基因调控(卷名:生物学)
regulation of gene expression
━━━━━━━━━━━━━━━━━━━━━━━━
简史
研究方法
筛选突变型
激素诱导
离体生化方法
重组DNA技术
原核生物的基因调控
DNA水平上的基因调控
操纵子模型
真核生物的基因调控
染色体DNA水平上的基因调控
转录水平上的基因调控
RNA加工过程中的调控
翻译控制
翻译后控制
实用意义━━━━━━━━━━━━━━━━━━━━━━━━
生物体内控制基因表达的机制。基因表达的主要过程是基因的转录和信使核糖核酸(mRNA)的翻译。基因调控主要发生在三个水平上,即①DNA水平上的调控、转录控制和翻译控制;②微生物通过基因调控可以改变代谢方式以适应环境的变化,这类基因调控一般是短暂的和可逆的;③多细胞生物的基因调控是细胞分化、形态发生和个体发育的基础,这类调控一般是长期的,而且往往是不可逆的。基因调控的研究有广泛的生物学意义,是发生遗传学和分子遗传学的重要研究领域。
通过基因调控,微生物可以避免过多地合成氨基酸、核苷酸之类物质。如果使它们的调节基因发生突变,就可以得到大量合成这些物质的菌种,把这些菌种用在发酵工业上,使产量大幅度增长。在遗传工程的研究中应用基因调控的原理可使外源基因表达(见重组DNA技术),所以基因调控的理论探讨还具有生产实践意义。
简 史
1900年F.迪纳特发现在含有乳糖和半乳糖的培养液中培养的酵母菌细胞中有分解半乳糖的酶,但是在葡萄糖的培养液中培养的酵母菌细胞中没有相应的酶。1930年H.卡尔斯特伦在关于细菌的研究中也发现类似的现象,并把生物细胞中的酶区分为组成酶和适应酶两类,前者是在任何情况下都存在的酶,后者是只在诱导物(一般即作用底物)存在的情况下才出现的酶。1938年J.尤德金用质量作用定律解释适应酶的出现。1946年法国分子生物学家J.莫诺开始研究与大肠杆菌的乳糖发酵有关的酶的诱导合成现象。同年美国微生物遗传学家J.莱德伯格等发现细菌接合的现象,接着在1948年分离得到大量的不能利用乳糖的大肠杆菌的突变型,也分离到不接触乳糖也能在细胞中出现分解乳糖的 β-半乳糖苷酶的类型,亦即失去基因调控能力的突变型。在对大肠杆菌遗传学研究的基础上,J.莫诺和F.雅各布等对大肠杆菌的乳糖发酵中的酶和一系列突变型继续进行了广泛深入的研究,终于在1960~1961年提出了乳糖操纵子模型,开创了基因调控机制的研究。1967年美国学者W.吉尔伯特等分离得到阻遏蛋白,1969年美国分子遗传学家J.夏皮罗等分离得到乳糖操纵子,使基因调控的研究逐渐成为分子遗传学的一个重要内容。研究方法
筛选突变型 这是在原核生物中广泛应用的方法,例如在乳糖操纵子的研究中筛选失去了基因调控能力的组成型,包括调节基因发生突变和操纵基因发生突变的突变型,以及筛选即使有乳糖或其他诱导物存在的情况下仍然不能合成β-半乳糖糖苷酶的超阻遏型等等。激素诱导 在高等的真核生物中,除了离体培养的体细胞以外(见体细胞遗传学),一般较难得到上述那些突变型。因此常利用激素能诱导产生特定蛋白质的现象来研究编码这些蛋白质的基因调控。
离体生化方法 真核生物的染色体主要由 DNA和蛋白质组成。所以分别抽提这些成分然后逐一添加其他成分再进行离体转录或离体翻译的测定,这也是真核生物基因调控研究中经常采用的研究手段。
分子杂交、电镜观察以及各种一般的生物化学方法都是基因调控的重要研究手段。
重组DNA技术 许多基因,包括只在某一发育阶段活动的基因(见基因文库)都可以从基因组中分离出来进行研究。应用核酸的顺序分析技术可以测定基因的核苷酸顺序。再应用体内与体外的基因表达系统就能直接了解基因调控的机制。
原核生物的基因调控
DNA水平上的基因调控 鼠伤寒沙门氏菌(Salmoella typhimurium)有两个编码鞭毛蛋白的基因H1和H2,这两个基因并不紧密连锁。H2 的一边有一个调节基因 ( H1 repressor gene,rh1),它所编码的阻遏蛋白作用于 H1而使它不表达。H2基因的另一边有一段经常发生倒位的长约970对核苷酸的DNA序列,它的一端包括一个启动子(P)。如果这一序列的取向使P和H2、rh1邻接,这两个基因便得以转录,细胞中出现H2基因所编码的一种鞭毛蛋白和rh1基因所编码的阻遏蛋白,阻遏蛋白作用于 H1基因而使它不表达,细菌的鞭毛于是由H2蛋白构成。当这一序列发生倒位而呈另一种取向,使P远离H2和rh1,于是H2所编码的鞭毛蛋白和阻遏蛋白都停止合成,H1便得以表达,细菌的鞭毛便变为由H1蛋白所构成。鞭毛蛋白就是细菌的鞭毛抗原。这种抗原由一种状态(称为相)变为另一状态的现象称为相转变。类似的现象也出现在大肠杆菌的 Mu-1 噬菌体中。Mu-1 噬菌体的染色体上有一段长约 3000对核苷酸的核苷酸序列(称为 G),这一序列也经常发生倒位。它的一个取向使细胞中合成的噬菌体外壳蛋白是 U和 S,这种噬菌体能感染大肠杆菌,可是不能感染弗氏柠檬细菌(Citrobacter freundii)。它的另一取向则使细胞中合成的噬菌体的外壳蛋白是U′和S′,这种噬菌体能感染弗氏柠檬细菌,可是不能感染大肠杆菌。
操纵子模型 细菌和噬菌体的基因调控多数发生在转录水平上,操纵子是细菌的主要的基因调控单位,也就是转录单位。大肠杆菌的乳糖操纵子是第一个被发现的典型的操纵子,它包括依次排列着的启动子(promoter,P)、操纵基因(operator,lac O)和三个结构基因。结构基因lac Z编码分解乳糖的 β-半乳糖苷酶、lac Y编码吸收乳糖的β-半乳糖苷透性酶,lac A编码β-半乳糖苷乙酰转移酶。紧靠着lac Z是操纵基因lacO,它不编码任何蛋白质,它是另一位置上的调节基因lac I所编码的阻遏蛋白的结合部位。阻遏蛋白是一种变构蛋白,当细胞中有乳糖或其他诱导物的情况下阻遏蛋白便和它们相结合,这一结合使阻遏蛋白的构象变为不能结合在lac O上,于是转录便得以进行,吸收和分解乳糖的酶便得以产生(图la);如果细胞中没有乳糖或其他诱导物则阻遏蛋白就结合在lac O上,从而阻止了结合在旁边的启动子P上的RNA多聚酶的前进道路,使转录不能进行(图1b)。通过这种方式的基因调控能使细菌在有乳糖的环境中合成有关利用乳糖的酶,在没有乳糖的环境中停止合成这些酶。
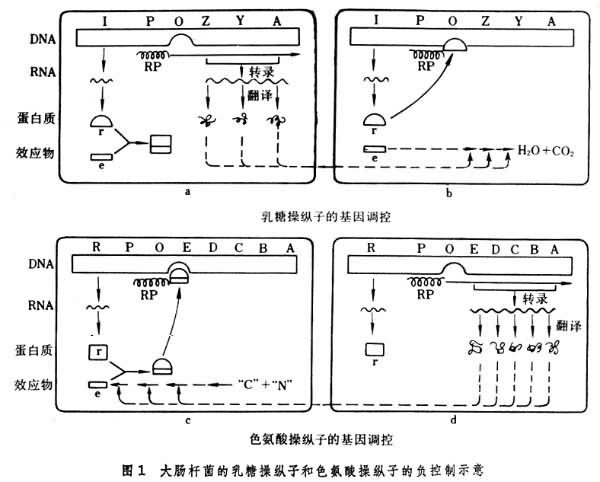 一个操纵子是一个整体,因为:①从结构上看来,一个操纵子的全部基因都排列在一起;②一个操纵子虽然包括若干个结构基因,可是通过转录形成的却是单个多基因mRNA;③属于一个操纵子的结构基因所编码的蛋白质总是按一定的比率合成;④一个操纵子中的靠近操纵基因的结构基因如果发生某些突变则后面的一系列基因所编码的蛋白质的量便减少。这种现象称为极性效应,这种突变型称为极性突变型;⑤操纵基因和结构基因间有顺反位置效应(见互补作用)。
一个操纵子是一个整体,因为:①从结构上看来,一个操纵子的全部基因都排列在一起;②一个操纵子虽然包括若干个结构基因,可是通过转录形成的却是单个多基因mRNA;③属于一个操纵子的结构基因所编码的蛋白质总是按一定的比率合成;④一个操纵子中的靠近操纵基因的结构基因如果发生某些突变则后面的一系列基因所编码的蛋白质的量便减少。这种现象称为极性效应,这种突变型称为极性突变型;⑤操纵基因和结构基因间有顺反位置效应(见互补作用)。乳糖操纵子的基因lacI所编码的阻遏蛋白所起的是负控制作用,当它作用于操纵基因时转录的进行被阻止。另一个调节基因称为环腺苷受体蛋白基因 (cyclic AMP receptor protein gene,CRP),它所编码的CRP蛋白质和环腺苷酸(cAMP)相结合并以结合状态作用于启动子时便能促进转录的进行。这种基因调控属于正控制。葡萄糖的降解物能降低细胞中的cAMP的浓度,cAMP浓度降低后CRP蛋白便不再和它结合,从而改变构象并失去了促进转录的功能。这一效应称为葡萄糖效应。因此 CRP蛋白又称为降解物激活蛋白,CRP基因又称为CAP基因。由此可见乳糖操纵子既受到负控制,也受到正控制。这种双重的基因调控同样符合于细菌的需要:在有乳糖的情况下细菌需要合成、吸收和分解乳糖的酶,可是在同时存在葡萄糖和乳糖的情况下细菌就没有必要再去利用乳糖,所以也就没有必要合成吸收和分解乳糖的酶。
相类似的基因调控也作用于合成代谢。例如在细菌自身合成或从外面供给了足够量的色氨酸的情况下,色氨酸便和特定的调节基因所编码的阻遏蛋白相结合,结合状态的阻遏蛋白作用于操纵基因,从而转录被阻止,合成色氨酸所必需的酶便停止合成(图1c)。在色氨酸用于合成蛋白质而减少的情况下,游离状态的阻遏蛋白便不再作用于操纵基因,于是转录便能进行,酶便合成,色氨酸便又通过这些酶的作用而产生(图1d)。色氨酸操纵子是色氨酸合成代谢中的基因调控单位,也是一个整体,具有和乳糖操纵子相同的特性。
细菌的基因调控还有其他形式,但是乳糖操纵子是它的基本模式。调控方式有正控制和负控制,且往往正负控制同时作用于一个操纵子。这种多重控制比单一控制更有效。在大肠杆菌中已知的操纵子已超过100。
真核生物的基因调控
真核生物的基因调控比原核生物复杂得多。这是因为这两类生物在三个不同水平上存在着重大的差别:①在遗传物质的分子水平上,真核细胞基因组的 DNA含量和基因的总数都远高于原核生物,而且 DNA不是染色体中的唯一成分,DNA和蛋白质以及少量的RNA构成以核小体为基本单位的染色质;②在细胞水平上,真核细胞的染色体包在核膜里面,转录和翻译分别发生在细胞核和细胞质中,这两个过程在时间上和空间上都是分开的,而且在转录和翻译之间存在着一个相当复杂的 RNA加工过程;③在个体水平上,真核生物是由不同的组织细胞构成的,从受精卵到完整个体要经过复杂的分化发育过程,除了那些为了维持细胞的基本生命活动所必需的基因之外,其他不同组织的细胞中的基因总是在不同的时空序列中被活化或受阻遏。与分化发育有关的基因调控机制是发生遗传学研究的主要内容。染色体 DNA水平上的基因调控 通过改变基因组中有关基因的数量和顺序结构而实现的基因调控。
染色质丢失 在发育过程中一些体细胞失去了某些基因,这些基因便永不表达,这是一种极端形式的不可逆的基因调控。
在某些线虫、原生动物、甲壳动物发育过程中的体细胞有遗传物质丢失现象。在这些生物中,只有生殖细胞才保留着该种生物基因组的全套基因。例如在马副蛔虫 (Ascaris megacephala)卵裂的早期就发现有染色体的丢失现象。蜜蜂的工蜂和蜂后是二倍体,而单倍体则发育成为雄蜂。这也可以认为是一种通过染色体丢失的基因调控。
基因扩增 另一种改变基因数量而调节基因表达的方式称为基因扩增。基因扩增是细胞短期内大量产生出某一基因拷贝的一种非常手段。某些脊椎动物和昆虫的卵母细胞能够专一性地增加编码核糖体RNA的DNA(rDNA)序列。例如非洲爪蟾(Xenopus laevis)的卵母细胞中的rDNA的拷贝数可由平时的 1500急剧增加至2000000。这一基因扩增仅发生在卵母细胞中,它适应于胚胎发育中对于大量核糖体的需要。当胚胎期开始时这些染色体外的rDNA拷贝即失去功能并逐渐消失。
除了rDNA的专一性扩增以外,还发现果蝇的卵巢囊泡细胞中的绒毛膜蛋白质基因在转录之前也先进行专一性的扩增。通过这一手段,细胞在很短的时间内积累起大量的基因拷贝,从而合成出大量的绒毛膜蛋白质。
染色体上DNA顺序的重排 改变基因组中有关基因顺序结构的基因调控方式。哺乳动物的免疫球蛋白的可变区与恒定区的顺序分别由不同的基因片段编码。它们处于同一染色体上但是相距较远,中间还有一些编码连接区的DNA顺序。在产生抗体的浆细胞成熟过程中,这三个序列通过染色体重排而成为一个完整的转录单位。由于可变区基因片段为数众多,而且不同的连接方式又带来相应的核苷酸顺序的变化,所以通过这种形式的 DNA重排可以产生种类繁多的免疫球蛋白基因(见免疫遗传学)。
在啤酒酵母(Saccharomyces cerevisiae)的细胞中有关接合型的两个基因 a和 α究竟哪一个表达取决于一种特殊的转座因子的移位。但就大多数已经发现的真核生物的转座因子来讲,它们是否参与正常的基因调控现在还没有定论。
转录水平上的基因调控 真核生物的基因调控主要表现在对基因转录活性的控制上而不涉及 DNA序列在数量和结构上的改变。转录水平上的基因调控可以通过不同的途径实现。
异染色质化和染色质的活化 染色质处在固缩的状态称为异染色质化。在异染色质化部位的基因的转录活性显著降低。真核生物可以改变染色体某一区域的异染色质化的程度而控制基因的表达。雌性哺乳动物细胞中的一个 X染色体的失活便是高度异染色质化的结果(见剂量补偿效应)。基因由于改变位置而处在异染色质区附近时,转录作用也会受到阻碍(见位置效应)。粉蚧科的一种介壳虫有两类个体,一类个体的细胞中的10个染色体都没有异染色质化,因而都是有功能的;另一类个体细胞的 10个染色体中有5个高度异染色质化,因而是没有功能的。这就造成了和蜜蜂相似的情况,蜜蜂的二倍体是雌性的,单倍体是雄性的;在这里则前一类个体是雌性的,而后一类个体是雄性的。
与异染色质化相反的情况是染色质的活化。在活化的染色质中,基因 DNA以某种不同的方式装配到染色质的核小体中,使RNA多聚酶能够转录染色质中的DNA。研究结果表明凡有基因表达活性的染色质 DNA对核酸内切限制酶的降解作用比没有转录活性的染色质要敏感得多。例如鸡的网织细胞能大量合成珠蛋白,从这种细胞中分离的含珠蛋白基因的染色质DNA易遭受DNA酶I的降解作用,而大多数其他的染色质 DNA则不被降解,在不合成珠蛋白的鸡输卵管细胞中含珠蛋白基因的染色质 DNA也不被这种酶降解。在活化的染色质中 DNA的超螺旋状态有所改变,这是基因活化的前提。
修饰作用 真核细胞修饰 DNA的主要途径是胞嘧啶(c)在5位上的甲基化反应。5-甲基胞嘧啶通常位于鸟嘌呤(G)的旁边。可见 GC顺序最容易被甲基化。在刚刚完成复制的 DNA分子中只有母链(模板链)是甲基化的。新生 DNA链的甲基化在母链的指导下进行。用限制酶进行分析的结果表明在不转录的DNA中的GC有 70%以上是甲基化的,而在表达活性高的DNA中,GC顺序只有20~30%是甲基化的。这意味着 DNA甲基化的作用也是一种基因调控手段。
蛋白质也可以被修饰,修饰作用包括乙酰化、磷酸化等。除了组蛋白和染色体 DNA牢固结合以外,许多非组蛋白也可以和 DNA相结合,对这些蛋白质的修饰作用同样能改变它们与 DNA的结合方式,并改变染色质和核小体的结构,从而影响基因的转录活性。某些非组蛋白成分还能和激素相结合而激活某些基因。此外,RNA聚合酶也可以由于被修饰而改变活性。
激素和基因转录的诱导 细菌的代谢作用直接受环境的影响,它的基因调控的信号常来自环境因素。多细胞的高等生物的代谢作用较少为环境所影响,它的基因调控的信号常来自体内的激素。
在摇蚊(Chironomus)和果蝇(Drosophila)等双翅目昆虫的唾腺中的巨大的多线染色体上可以看到一条条各有特征的横纹。在幼虫和蛹期的各个发育时期可以看到某些横纹变得疏松膨大,这膨大处称为疏松区。疏松区的出现有一定的时间表,而且各个疏松区出现以后隔一定时间又消失。这些部位是合成大量RNA的部位,而且通过分子杂交可以证明疏松区的成分具有mRNA的性质。用蜕皮激素处理幼虫或离体的唾腺细胞,可以诱发某些横纹形成疏松区,意味着某些基因被激活。这是激素诱发特定基因的转录的最为直观的证据。
在高等动物中,注射雌性激素能促使公鸡或小鸡的肝脏细胞中产生卵黄蛋白原mRNA并合成卵黄蛋白原;注射孕酮能促使爬行动物或鸟类的输卵管细胞产生卵清蛋白mRNA并合成卵清蛋白;脑垂体前叶的促乳腺激素能促使哺乳动物的乳腺细胞合成酪蛋白。
甾体激素作用的机制一般认为是这样的,它首先和靶细胞的细胞质中的受体蛋白结合成为激素和受体的复合物,然后这一复合物进入细胞核,在染色体上有某些非组蛋白存在的情况下,复合物便能结合在染色体的特定位置上,从而促使特定基因转录。
RNA多聚酶的分工 与原核生物不同,真核生物有三种不同的 RNA多聚酶,它们各自负责不同类型的基因的转录。从表中不难看出由RNA多聚酶Ⅰ和Ⅲ转录的RNA都与所有细胞的生命活动的基本功能──翻译有关,而只有 RNA多聚酶Ⅱ才能转录结构基因而进一步产生蛋白质。显然这种分工反映了这三类基因在表达机制上的重大差别(见表)。
 在转录启动时,不同的RNA多聚酶能识别不同类型的基因,识别的机制在于每种类型的基因都有共同或类似的调控顺序。在活体和离体的实验中已经证明在 RNA多聚酶Ⅱ的离转录起点的上游(5′方向)约 25~30个核苷酸处,有一段长约 8~10个核苷酸的相当保守的、富含A(腺嘌呤核苷酸)和T(胸腺嘧啶核苷酸)的核苷酸顺序,称为TATA框。有证据表明在转录起始的上游更远的部位,还有其他核苷酸顺序也与 RNA多聚酶Ⅱ的正常活动有关。
在转录启动时,不同的RNA多聚酶能识别不同类型的基因,识别的机制在于每种类型的基因都有共同或类似的调控顺序。在活体和离体的实验中已经证明在 RNA多聚酶Ⅱ的离转录起点的上游(5′方向)约 25~30个核苷酸处,有一段长约 8~10个核苷酸的相当保守的、富含A(腺嘌呤核苷酸)和T(胸腺嘧啶核苷酸)的核苷酸顺序,称为TATA框。有证据表明在转录起始的上游更远的部位,还有其他核苷酸顺序也与 RNA多聚酶Ⅱ的正常活动有关。在由RNA多聚酶Ⅲ转录的5SRNA基因的内部(而不是在它的转录起点的上游)有一段与转录控制有关的顺序,长约30碱基对,它的存在对该基因转录的起始起着控制作用。已经证明这段顺序能专一性地结合一种蛋白质因子,后者可以指导 RNA多聚酶Ⅲ从它的结合位置的上游约50碱基对处开始转录。
RNA加工过程中的调控 真核生物的RNA加工过程主要包括三个步骤:①在新生RNA的5′端加上一个甲基化的鸟嘌呤核苷酸,形成一个所谓的帽子(cap)即m7GpppN(m7G是7-甲基鸟嘌呤核苷,P是磷酸,N是 RNA的5′端第一个核苷酸)这一过程通常发生在新生链完成之前。②在转录后的RNA3′部位上加上多聚腺嘌呤核苷酸(多聚A)尾部。这种加尾作用一般不直接发生在转录初产物的3′末端上,而另外需要核酸内切酶的作用产生一个新的3′末端,然后再加上多聚A。③对于具有内含子的那部分 RNA顺序必须被切除,接着两边的外显子再重新连起来,这一过程称为拼接。拼接是个十分精确的过程,它的机制还没有阐明,但几乎所有的内含子在5′边界处都有CT顺序,在3′边界处都有AG顺序。多聚A加尾作用一般发生在拼接之前,但不总是如此。现在还不清楚影响这个过程的各种因素,但已经知道同一种基因的转录产物前体mRNA可以被加工成几种不同的mRNA。
几乎所有的真核生物的 mRNA都有一个5′帽端,但并不是所有基因的mRNA都有3′多聚A尾部,也不是所有基因的mRNA都必须经过拼接。根据这后两种加工过程的有无和复杂程度,可将真核基因的转录单位分为两大类型:一类是简单的只编码产生一种蛋白质的基因,另一类是复杂的编码两种或更多种蛋白质的转录单位(图2)。在3′端加尾部位或拼接部位上的变化都会使同一基因最终形成不同的蛋白质产物。图2a是表示多聚 A加尾部位不同而使基因终产物不同的模式,属于这种情况的有腺病毒的晚期基因、小鼠免疫球蛋白的重链基因和一种多肽激素降血钙素基因。图2b是表示拼接部位不同而使基因终末产物不同的模式,迄今为止仅在哺乳动物细胞的病毒(如腺病毒、SV40病毒、多瘤病毒和反转录病毒等)中发现有这种形式的加工调控。
 翻译控制 真核生物的翻译控制的主要形式是控制mRNA的稳定性。mRNA5′端的加帽作用以及它的3′端的多聚A的加尾作用都有助于 mRNA分子的稳定。在某些真核生物中mRNA进入细胞质以后并不立即作为模板进行蛋白质合成,而是与一些蛋白质结合形成RNA蛋白质(RNP)颗粒,在这种状态的mRNA半衰期可以延长。家蚕的丝芯蛋白基因是单拷贝的,可是在几天内一个细胞中可以合成多至1010个丝芯蛋白分子。这便是因为它的mRNA分子和蛋白质结合成为 RNP颗粒而延长了寿命的结果。据估计,一个丝芯蛋白基因在几天内可产生105个mRNA分子,因此每个mRNA分子作为模板可以合成105个蛋白质分子。
翻译控制 真核生物的翻译控制的主要形式是控制mRNA的稳定性。mRNA5′端的加帽作用以及它的3′端的多聚A的加尾作用都有助于 mRNA分子的稳定。在某些真核生物中mRNA进入细胞质以后并不立即作为模板进行蛋白质合成,而是与一些蛋白质结合形成RNA蛋白质(RNP)颗粒,在这种状态的mRNA半衰期可以延长。家蚕的丝芯蛋白基因是单拷贝的,可是在几天内一个细胞中可以合成多至1010个丝芯蛋白分子。这便是因为它的mRNA分子和蛋白质结合成为 RNP颗粒而延长了寿命的结果。据估计,一个丝芯蛋白基因在几天内可产生105个mRNA分子,因此每个mRNA分子作为模板可以合成105个蛋白质分子。某些激素对mRNA也能起稳定作用。例如在离体培养的乳腺组织中添加催乳激素能使酪蛋白的mRNA分子在24小时内积累到25000个拷贝。在这期间mRNA的半衰期增加了20倍,但新的mRNA合成只增加2~3倍。如果在培养液中再除去催乳激素,48小时内酪蛋白mRNA便丧失了95%,可见该激素的主要作用在于维持酪蛋白mRNA的稳定性。
翻译控制的另外两种形式是对翻译速度的控制和进行有选择的翻译,例如,海胆未受精的卵与受精的卵在mRNA的含量和组成上都相同,但是受精卵的翻译活性至少高出50倍。和这情况不相同的是蚶的卵,受精卵蛋白质合成的总量并没有提高,可是对于未受精和受精卵进行离体翻译的双相电泳分析结果说明虽然两者的mRNA在含量和种类上都是相同的,但某些蛋白质是受精前的卵所特有的,而另一些蛋白质则是受精后特有的。
翻译后控制 翻译后控制的事例不多。一般认为脑垂体后叶细胞产生的促肾上腺皮质激素和脂肪酸释放激素是由同一原始翻译产物经不同的加工而形成的。迄今为止对于真核生物基因调控作用的了解仍然处在探索的阶段,特别是对于高等动植物的基因调控过程了解得更少,还不能形成一个完整的模式。1972年美国学者E.戴维森和R.J.布里顿在实验事实还不充分的情况下提出了一个真核生物的基因调控模型,这一模型也可以用来解释真核生物中大量的脱氧核糖核酸重复顺序的功用。
按照这一模型,外来的信号物质和感应蛋白结合后作用于感应基因,于是综合基因组转录产生激活蛋白的mRNA,并进一步合成激活蛋白,这些激活蛋白又作用于结构基因前面的接受顺序,于是结构基因转录而产生一系列的酶或其他蛋白质。
在这模型中假定感应基因便是一些重复顺序,又假定各个结构基因分布在染色体的不同位置上。这一模型企图解释重复顺序的功能以及分布在不同位置上的基因怎样能被协调控制。
曾经发现在真核生物的细胞核中存在着大量与结构基因无关的 RNA,这一事实是和这一模型符合的。这一模型还是一个有待充分验证的假说。这一模型提出以后,又出现了一些修改这一模型的假说,它们都可以作为进一步研究的出发点。
实用意义
细菌通过基因调控可以避免合成过量的氨基酸、核苷酸等物质。人们要利用细菌来生产这些物质,就必须使它们丧失有关的基因调控作用。在一般的野生型细菌中,阻遏蛋白和氨基酸等代谢最终产物结合后便作用于操纵基因而使转录停止。有两类突变型可以使细菌处于消阻遏状态而合成过量的氨基酸等物质。一类是操纵基因突变型,由于操纵基因的结构改变,使阻遏蛋白不能和它结合,因而操纵子便经常处于活动状态;另一类是调节基因突变型,它编码一种不能和代谢最终产物相结合的阻遏蛋白,因而阻遏蛋白不再作用于操纵基因而同样可使操纵子经常处于活动状态。氨基酸的结构类似物(例如5-甲基色氨酸是色氨酸的类似物)有抑制细菌生长的作用。许多抗类似物突变型属于上述两类突变型,在这些突变型细菌中与合成这一氨基酸直接有关的酶的量增加了,这一氨基酸也大量地被合成。目前在氨基酸、核苷酸的发酵生产中所应用的菌种多数是这些突变型。