再生(动物)(卷名:生物学)
regeneration (animal)
━━━━━━━━━━━━━━━━━━━━━━━━
各类动物的再生能力
原生动物
海绵动物
腔肠动物
扁形动物
线形动物
纽形动物
环节动物
软体动物
节肢动物
棘皮动物
脊索动物
脊椎动物
两栖类肢体再生过程
伤口愈合
组织去分化
再生芽基形成
再生体的形态发生
影响再生过程的因素
损伤的影响
表皮的影响
神经的影响
激素的影响
再生机制的分析
再生芽基细胞的来源
现生芽基细胞的发育潜能
再生与极性
再生芽基是一个自主分化的体系━━━━━━━━━━━━━━━━━━━━━━━━
生物体在个体发育中已经发育了的器官,因自然损伤或人为切割,丢失后的形态和功能的重建。再生是生物体的基本属性之一。再生可以在机体的不同结构水平上进行。红细胞和各种上皮细胞的不断增殖,毛发、鳞甲的脱落更换以及创伤愈合,是细胞和组织水平的再生;蝾螈幼虫或成体的肢体(腿)或尾被切断后可再生出失去的部分,属于器官水平的再生;水螅或涡虫被横切为两段,可分别再生,成为两个个体,是整体水平的再生。再生现象在位于系统发育不同等级的生物中均普遍存在,只是表现水平有所不同。整体水平的再生和无性生殖很难区分。植物的一片叶或一段茎可以再生出完整的植株;沿长轴把水螅切为两段或更多的片段,每一段都能再生为一个新的水螅。这样的情形也可以看成是无性繁殖。再生是生物在长期进化过程中的一种适应。有些昆虫和蟹类的足或爪被敌害捕获时会通过自切,丢弃足或爪而逃遁,蜥蜴遇敌害追捕时会卸下尾巴,事后都能分别再生出失去的足、爪或尾巴。鹿角脱落则是哺乳类自切能力的表现。动物再生有变形再生和新建再生两种基本类型。
各类动物的再生能力
较低等的动物再生能力比较强,高等动物的整体和器官再生能力明显降低,但仍然保持了组织的再生能力。
原生动物 受到损伤后,只要保留一定的细胞核成分和少量的细胞质就能够再生成一个新个体。如喇叭虫虫体缺损后可用补充修复的方式再生成完整的个体。急游虫再生过程中大核起重要作用。
海绵动物 群体海绵,如浴海绵和针海绵的碎块,在适当的条件下能恢复为完整的海绵。甚至人为地把海绵的小块磨碎并用稀薄的麻布过滤,分离开来的细胞能够重新聚合成小的团块,并各自长成一个完整的小海绵(见细胞的亲和性)。
腔肠动物 如将水螅横切为前后两段或多段,每段都能重建成完整的小水螅并长到原先的大小。甚至仅相当于原来个体的 1/200的小片段也有这种能力。营群体生活的筒螅类,水螅头被切除后也可以再生。不同腔肠动物再生能力也有差异。一般水螅型的再生能力很强,而水母型的再生能力却很弱。
扁形运物 涡虫是再生研究的常用材料。把它横切为两半,再生立即从切面开始:未分化的和去分化的细胞在切面聚集形成再生芽基,芽基经有丝分裂产生新细胞,发育出失去部分的各种组织,前半段重新形成后段;后半段则发育出新的前半段;恢复成两个形态和功能均属完整的新涡虫。其他扁形动物的再生能力却很小。
线形动物 再生能力很弱,被切割后只能封闭损伤表面。有人认为这与其体细胞高度分化和细胞定数有关。
纽形动物 一很小片段可再生成完整的个体。
环节动物 多毛纲如沙蚕或寡毛纲如环毛蚓都有很强的再生能力。但均有一定限制:如切断面前端的再生能力因种类和切割水平不同而异。例如切去一种异唇蚓(Allolobophora foetida)前端4~5节,经再生可以恢复原来的总体节数,如果切除长度超过5体节,仍只能再生出4~5个体节,如果切除面在生殖节之后(第10~14节)也只能再生出前部的4~5节而缺乏生殖节。异唇蚓再生出后段的能力要比再生出前段的能力强。环节动物再生时先在切割面形成再生芽基,然后从芽基发育出失去的部分。环节动物门中的蛭纲内的动物却完全不能再生。
软体动物 再生能力相当差。腹足类的眼和眼柄可以再生,头和足也能部分再生,整个头部则不能再生。如果把脑和神经结连同头的一部分一起切除,就不能再生了。头足类仅能再生失去的腕。
节肢动物 大多数甲壳类,无论幼虫或成体,其附肢失去或被截断后,伤口即由几丁质封闭,并在其下形成一个再生芽,逐渐发育成一个新的附肢。由于有老的几丁质层覆盖,新生的比较小的附肢要经过一次蜕皮才能显露,经过几次蜕皮才能长到原来的大小。
棘皮动物 除海胆纲或许因为有坚硬的外壳,仅能修复管足和体表的覆盖部分及其附属的棘针和棘钳外,其余各纲都有很强的再生能力。并和无性繁殖界限模糊。如果一条海参在身体的近中部分自切,两个断块可各自补足缺乏的前端或后端。海参还可以从肛孔喷出肠道和其他器官而后重新修复。海星和海蛇尾也有类似的情况。海星一条臂脱落后,脱落的臂亦可再生出基盘并再生出所缺失的一切器官。
脊索动物 被囊类(海鞘)普遍具有再生能力。如一种群体海鞘(Perophor viridis)可从匍匐枝上出芽形成新虫体。它只要保留一个很小的皮囊,加上一点围鳃腔或围心腔的上皮即可再生出有功能的完整个体。
脊椎动物 器官再生能力不及无脊椎动物。比较典型的器官再生是蝾螈的腿或水晶体、蛙类蝌蚪的肢体和尾巴的再生等。大多数脊椎动物的器官再生能力很有限。鱼类仅能再生出缺损的鳍而不能再生尾。蜥蝎能自切并再生新尾,但再生的尾椎骨数目和形态均不正常,其外表覆盖的鳞片也和正常的不同。鸟类仅再生羽毛能力很强,有些鸟能定期脱换喙,如雄海鸭。哺乳动物再生能力限于组织水平。骨骼、肌肉或皮肤及其衍生物都有很强的再生能力。肝在特定情况下也能大幅度代偿性增生。
两栖类肢体再生过程
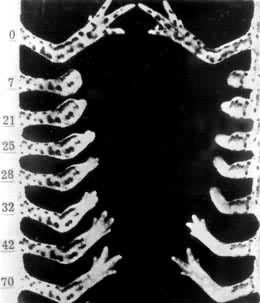
成年蝾螈的再生速度比幼体慢得多,但肢体被切割后,残存的部分都能再生并恢复到原先的大小(图1)。蝾螈肢体再生的组织学过程大体可分以下几个阶段(图2):
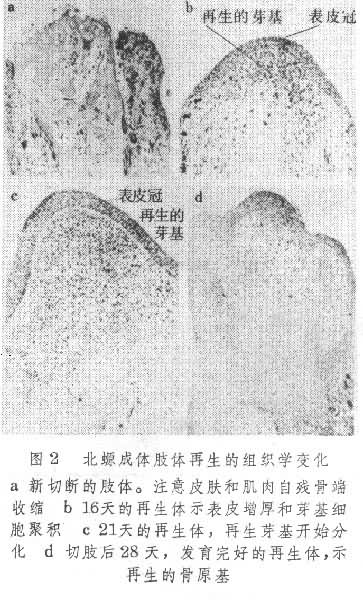
伤口愈合 切肢后,损伤的细胞碎屑和凝结的血块构成伤口的临时覆盖,伤口边缘的表皮细胞借变形运动迅速向伤区迁移,覆盖裸露的伤口内部组织,这一过程一般在1至2天内完成。继而表皮细胞增生,形成加厚的表皮冠,它们失去原先的分化特征而呈现类似胚胎肢芽表皮的外貌。
组织去分化 伤区被表皮细胞覆盖后,此过程几乎立即开始。邻近伤区残肢内部组织如肌肉、软骨(骨)以及结缔组织去分化形成一个个彼此极为相象的间叶状细胞,从形态上很难区分它们原先来自何种组织。
再生芽基形成 由残肢组织去分化而形成的间叶状细胞,可能受到表皮冠的影响聚集在它下面。间叶状芽基细胞行有丝分裂以增加细胞数量,并不断向再生肢顶部集中。由于表皮尚未形成新的基膜而得以和内部的芽基细胞密切接触。随着芽基细胞向再生肢顶端集中,增厚的表皮冠相应地变薄,使残肢末端呈芽状,故称之为再生芽基。此时内部血流恢复通畅。再生芽基形成后残肢去分化随即终止。
再生体的形态发生 再生体的组织分化与胚胎肢芽的分化过程类似,区别在于前者是在同已有的骨骼和肌肉保持接触的基础上,分化出失去的各种组织成分。在芽基形成的较晚阶段,增殖并积聚的间叶状芽基细胞在再生体的中央接近原有的骨末端整齐地排列。这是软骨形成的雏型。继而更远端的软骨成分依次出现,并逐步骨化为硬骨。骨周围的芽基细胞分化为肌肉,和保留下来的肌肉衔接,由近及远地依次发育。血流和神经分布也逐渐恢复到损伤前的水平,最终形成一个有功能的肢体。
影响再生过程的因素
再生过程一方面取决于再生体的结构、组织分化的程度或个体发育的阶段等因素,另一方面也会因环境或特殊的处理而增强、降低或被抑制。再生的速度明显的依赖于温度。例如涡虫在低于 3℃情况下不能再生,在29.7℃的情况下再生很快,而超过31.5℃,再生又缓慢了。
在水螅中发现四种影响再生的物质:促头部因子和抑头部因子,促足部因子和抑足部因子。促头部因子是分子量为1124道尔顿的11肽,其氨基酸排列顺序为:pGlu-Pro-Pro-Gly-Gly-Ser-Lys-Val-Ile-Leu-Phe。促足部因子是分子量介于500~1000 的多肽。两种抑制因子的分子量均小于500,化学性质还不清楚。这些物质的作用是专一的,头部因子只影响头的形成,足部因子只影响足的形成。它们沿水螅体轴呈梯度分布,头部因子的浓度在头端最高,足部因子在足端最高,由此决定再生的极性。头部因子的释放是诱导头部再生的必要条件,而足部因子的释放则是诱导足部再生的必要条件。
损伤的影响 再生的发动需要一定的条件,并受整体因素或组织之间相互作用的调节。损伤如切割或结扎等通常是发动再生的最基本条件。
表皮的影响 表皮对两栖类再生的发动有重要促进作用,真皮则有阻碍作用。如果被切断的肢体伤面没有表皮覆盖,就不能形成再生芽基。如果在切割后立即给伤面覆盖一块皮(包括真皮)则阻止再生。但是如果在切割后1~2天,当伤区表皮已经完全形成了,再缝上一块皮(包括真皮)就不再阻止再生。成体蛙的肢体不能再生是因为切割后伤区愈合是由全部皮肤(包括真皮和表皮)迁移并覆盖伤面。可以再生的蝌蚪,伤区愈合则只靠表皮层迁移完成。如果把蝌蚪的幼虫皮移到蛙的残肢上代替成体皮覆盖创面,则可以使已失去再生能力的蛙恢复部分的再生能力。
研究伤区表皮的超微结构表明,由伤区周围迁移来覆盖伤面的表皮细胞呈现明显的去分化特征。这些细胞由于和内部组织之间没有基膜相隔得以直接接触。
有证据表明伤区表皮的作用可能是促进邻近组织的去分化。另一个重要的作用是形成加厚的表皮冠,吸引去分化的芽基细胞在其下聚积,形成再生芽基。不能再生肢体的成蛙或更高等的脊椎动物在伤面处均不能形成表皮冠,它们的间叶状细胞仅参与局部的组织修复而不聚积成芽基。
神经的影响 再生是成体的形态发生过程,除受切断面局部组织相互作用影响外,还受整体的神经、内分泌因素控制。很早就发现切断肢体的神经供应能有效地抑制再生。如把成体蝾螈的腿神经或上臂神经在切肢的同时或在再生早期破坏,再生便受到抑制,或使再生芽基停止生长。蝾螈幼体切肢前切断其神经,残肢去分化过程不受限制地进行结果芽基不能形成,整个残肢终被破坏、吸收。在再生不同时期切断神经的实验证明再生对神经的依赖在于早期的去分化过程。当再生芽基形成并达到一定体积后,神经对于再生体的形态发生就不再必要了。
S.J.辛格曾提出神经阈值学说,认为再生能力与神经供应的数量有直接关系。当肢体的神经纤维的数量减少到小于正常数量的1/3至1/2时,再生便不能进行。反之增加或补充神经供应则可以发动或重新唤起再生。例如将蝾螈腿神经末端移位到腿的基部可以在该处诱发一个再生芽样的腿原基并可发育出一条完整的腿;补充伤区神经供应可以唤起已失去再生能力的成年蛙部分恢复前肢再生;在蜥蜴和新生的负鼠上的实验也得到类似的结果。对神经影响再生的作用的本质曾设想是神经通过释放某种化学物质──神经营养因子来刺激再生的。
对于两栖类肢体再生,历来认为神经是有决定意义的。然而,胚胎期肢芽的发育并不依赖神经。于是,很自然地会提出这样的问题:肢体再生对神经的依赖关系是不是在发育过程中产生的呢?中国胚胎学家庄孝僡、王亚辉1956年发现东方蝾螈神经胚胎时期割除神经板后得到的无神经幼虫,切断前肢后,仍能再生出形态完善的肢体。这一结果提示肢体再生对神经的依赖可能是在个体发育过程中随神经联系的建立而出现的一种次生现象。正常或无神经幼虫肢体的再生是否依赖神经,取决于是否曾建立过这种联系。也就是说再生能不能进行的根本原因在于肢体组织的特性,神经也是通过对组织及其相互关系的影响而起作用的。
激素的影响 1926年O.E.肖泰发现成体冠蝾螈在切除垂体的同时切断肢体,则不能再生。如果在截肢后间隔若干天再切除垂体,再生的程度依推迟时间的长短而定,切肢13天后再切除垂体将不影响再生。这说明垂体激素仅影响再生的初期阶段。给切除垂体动物植入肾上腺或注射ACTH或肾上腺皮质激素补偿均能使这样的动物恢复再生能力。反之,使用抑制肾上腺皮质激素的药物处理则能抑制再生。 这表明垂体-肾上腺协同机制的确在肢体再生中起作用。
生长素和促乳素直接影响细胞代谢,因而也可能影响芽基形成和生长。促肾上腺素和促甲状腺素可引起靶组织专一性的改变而导至再生能力的变化。无尾类蝌蚪一经变态就失去了再生能力以及成体蝾螈的再生明显的比幼虫缓慢等事实均提示激素变化可能影响再生能力。
再生机制的分析
再生芽基细胞的来源 不同类型动物中再生芽基细胞来源不同。低等动物再生芽基细胞既可来自局部组织因去分化形成的间叶状细胞,也可以来自离切割面很远的部分,如分布全身的贮备细胞。涡虫经X射线照射后即失去再生能力。如果只照射其前半段,再通过照射区切割,结果切下来的全部被照射过的前段既不能再生,也不能存活;而带有未照射过的组织的后段残片,虽然再生速度有所延缓却能在照射过的切割面上形成再生芽,并分化出完整的头。这说明再生芽基细胞可来自离切割面很远的身体部分。
高等动物(如有尾两栖类)再生芽基的形成是由于残存组织──特别是接近切割面的内部组织──去分化形成间叶状芽基细胞在伤区表皮下的聚积。全部中胚层组织都能以这种方式为芽基提供细胞。中胚层间叶细胞,神经鞘细胞也可参与芽基形成。表皮细胞则既不进入芽基也不能转变为内部的中胚层细胞。用 X射线局部照射蝾螈肢体,然后分别通过照射区或非照射区切割肢体,结果通过照射区切割的不能再生,通过被铅板遮盖的未照射区切割的可以再生出完好肢体(图3)。这表明再生物质来源于伤区局部。

再生芽基细胞的发育潜能 去分化的、在外表上极其相似的芽基细胞在发育潜能方面是否也恢复到胚胎未分化细胞的多潜能状态,从而能分化出各种组织,或是来自不同组织的去分化芽基细胞仍保持着各自的组织专一性,在再生过程中仍分化为原先的组织这仍然是一个悬而未决的问题。
在低等动物,贮备细胞或成体未分化细胞显然是多潜能的,另一方面已分化细胞转变分化方向也是可能的。如果在中部把涡虫横切为两段,在各自的切割面上积聚的芽基细胞,一部分来自局部组织去分化,一部分来自贮备细胞。来自局部组织的去分化细胞分化命运可以相当不同。位于前半段末端的分化为身体后半段组织和器官,位于后半段前端的则分化出身体的前半段组织和器官。这说明芽基细胞有多种发育潜能,残存的已分化组织或器官对芽基的分化有明显的调节作用。
高等动物如蝾螈的前肢在切肢之前或与之同时摘除肱骨,能再生出切面以下的全部骨骼。 将未经X射线照射的后肢肌肉或尾部肌肉移置到经过照射的摘除肱骨的前肢内,结果也有少数可再生出骨骼。将标记的肌肉植入去肱骨的蝾螈幼虫腿中也得到类似的结果。这说明非骨组织能够提供芽基细胞,再分化出软骨和骨膜。很可能在肌肉组织中存在着多潜能的细胞。这些实验结果似乎说明再生中组织之间可以转变。另一方面,也有实验指出骨组织在再生中是相对恒定的,不能去分化为多潜能的细胞。如将三倍体的软骨移植到两倍体美西螈,发现三倍体细胞只出现在再生肢的软骨组织中。所以芽基细胞的分化潜能还是一个尚待阐明的问题。
再生与极性 生物体及其器官都存在形态和生理上的极性。如一条蚯蚓分前端、后端,两端各有不同的生理特性;蝾螈的肢体可以区分出前-后轴,背-腹轴和远心-近心轴等。再生的部分通常都能维持原有的极性,保持机体的统一。可以用各种实验方法使极性转变以探索再生与极性的关系。筒水螅的口极氧耗最高,沿身体主轴向下氧耗量逐渐降低。涡虫自前而后存在着蛋白质合成速率递减的梯度说明生物体中存在着生理代谢的极性。这种代谢的极性和形态发生密切相关。如水螅被横切成若干段后,原来向着口极的一端再生出新的水螅头,相反的一端再生出基盘。如果把向着口极的一端结扎起来或插入沙中,则可在向着基盘的一端再生出新的水螅头。用代谢毒物处理涡虫降低其前端代谢优势,结果原来的后端(尾)被切割后可再生一个新头成为双头涡虫。
蝾螈的肢体也呈现明显的极性。根据肢体的空间构型可以区分出 3个轴,远近轴、背腹轴和前后轴。远近轴可像水螅一样改变,但前后轴、背腹轴则相当恒定。在整个再生过程中维持不变。如果将再生芽基的背腹或前后轴转动使与残肢的背腹或前后轴呈一定的角度,则可导致将来再生出的肢体的畸形。肢体的外部组织和内部组织也有极性,如果将肢体的皮肤转动一定的角度使与其下的内部组织极性不一致也可以导致再生体畸形,如出现倍生肢、多指或掌心掌背极性反转以及缺指或再生障碍等。这表明残肢各种组织之间相互作用并最终达到协调是再生正常进行的前提。
再生的形态发生机制还有待研究。已有一些假说如1976年S.V.布赖恩特提出钟面模型假说,主张处于肢体断面不同部位的细胞具有不同的形态发生位值。在正常再生过程中,这些具有不同位值的细胞协调地相互作用保证再生体的形态发生能正常进行。如果将再生芽基的轴转动一定角度,造成细胞之间位值不相匹配,就会导致再生异常或抑制。
再生芽基是一个自主分化的体系 蝾螈肢体的再生芽基十分类似发育中的肢芽,不同的是肢芽发育成完整的肢体,而再生芽基仅再生出失去的部分。早期再生芽基具有胚胎器官原基的性质,在原基范围内各部分之间显示相当的调整能力(如将年轻的芽基切为两半,可各自发育为形态完整的再生体),各部分的发育命运只是在再生进行中受到残肢组织的影响才逐渐决定的。但是,再生芽基作为一个整体而言,一开始就是一个按其来源能进行自主分化的发育系统,不因移植到新的环境而改变其发育方向。如将尾的早期再生芽基移植到切割后的残肢断面则发育出肢-尾混合型再生体。如果用X射线损伤形成肢体芽基的细胞,移植的尾芽基只能发育成尾。这说明尾再生芽基可以自主分化,残肢只能以其细胞插入生长的方式影响芽基。用异位移植和外植的方法培养圆锥期及更晚的肢体再生芽基能够分化出前软骨和横纹肌,说明再生芽基达到圆锥期时它的分化就不再依赖残肢的诱导而已成为一个自主分化的体系。1982年在美西螈进行的再生芽基和发育中芽基互换的实验证明再生肢和发育中的肢体模式形成的控制机制是一样的。1982年罗尔曼 -丁斯莫尔等把蝾螈前肢和后肢再生芽基互换的实验结果表明:如属同侧互换,移植的再生芽基的轴性和残肢的背-腹,前-后轴保持一致,结果芽基将按其来源发育为完整的前肢或后肢。如属异侧互换,移植的再生芽基的轴性和残肢的背腹、前后轴不协调一致,结果除按芽基来源再生出前肢或后肢外,还可出现多肢或多指的再生体。
脊椎动物的肢体发育的模式调节机制可能是相同的。然而,令人费解的是为什么各种动物肢体再生能力不同,只有很少种类的肢体能够再生。即便在同一种类,为什么在一个发育时期(如蛙的蝌蚪),肢体能够再生,而另一发育时期(变态后的小蛙)就不能再生。在系统发育和个体发育过程中,肢体再生能力丧失的原因何在,能否用实验方法使之恢复,仍是一个引人瞩目而尚未解决的问题。