complex (see figure-->). Its varied vegetation illustrates the intense competition for light (sunlight) that goes on in this environment in which other climatic factors are not limiting at any time of year and the vegetation is thus allowed to achieve an unequaled luxuriance and biomass. The amount of sunlight filtering through the many layers of foliage in a tropical rainforest is small; only about 1 percent of the light received at the top of the canopy reaches the ground. Most plants depend on light for their energy requirements, converting it into chemical energy in the form of carbohydrates by the process of photosynthesis in their chlorophyll-containing green tissues. Few plants can persist in the gloomy environment at ground level, and the surface is marked by a layer of rapidly decomposing dead leaves rather than of small herbaceous plants. Mosses grow on tree butts, and there are a few forbs such as ferns and gingers, but generally the ground is bare of living plants, and even shrubs are rare. However, tree seedlings and saplings are abundant; their straight stems reach toward the light but receive too little energy to grow tall enough before food reserves from their seeds are exhausted. Their chance to grow into maturity comes only if overhanging vegetation is at least partially removed through tree death or damage by wind. Such an occurrence permits more solar radiation to reach their level and initiates rapid growth and competition between saplings as to which will become a part of the well-lit canopy.
The tropical rainforest is structurally very complex (see figure-->). Its varied vegetation illustrates the intense competition for light (sunlight) that goes on in this environment in which other climatic factors are not limiting at any time of year and the vegetation is thus allowed to achieve an unequaled luxuriance and biomass. The amount of sunlight filtering through the many layers of foliage in a tropical rainforest is small; only about 1 percent of the light received at the top of the canopy reaches the ground. Most plants depend on light for their energy requirements, converting it into chemical energy in the form of carbohydrates by the process of photosynthesis in their chlorophyll-containing green tissues. Few plants can persist in the gloomy environment at ground level, and the surface is marked by a layer of rapidly decomposing dead leaves rather than of small herbaceous plants. Mosses grow on tree butts, and there are a few forbs such as ferns and gingers, but generally the ground is bare of living plants, and even shrubs are rare. However, tree seedlings and saplings are abundant; their straight stems reach toward the light but receive too little energy to grow tall enough before food reserves from their seeds are exhausted. Their chance to grow into maturity comes only if overhanging vegetation is at least partially removed through tree death or damage by wind. Such an occurrence permits more solar radiation to reach their level and initiates rapid growth and competition between saplings as to which will become a part of the well-lit canopy.Gaps in the canopy of a tropical rainforest provide temporarily well-illuminated places at ground level and are vital to the regeneration of most of the forest's constituent plants. Few plants in the forest can successfully regenerate in the deep shade of an unbroken canopy; many tree species are represented there only as a population of slender, slow-growing seedlings or saplings that have no chance of growing to the well-lit canopy unless a gap forms. Other species are present, invisibly, as dormant seeds in the soil. When a gap is created, seedlings and saplings accelerate their growth in the increased light and are joined by new seedlings sprouting from seeds stored in the soil that have been stimulated to germinate by light or by temperature fluctuations resulting from the sun's shining directly on the soil surface. Other seeds arrive by various seed-dispersal processes (see below). A thicket of regrowth rapidly develops, with the fastest-growing shrubs and trees quickly shading out opportunistic, light-demanding, low-growing herbaceous plants and becoming festooned with lianas. Through it all slower-growing, more shade-tolerant but longer-lived trees eventually emerge and restore the full forest canopy. The trees that initially fill in the gap in the canopy live approximately one century, whereas the slower-growing trees that ultimately replace them may live for 200 to 500 years or, in extreme cases, even longer. Detailed mapping of the trees in a tropical rainforest can reveal the locations of previous gaps through identification of clumps of the quicker-growing, more light-demanding species, which have yet to be replaced by trees in the final stage of successional recovery. Local, natural disturbances of this sort are vital to the maintenance of the full biotic diversity of the tropical rainforest (see Rainforest Regeneration).
 Just as tropical rainforest plants compete intensely for light above ground, below ground they vie for mineral nutrients. The process of decomposition of dead materials is of crucial importance to the continued health of the forest because plants depend on rapid recycling of mineral nutrients. bacteria and fungi (fungus) are primarily responsible for this process. Some saprophytic flowering plants that occur in tropical rainforests rely on decomposing material for their energy requirements and in the process use and later release minerals. Some animals are important in the decomposition process; for example, in Malaysia termites have been shown to be responsible for the decomposition of as much as 16 percent of all litter, particularly wood. Most trees in the tropical rainforest form symbiotic mycorrhizal (mycorrhiza) associations with fungi that grow in intimate contact with their roots; the fungi obtain energy from the tree and in turn provide the tree with phosphorus and other nutrients, which they absorb from the soil very efficiently. A mat of plant roots explores the humus beneath the rapidly decomposing surface layer of dead leaves and twigs, and even rotting logs are invaded by roots from below. Because nutrients are typically scarce at depth but, along with moisture, are readily available in surface layers, few roots penetrate very deeply into the soil. This shallow rooting pattern increases the likelihood of tree falls during storms, despite the support that many trees receive from flangelike plank buttresses growing radially outward from their trunk bases. When large trees fall, they may take with them other trees against which they collapse or to which they are tied by a web of lianas (liana) and thereby create gaps in the canopy.
Just as tropical rainforest plants compete intensely for light above ground, below ground they vie for mineral nutrients. The process of decomposition of dead materials is of crucial importance to the continued health of the forest because plants depend on rapid recycling of mineral nutrients. bacteria and fungi (fungus) are primarily responsible for this process. Some saprophytic flowering plants that occur in tropical rainforests rely on decomposing material for their energy requirements and in the process use and later release minerals. Some animals are important in the decomposition process; for example, in Malaysia termites have been shown to be responsible for the decomposition of as much as 16 percent of all litter, particularly wood. Most trees in the tropical rainforest form symbiotic mycorrhizal (mycorrhiza) associations with fungi that grow in intimate contact with their roots; the fungi obtain energy from the tree and in turn provide the tree with phosphorus and other nutrients, which they absorb from the soil very efficiently. A mat of plant roots explores the humus beneath the rapidly decomposing surface layer of dead leaves and twigs, and even rotting logs are invaded by roots from below. Because nutrients are typically scarce at depth but, along with moisture, are readily available in surface layers, few roots penetrate very deeply into the soil. This shallow rooting pattern increases the likelihood of tree falls during storms, despite the support that many trees receive from flangelike plank buttresses growing radially outward from their trunk bases. When large trees fall, they may take with them other trees against which they collapse or to which they are tied by a web of lianas (liana) and thereby create gaps in the canopy. Tree growth requires substantial energy investment in trunk development, which some plants avoid by depending on the stems of other plants for support. Perhaps the most obvious adaptation of this sort is seen in plants that climb from the ground to the uppermost canopy along other plants by using devices that resemble grapnel-like hooks. Lianas (liana) are climbers that are abundant and diverse in tropical rainforests; they are massive woody plants whose mature stems often loop through hundreds of metres of forest, sending shoots into new tree crowns as successive supporting trees die and decay. Climbing palms or rattans (Calamus) are prominent lianas in Asian rainforests, where the stems, which are used to make cane furniture, provide a valuable economic resource.
Tree growth requires substantial energy investment in trunk development, which some plants avoid by depending on the stems of other plants for support. Perhaps the most obvious adaptation of this sort is seen in plants that climb from the ground to the uppermost canopy along other plants by using devices that resemble grapnel-like hooks. Lianas (liana) are climbers that are abundant and diverse in tropical rainforests; they are massive woody plants whose mature stems often loop through hundreds of metres of forest, sending shoots into new tree crowns as successive supporting trees die and decay. Climbing palms or rattans (Calamus) are prominent lianas in Asian rainforests, where the stems, which are used to make cane furniture, provide a valuable economic resource.Epiphytes (epiphyte) are particularly diverse and include large plants such as orchids (orchid), aroids, bromeliads, and ferns in addition to smaller plants such as algae, mosses, and lichens. In tropical rainforests epiphytes are often so abundant that their weight fells trees. Epiphytes that grow near the upper canopy of the forest have access to bright sunlight but must survive without root contact with the soil. They depend on rain washing over them to provide water and mineral nutrients. During periods of drought, epiphytes undergo stress as water stored within their tissues becomes depleted. The diversity of epiphytes in tropical deciduous forests is much less than that of tropical rainforests because of the annual dry season (see ).
Parasitic (parasitism) flowering plants also occur. Hemiparasitic mistletoes (mistletoe) attached to tree branches extract water and minerals from their hosts but carry out their own photosynthesis. Plants that are completely parasitic also are found in tropical rainforests. Rafflesia (Rafflesiaceae), in Southeast Asia, parasitizes the roots of certain lianas and produces no aboveground parts until it flowers; its large orange and yellow blooms, nearly one metre in diameter, are the largest flowers of any plant.
Stranglers (strangler fig) make up a type of synusia virtually restricted to tropical rainforests. In this group are strangler figs (strangler fig) ( Ficus), which begin life as epiphytes, growing from seeds left on high tree branches by birds or fruit bats. As they grow, they develop long roots that descend along the trunk of the host tree, eventually reaching the ground and entering the soil. Several roots usually do this, and they become grafted together as they crisscross each other to form a lattice, ultimately creating a nearly complete sheath around the trunk. The host tree's canopy becomes shaded by the thick fig foliage, its trunk constricted by the surrounding root sheath and its own root system forced to compete with that of the strangling fig. The host tree is also much older than the strangler and eventually dies and rots away, leaving a giant fig “tree” whose apparent “trunk” is actually a cylinder of roots, full of large hollows that provide shelter and breeding sites for bats, birds, and other animals (see ). Stranglers may also develop roots from their branches, which, when they touch the ground, grow into the soil, thicken, and become additional “trunks.” In this way stranglers grow outward to become large patches of fig forest that consist of a single plant with many interconnected trunks.
Relationships between the flora and fauna
Some of the tallest trees and lianas, and the epiphytes they support, bear flowers and fruits at the top of the rainforest canopy, where the air moves unfettered by vegetation. They are able to depend on the wind for dispersal of pollen from flower to flower, as well as for the spreading of fruits and seeds away from the immediate environment of the parent plant (see “Flying” Trees). Ferns, mosses, and other lower plants also exploit the wind to carry their minute spores. However, a great many flowering plants, including many that grow in the nearly windless environment of the understory, depend on animals to perform these functions. They are as dependent on animals for reproductive success as the animals are on them for food—one example of the mutual dependence between plants and animals (see ).
 Many rainforest trees have sizable seeds (seed and fruit) from which large seedlings emerge and thrust their way through the thick mat of dead leaves on the dark forest floor. They develop tall stems, using food reserves in the seed without having to rely on sunlight, which is usually too dim, to meet their energy requirements. Because large seeds cannot be dispersed by the wind, these plants depend on a variety of animals to perform this function and have evolved many adaptations to encourage them to do so. Fruit bats (Old World fruit bat) are attracted by fragrant, sweet fruits typically borne conspicuously and conveniently on the outer parts of the tree canopy; the mango (Mangifera indica), native to the rainforests of India, provides a good example. The bats not only feed on fruits as they hang from the trees but also may carry a fruit away to another perch, where they eat the flesh and drop the seed. Smaller fruits may be swallowed whole, the seeds passing through the gut intact and being voided at a distance. The ground beneath trees used by fruit bats as a roost is commonly thick with seedlings of fleshy, fruit-bearing trees (see ).
Many rainforest trees have sizable seeds (seed and fruit) from which large seedlings emerge and thrust their way through the thick mat of dead leaves on the dark forest floor. They develop tall stems, using food reserves in the seed without having to rely on sunlight, which is usually too dim, to meet their energy requirements. Because large seeds cannot be dispersed by the wind, these plants depend on a variety of animals to perform this function and have evolved many adaptations to encourage them to do so. Fruit bats (Old World fruit bat) are attracted by fragrant, sweet fruits typically borne conspicuously and conveniently on the outer parts of the tree canopy; the mango (Mangifera indica), native to the rainforests of India, provides a good example. The bats not only feed on fruits as they hang from the trees but also may carry a fruit away to another perch, where they eat the flesh and drop the seed. Smaller fruits may be swallowed whole, the seeds passing through the gut intact and being voided at a distance. The ground beneath trees used by fruit bats as a roost is commonly thick with seedlings of fleshy, fruit-bearing trees (see ).A variety of birds (bird) eat fleshy fruits also, voiding or regurgitating the unharmed seeds. Birds of different sizes are typically attracted to similarly scaled fruits, which are carried on stems of appropriate thickness and strength. For example, large pigeons in New Guinea feed preferentially on larger fruits borne on thicker stems that can bear not only the weight of the fruit but also the weight of the large bird; smaller pigeons tend to feed on smaller fruits borne on thinner twigs. In such a manner, the diverse plant community is matched by a similarly diverse animal community in interdependence.
Terrestrial mammals also help to disperse seeds. In many cases this has favoured the positioning of flowers and fruits beneath the canopy on the trunks of trees accessible to animals unable to climb or fly, an adaptation called cauliflory. In some cases fruits are grown in the canopy but drop as they ripen, opening only after they fall to attract ground-dwelling animals that will carry them away from the parent tree. The durian fruit Durio zibethinus of Southeast Asian rainforests is an example; its fruits are eaten and its seeds dispersed by a range of mammals, including pigs, elephants, and even tigers.
Many other animals, from ants to apes, are involved in seed dispersal. In the Amazon basin of Brazil, where large areas of tropical rainforest are seasonally flooded, many trees produce fruit attractive to fish, which swallow them whole and void the seeds (see ). Squirrels are also important seed dispersers in parts of South America. In the tropical rainforests of northeastern Australia, cassowaries (cassowary) are responsible for generating mixed clumps of tree seedlings of several species that grow from their dung sites.
It is important for seeds to be spread away from parent plants, both to allow seedlings to escape competition with the parent and to expand the range of the species. Another capacity important to seed survival, particularly in the diverse tropical rainforest community, involves the evasion of seed predators. Many different beetles and other insects are specialized to feed on particular types of seed. Seeds concentrated beneath a parent plant are easy for seed predators to locate. Seeds that are carried away to areas occupied by different plant species—and different seed predators—are more likely to survive (see ).
In addition to dispersing seeds, animals are vital to tropical rainforest reproduction through flower pollination. Many insects such as bees, moths, flies, and beetles as well as birds and bats carry out this activity. Birds such as the hummingbirds (hummingbird) of South and Central America and the flower-peckers (flowerpecker) of Asia have adaptations that allow them to sip nectar from flowers. In the process they inadvertently become dusted with pollen, which they subsequently transport to other flowers, pollinating them. The plants involved also show special adaptations in flower structure and colour. Most flowers pollinated by birds are red, a colour highly visible to these animals, whereas flowers pollinated by night-flying moths are white or pink, and those pollinated by insects that fly during the day are often yellow or orange. Bats are important pollinators of certain pale, fragrant flowers that open in the evening in Asian rainforests (see also Chiropterophilous Plants).
Biological productivity
Of all vegetation types, tropical rainforests grow in climatic conditions that are least limiting to plant growth. It is to be expected that the growth and productivity (total amount of organic matter produced per unit area per unit time) of tropical rainforests would be higher than that of other vegetation, provided that other factors such as soil fertility or consumption by herbivorous animals are not extremely low or high.
Various methods are employed to assess productivity. Gross primary productivity is the amount of carbon fixed during photosynthesis by all producers in the ecosystem. However, a large part of the harnessed energy is used up by the metabolic processes of the producers ( respiration). The amount of fixed carbon not used by plants is called net primary productivity, and it is this remainder that is available to various consumers in the ecosystem—e.g., the herbivores, decomposers, and carnivores. Of course, in any stable ecosystem there is neither an accumulation nor a diminution in the total amount of organic matter present, so that overall there is a balance between the gross primary productivity and the total consumption. The amount of organic matter in the system at any point in time, the total mass of all the organisms present, is called the biomass. (For further discussion of productivity, see biosphere: Resources of the biosphere (biosphere).)
The biomass of tropical rainforests is larger than that of other vegetation. It is not an easy quantity to measure, involving the destructive sampling of all the plants in an area (including their underground parts), with estimates made of the mass of other organisms belonging to the ecosystem such as animals. Measurements show that tropical rainforests typically have biomass values on the order of 400 to 700 metric tons per hectare, greater than most temperate forests and substantially more than other vegetation with fewer or no trees. A measurement of biomass in a tropical deciduous forest in Thailand yielded a value of about 340 metric tons per hectare.
Increase in biomass over the period of a year at one rainforest site in Malaysia was estimated at 7 metric tons per hectare, while total litter fall was 14 metric tons, estimated mass of sloughed roots was 4 metric tons, and total live plant matter eaten by herbivorous animals (both invertebrate and vertebrate) was about 5 metric tons per hectare per year. These values add up to a total net production of 30 metric tons per hectare per year. Respiration by the vegetation itself was estimated at 50 metric tons, so that gross primary productivity was about 80 metric tons per hectare per year. Compared with temperate forests, these values are approximately 2.5 times higher for net productivity and 4 times higher for gross productivity, the difference being that the respiration rate at the tropical site was 5 times that of temperate forests.
Despite the overall high rates of productivity and biomass in tropical rainforests, the growth rates of their timber trees are not unusually fast; in fact, some temperate trees and many smaller herbaceous plants grow more rapidly. The high productivity of tropical rainforests instead results in their high biomass and year-round growth. They also have particularly high levels of consumption by herbivores (see ), litter production, and especially plant respiration.
Additional Reading
P.W. Richards, The Tropical Rain Forest (1952), is perhaps the best early account, containing a mine of useful information. Kathlyn Gay, Rainforests of the World: A Reference Handbook (1993), describes the interaction of rainforests and climate. Regional accounts of value are T.C. Whitmore, Tropical Rain Forests of the Far East, 2nd ed. (1984); and Paul Adam, Australian Rainforests (1992).
- Voreqe Bainimarama
- Vorkuta
- Vorobyev, Arkady
- Voronezh
- Vorontsov, Mikhail Illarionovich
- Vorontsov, Mikhail Semyonovich, Prince
- Voroshilov, Kliment Yefremovich
- Vorschule
- Vorskla River, Battle of the
- Vorster, John
- Vorticella
- Vortigern
- vortograph
- voršud
- Vosges
- Voskhod
- Voskresensk
- Vossius, Gerardus Johannes
- Voss, Johann Heinrich
- Vostochno-Kazakhstan
- Vostok
- Vostok Island
- vote
- vote of confidence
- Votic language