conservation
ecology
Introduction
study of the loss of Earth's biological diversity and the ways this loss can be prevented. Biological diversity, or biodiversity, is the variety of life either in a particular place or on the entire Earth, including its ecosystems, species, populations, and genes. Conservation thus seeks to protect life's variety at all levels of biological organization.
 Species extinction is the most obvious aspect of the loss of biodiversity. For example, species form the bulk of the examples in a comprehensive assessment of the state of the planet published in the early 21st century by the Millennium Ecosystem Assessment, an international effort coordinated by the United Nations Environment Programme. The subject of conservation is broader than this, however. Even a species that survives extinction can lose much of its genetic diversity as local, genetically distinct populations are lost from most of the species' original range. Furthermore, ecosystems may shrink dramatically in area and lose many of their functions, even if their constituent species manage to survive. Conservation is involved with studying all these kinds of losses, understanding the factors responsible for them, developing techniques to prevent losses, and, whenever possible, restoring biodiversity.
Species extinction is the most obvious aspect of the loss of biodiversity. For example, species form the bulk of the examples in a comprehensive assessment of the state of the planet published in the early 21st century by the Millennium Ecosystem Assessment, an international effort coordinated by the United Nations Environment Programme. The subject of conservation is broader than this, however. Even a species that survives extinction can lose much of its genetic diversity as local, genetically distinct populations are lost from most of the species' original range. Furthermore, ecosystems may shrink dramatically in area and lose many of their functions, even if their constituent species manage to survive. Conservation is involved with studying all these kinds of losses, understanding the factors responsible for them, developing techniques to prevent losses, and, whenever possible, restoring biodiversity. Conservation is a crisis discipline, one demanded by the unusual rates of loss; it is also a mission-driven one. By analogy, ecology and conservation have the same relationship as physiology and medicine. Human physiology studies the workings of the human body, whereas medicine is mission-oriented and aims to understand what goes wrong and how to treat it. The major parts of this article thus deal first with the “pathology” of extinction—why and how biodiversity is lost—and second with the “treatment” methods to prevent these losses.
Conservation is a crisis discipline, one demanded by the unusual rates of loss; it is also a mission-driven one. By analogy, ecology and conservation have the same relationship as physiology and medicine. Human physiology studies the workings of the human body, whereas medicine is mission-oriented and aims to understand what goes wrong and how to treat it. The major parts of this article thus deal first with the “pathology” of extinction—why and how biodiversity is lost—and second with the “treatment” methods to prevent these losses.Conservation is often considered a purely biological topic, as exemplified by major scientific journals with titles such as Conservation Biology and Animal Conservation as well as college textbooks with such titles as Principles of Conservation Biology and Essentials of Conservation Biology. However, because the underlying cause of the loss of biodiversity is increasing human activity, conservation must inevitably involve human interactions. Many of the techniques to prevent the loss of biodiversity involve issues of economics, law, social sciences, and religion—all of which are covered by the journals and textbooks cited above.
The “pathology” section of this article begins by documenting the losses of species. In doing so, it shows that a set of common factors are responsible; these are then individually identified and discussed. The final part of the section demonstrates that some species and ecosystems are much more likely to lose biodiversity than others. The other main division, the “treatment” section, considers a variety of “therapies” that address the problems identified in the first section.
The pathology of extinction
Rates of natural and present-day species extinction
Estimates of recent and future extinction rates Estimates of recent and future extinction ratesAccording to the best estimates of the world's environmental experts, human activities have driven species to extinction at rates perhaps 1,000 times the natural, or background, rate, and future rates of extinction will likely be higher. To show how the experts arrived at these conclusions, it is necessary to pose and attempt to answer a series of extremely difficult questions. How many species are there? How fast were species disappearing before human activity became pervasive? How fast are they becoming extinct at present? And finally, it is necessary to ask a further question: What does the future hold for extinctions if current trends continue?
How many species are there?
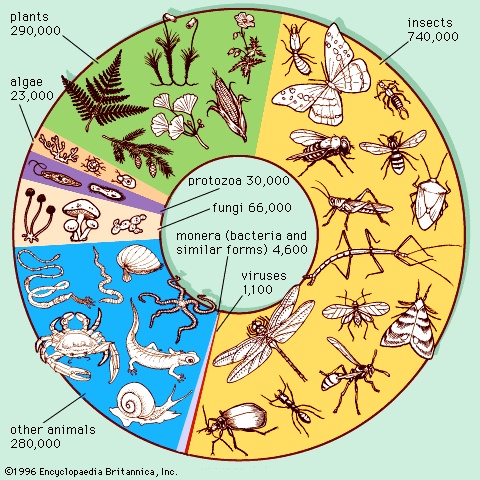 Any absolute estimate of extinction rate, such as extinctions per year, requires knowledge of how many species there are. Unfortunately, this number is not known with any great degree of certainty, and the problems of estimating it are formidable. Taxonomists have described—that is, have given names to—about 1.5 million species. Only about 100,000 of them, comprising terrestrial vertebrates, some flowering plants, and attractive and collectible invertebrates such as butterflies and snails, are popular enough for taxonomists to know well. Birds are exceptionally well-known—there are roughly 10,000 bird species, with only one or two new species being added each year.
Any absolute estimate of extinction rate, such as extinctions per year, requires knowledge of how many species there are. Unfortunately, this number is not known with any great degree of certainty, and the problems of estimating it are formidable. Taxonomists have described—that is, have given names to—about 1.5 million species. Only about 100,000 of them, comprising terrestrial vertebrates, some flowering plants, and attractive and collectible invertebrates such as butterflies and snails, are popular enough for taxonomists to know well. Birds are exceptionally well-known—there are roughly 10,000 bird species, with only one or two new species being added each year.Those who describe species cannot always be certain that the specimen in hand has not been given a name by someone else in a different country and sometimes even in a different century. Consequently, some taxonomic groups may have more names assigned to them than constituent species, which would result in erroneously high species estimates. Potentially much more serious as a source of error is the fact that some species groups have relatively few named members compared with the numbers that experts think exist in those groups. For example, taxonomists have only sparsely sampled some potentially rich communities, such as the bottom of the deep ocean and the canopies of rainforests.

 One estimate of how many species might still be undescribed involves a comparison of fungi (fungus) and flowering plants (angiosperms (angiosperm)). In Great Britain, where both groups are well-known, there are six times as many named species of fungi as of flowering plants. If this ratio applies worldwide, the world total of about 300,000 species of flowering plants, which are fairly well-known globally, predicts a total of about 1.8 million species of fungi, which are not. Because only about 70,000 species of fungi currently have names, the prediction would suggest that these named species constitute only 1/25of the species that exist. On the other hand, samples from poorly known parts of the world suggest that the named species constitute a much larger fraction. Although this estimate of the number of species of fungi is almost certainly too high, it nonetheless suggests that there are large numbers of unknown species.
One estimate of how many species might still be undescribed involves a comparison of fungi (fungus) and flowering plants (angiosperms (angiosperm)). In Great Britain, where both groups are well-known, there are six times as many named species of fungi as of flowering plants. If this ratio applies worldwide, the world total of about 300,000 species of flowering plants, which are fairly well-known globally, predicts a total of about 1.8 million species of fungi, which are not. Because only about 70,000 species of fungi currently have names, the prediction would suggest that these named species constitute only 1/25of the species that exist. On the other hand, samples from poorly known parts of the world suggest that the named species constitute a much larger fraction. Although this estimate of the number of species of fungi is almost certainly too high, it nonetheless suggests that there are large numbers of unknown species.For insects, there are about 1 million described species, yet estimates of how many insect species exist range from 10 to 100 times this number. As an example of how part of one such estimate was made, an entomologist collected a large sample of canopy-dwelling beetles from one species of tropical tree. The tree species had 163 beetle species specific to it. There are about 50,000 tree species in the world's tropical forest (tropical rainforest)s, so simple multiplication predicts roughly 8 million species of tropical forest-canopy beetles. Since 40 percent of described insects are beetles, the total number of tropical forest-canopy insects could be 20 million. Adding half that number for insect species found on the ground beneath the trees gives an estimated grand total of 30 million species of tropical forest insects alone.
This example is critically dependent on the first number—the 163 species found only on the one species of tree. In addition, other calculations suggest that this estimate is too high. The number of insect species in tropical forests is more likely to be between 7 and 15 million, which, however, still means that most of Earth's species would be tropical forest insects. Given the considerable uncertainties, many scientists consider that the total of all species is very roughly 10 million, a number that likely translates to “somewhere between 5 and 20 million with arguments for numbers both lower and higher.”
An obvious concern follows regarding the usefulness of such calculations as a basis for assessing the loss of species. Any absolute estimate of species extinctions must be extrapolated from the 100,000 well-known species of living plants and animals, to the roughly 1.5 million described species, to the likely grand total of very roughly 10 million. Because of uncertainties about the total number of living species, published statements regarding the total number of species that become extinct per year or per day can vary a hundredfold.
Another approach to assessing species loss is to derive relative estimates—estimates of the proportion of well-known species that become extinct in a given interval. Estimating such proportions is the basis for the remainder of the discussion on rates of extinction, but it raises a critical concern of its own—namely, are these proportions actually typical of the great majority of species that are still undescribed? They are likely to be so if extinction rates in widely different species groups and regions turn out to be broadly similar.
There also is another way in which estimates of extinctions can be made relative. Extinctions have always been a part of Earth's history. It is possible to make any estimates of massive future extinction relative to that history.
Calculating background extinction rates
 Average species life spans and extinction rates for various groups of fossil marine speciesTo discern the effect of modern human activity on the loss of species requires determining how fast species disappeared in the absence of that activity. Studies of marine fossils show that species last about 1–10 million years (see the table (Average species life spans and extinction rates for various groups of fossil marine species), second column). Assume that all these extinctions happened independently and gradually—i.e., the “normal” way—rather than catastrophically, as they did at the end of the Cretaceous Period about 65 million years ago, when dinosaurs and many other land and marine animal species disappeared. On that basis, if one followed the fates of one million species, one would expect to observe about 0.1–1 extinction per year—in other words, one species going extinct every 1–10 years.
Average species life spans and extinction rates for various groups of fossil marine speciesTo discern the effect of modern human activity on the loss of species requires determining how fast species disappeared in the absence of that activity. Studies of marine fossils show that species last about 1–10 million years (see the table (Average species life spans and extinction rates for various groups of fossil marine species), second column). Assume that all these extinctions happened independently and gradually—i.e., the “normal” way—rather than catastrophically, as they did at the end of the Cretaceous Period about 65 million years ago, when dinosaurs and many other land and marine animal species disappeared. On that basis, if one followed the fates of one million species, one would expect to observe about 0.1–1 extinction per year—in other words, one species going extinct every 1–10 years.Human life spans provide a useful analogy to the foregoing. If humans live for about 80 years on average, then 1 in 80 individuals should die each year under normal circumstances. If, however, many more than 1 in 80 were dying each year, then something would be abnormal. There might be an epidemic, for instance.
To make comparisons of present-day extinction rates conservative, assume that the normal rate is just one extinction per million species per year. This then is the benchmark—the background rate against which one can compare modern rates. For example, given a sample of 10,000 living described species (roughly the number of modern bird species), one should see one extinction every 100 years. Comparing this to the actual number of extinctions within the past century provides a measure of relative extinction rates.
The estimates of the background extinction rate described above derive from the abundant and widespread species that dominate the fossil record. By contrast, as the article later demonstrates, the species most likely to become extinct today are rare and local. Thus, the fossil data might underestimate background extinction rates. Importantly, however, these estimates can be supplemented from knowledge of speciation rates—the rates that new species come into being—of those species that often are rare and local. These rates cannot be much less than the extinction rates, or there would be no species left.
To explore the idea of speciation rates, one can refer again to the analogy of human life spans and ask: How old are my living siblings? The answer might be anything from that of a newborn to that of a retiree living out his or her last days. The average age will be midway between them—that is, about half a lifetime. Ask the same question for a mouse, and the answer will be a few months; of long-living trees such as redwoods, perhaps a millennium or more. The age of one's siblings is a clue to how long one will live.
 Species have the equivalent of siblings. They are the species' closest living relatives in the evolutionary tree (see evolution: Evolutionary trees (evolution))—something that can be determined by differences in the DNA. The closest relative of human beings is the bonobo (Pan paniscus), whereas the closest relative of the bonobo is the chimpanzee (P. troglodytes). Taxonomists call such related species sister taxa, following the analogy that they are splits from their “parent” species.
Species have the equivalent of siblings. They are the species' closest living relatives in the evolutionary tree (see evolution: Evolutionary trees (evolution))—something that can be determined by differences in the DNA. The closest relative of human beings is the bonobo (Pan paniscus), whereas the closest relative of the bonobo is the chimpanzee (P. troglodytes). Taxonomists call such related species sister taxa, following the analogy that they are splits from their “parent” species. The greater the differences between the DNA of two living species, the more ancient the split from their common ancestor. Studies show that these accumulated differences result from changes whose rates are, in a certain fashion, fairly constant—hence, the concept of the molecular clock (see evolution: The molecular clock of evolution (evolution))—which allows scientists to estimate the time of the split from knowledge of the DNA differences. For example, from a comparison of their DNA, the bonobo and the chimpanzee appear to have split one million years ago, and humans split from the line containing the bonobo and chimpanzee about six million years ago.
The greater the differences between the DNA of two living species, the more ancient the split from their common ancestor. Studies show that these accumulated differences result from changes whose rates are, in a certain fashion, fairly constant—hence, the concept of the molecular clock (see evolution: The molecular clock of evolution (evolution))—which allows scientists to estimate the time of the split from knowledge of the DNA differences. For example, from a comparison of their DNA, the bonobo and the chimpanzee appear to have split one million years ago, and humans split from the line containing the bonobo and chimpanzee about six million years ago.The advantage of using the molecular clock to determine speciation rates is that it works well for all species, whether common or rare. It works for birds and, in the previous example, for forest-living apes, for which very few fossils have been recovered. In the preceding example, the bonobo and chimpanzee split a million years ago, suggesting such species' life spans are, like those of the abundant and widespread marine species discussed above, on million-year timescales, at least in the absence of modern human actions that threaten them. This is just one example, however. Is there evidence that speciation can be much more rapid?
Until recently, there seemed to be an obvious example of a high rate of speciation—a “baby boom” of bird species. Its existence allowed for the possibility that the high rates of bird extinction that are observed today might be just a natural pruning of this evolutionary exuberance.

 On either side of North America's Great Plains are 35 pairs of sister taxa including western and eastern bluebirds (Sialia mexicana and S. sialis), red-shafted and yellow-shafted flickers (both considered subspecies of Colaptes auratus), and ruby-throated and black-chinned hummingbirds (Archilochus colubris and A. alexandri). According to the rapid-speciation interpretation, a single mechanism seemed to have created them all. Each pair of sister taxa had one parent species ranging across the continent. Then a major advance in glaciation during the latter part of the Pleistocene Epoch, some tens of thousands of years ago, split each population of parent species into two groups. Each pair of isolated groups evolved to become two sister taxa, one in the west and the other in the east. Finally, the ice retreated, and, as the continent became warm enough, about 10,000 years ago, the sister taxa expanded their ranges and, in some cases, met once again. (For additional discussion of this speciation mechanism, see evolution: Geographic speciation (evolution).)
On either side of North America's Great Plains are 35 pairs of sister taxa including western and eastern bluebirds (Sialia mexicana and S. sialis), red-shafted and yellow-shafted flickers (both considered subspecies of Colaptes auratus), and ruby-throated and black-chinned hummingbirds (Archilochus colubris and A. alexandri). According to the rapid-speciation interpretation, a single mechanism seemed to have created them all. Each pair of sister taxa had one parent species ranging across the continent. Then a major advance in glaciation during the latter part of the Pleistocene Epoch, some tens of thousands of years ago, split each population of parent species into two groups. Each pair of isolated groups evolved to become two sister taxa, one in the west and the other in the east. Finally, the ice retreated, and, as the continent became warm enough, about 10,000 years ago, the sister taxa expanded their ranges and, in some cases, met once again. (For additional discussion of this speciation mechanism, see evolution: Geographic speciation (evolution).)The story, while compelling, is now known to be wrong. Molecular data show that, on average, the sister taxa split 2.45 million years ago. This means that the average species life span for these taxa is not only very much older than the rapid-speciation explanation for them requires but is also considerably older than the one-million-year estimate for the extinction rate suggested above as a conservative benchmark.
Molecular-based studies find that many sister species were created a few million years ago, which suggests that species should last “a few million” years, too. Indeed, they suggest that the background rate of one extinction among a million species per year may be too high. Nevertheless, this rate remains a convenient benchmark against which to compare modern extinctions.
Recent extinction rates
To what extent has modern human activity increased extinction rates above the background rate? This discussion presents five well-known case histories of recent extinctions. From them, some general features can be deduced about recent extinctions that also provide clues to the future.
Pacific Island (Pacific Islands) birds (bird)
Polynesians reached such remote Pacific islands as the Hawaiian Islands, New Zealand, and Easter Island—Earth's last habitable areas for settlement—within the past 2,000 years. Over that period they left unambiguous evidence that their activity caused many species of birds to become extinct. The bones of many species persist into, but not through, archaeological layers that also contain evidence of human presence. No species is known to have disappeared in the longer intervals before first contact. The Polynesian settlers likely ate the large, probably unwary, and often flightless species. They also introduced pigs and rats to islands far too remote to have acquired native land mammals (see introduced species). The rats also would have found the naive birds, their eggs, and their young to be easy pickings, and the pigs would have destroyed the ground cover of the forests. With only Stone Age technology, the settlers may have exterminated as many as 2,000 bird species, some 17 percent of the world total. Locally, they often exterminated all the bird species they encountered.
In the Hawaiian Islands (Hawaii), for example, scientists have described 43 species only from their bones, a number that has increased as new extinct species have been discovered. Because bird bones are fragile and easily destroyed, all the extinct species may never be found. Nevertheless, the number that remain unknown can be calculated.
Suppose that every Hawaiian bird species that survived to be collected by naturalists since the 1800s were also found as bones. In that case, one would say that the bone record is complete. On the other hand, if only half the species that survived to modern times were also known from bones, one would know that the record is half complete. If this second case were true, then, by extension, only half the species that became extinct by modern times should be known from bones. Thus, for every one of the 43 bird species known only from bones, there ought to be another 43 extinct species still unknown. Half turns out to be about right—scientists have estimated 40 unknown species, for a total number of extinctions of 83.
The British explorer James Cook (Cook, James) found the Hawaiian Islands and their Polynesian settlers in 1778. With the peace that followed Great Britain's defeat of France in 1815, Cook's descriptions of whaling opportunities in the region led to increasing contact with Europe and North America. New colonists not only depleted the whales but also introduced cattle and goats to the islands for food. Like pigs, these alien herbivores destroyed native plants and greatly reduced natural habitats. Naturalists of the time described 18 bird species that did not survive this onslaught, so the total count of extinctions rises to 101. This still is an underestimate, because the 19th-century naturalists missed some species. On Molokai, for example, they recorded hearing a rail, but there is no specimen of it.
On the Hawaiian Islands today there remain a dozen species that are so extremely rare that the chances of saving them are poor; some may already be extinct. Another dozen species are rare and have uncertain futures. Of the estimated 136 bird species that lived on the islands before human settlement, only 11 are common enough to suggest that their future is secure.
If such estimates are expanded across the eastern Pacific, it will appear that the Polynesians exterminated 500–1,000 species of birds. But it is also possible to take a different approach, one that considers what species should occur on every island in the group. Rails (rail), for example, seem very adept at reaching even such remote islands as Henderson and Lisianski. Where scientists have searched for bird bones, they have found fossil rails. It seems likely that every sufficiently large island probably housed a unique species of rail. Most of these islands,but not the Hawaiian Islands, which were likely too remote, also housed unique species of doves and parrots, most of which are now extinct. Even before small-bodied species are counted, such studies suggest that Polynesians exterminated at least 1,000 species of birds. Some scientists put the number at 2,000 or even more.
The Polynesian and European colonizations of the Pacific exterminated species other than birds, of course. The Hawaiian Islands have 980 native plants. Of these, 84 are now extinct, and another 133 have wild populations that number fewer than 100 individuals each. It may never be known how many other species were exterminated as the Polynesians cleared land for their crops and likely burned dry forests.
In the 20th century, misguided individuals on a number of the Pacific islands introduced an African land snail, Achatina fulica, for food. It became a pest. So, like the song about the old woman who swallowed a fly, and then a spider to catch it, and so forth, a predatory snail, Euglandina rosea, was released to control the Achatina. The predatory snail preferred native Achatinella and Partula snails instead, driving many species to extinction.
 There is nothing special about the islands of the Pacific that makes their animals and plants prone to extinction. The most famous recent extinction of all, the extermination of the dodo (Raphus cucullatus) by humans and their introduced animals, was one of 33 species of birds, 30 species of land snails, and 11 species of reptiles lost from Mauritius, Rodrigues, and Réunion in the Indian Ocean in the past three centuries. Over the same period, St. Helena and Madeira in the Atlantic Ocean lost 36 species of land snails as a result of human activity. These island extinctions raise an obvious question: Are extinctions predominantly on islands and not continents? Although the disappearance of island species does demonstrate a pattern of sensitivity to becoming extinct, the next example shows the answer to the question above to be “no.”
There is nothing special about the islands of the Pacific that makes their animals and plants prone to extinction. The most famous recent extinction of all, the extermination of the dodo (Raphus cucullatus) by humans and their introduced animals, was one of 33 species of birds, 30 species of land snails, and 11 species of reptiles lost from Mauritius, Rodrigues, and Réunion in the Indian Ocean in the past three centuries. Over the same period, St. Helena and Madeira in the Atlantic Ocean lost 36 species of land snails as a result of human activity. These island extinctions raise an obvious question: Are extinctions predominantly on islands and not continents? Although the disappearance of island species does demonstrate a pattern of sensitivity to becoming extinct, the next example shows the answer to the question above to be “no.”Flowering plants (angiosperm)
 A small area at the southern tip of Africa houses an extraordinarily rich and unusual variety of plants. It has several types of vegetation, of which the fynbos (a species-rich scrubland of southwestern South Africa) contributes the most species. Of roughly 8,500 species, 36 have become extinct in the past century, and another 618 species are threatened. (In the field of conservation, the term threatened has a specific, technical meaning. It comes from the World Conservation Union's (International Union for Conservation of Nature) 【IUCN's】 Red Lists, the lists of species that are at risk of extinction. A species listed as “threatened” has a high probability of extinction in the wild within the next few decades if nothing is done to prevent it.) For the fynbos, invading alien plants—particularly various species of Australian wattle trees ( acacia)—and the conversion of natural areas to agriculture (agriculture, origins of) are the two major causes of species extinction and endangerment.
A small area at the southern tip of Africa houses an extraordinarily rich and unusual variety of plants. It has several types of vegetation, of which the fynbos (a species-rich scrubland of southwestern South Africa) contributes the most species. Of roughly 8,500 species, 36 have become extinct in the past century, and another 618 species are threatened. (In the field of conservation, the term threatened has a specific, technical meaning. It comes from the World Conservation Union's (International Union for Conservation of Nature) 【IUCN's】 Red Lists, the lists of species that are at risk of extinction. A species listed as “threatened” has a high probability of extinction in the wild within the next few decades if nothing is done to prevent it.) For the fynbos, invading alien plants—particularly various species of Australian wattle trees ( acacia)—and the conversion of natural areas to agriculture (agriculture, origins of) are the two major causes of species extinction and endangerment.Freshwater mussels (mussel) and clams (clam)
Terrestrial ecosystems are far from being the only places where recent extinctions have occurred. The Mississippi and St. Lawrence river basins were home to 297 North American species of the bivalve mollusk families Unionidae and Margaritiferidae. Of these, 21 have become extinct in the past century, and another 120 species are in danger of extinction. During this same period, engineers have extensively dammed and channeled North America's rivers. The Tennessee River, for example, is dammed along its entire length from Knoxville, Tenn., until it joins the Ohio River. Changing flowing rivers to effectively static reservoirs completely changes the habitats of the species that live in them. In addition, poor farming practices have greatly increased nutrient and pesticide runoff from croplands, and sewage effluent and runoff from cities have exacerbated the problem. Freshwater bivalves are sessile filter feeders (see filter feeding) and very sensitive to changes in water quality.
Many of these bivalve species in their juvenile stages parasitize freshwater fish and so are dependent on their survival as well. Some freshwater bivalves are harvested for the cultured pearl trade. And, as if these were not sufficient threats, two introduced species, the Asian clam (Corbicula fluminea) and the zebra mussel (Dreissena polymorpha), have rapidly expanded their North American ranges, outcompeting native species.
Freshwater fish
Some of the changes to North America's rivers that threaten their native bivalves have also seriously harmed the continent's freshwater fish. Of the approximately 950 species of freshwater fish of the United States, Canada, and Mexico, about 40 have become extinct in the past century. They have disappeared from a wide range of habitats—northern lakes, southern streams, wetlands, and particularly springs. Springs are often small, isolated habitats, which favour the evolution of unique species, but those features make the habitats extremely vulnerable to human disturbance. Introduced species—from the stocking of fish and frogs for food and of fish for sport and from accidental aquarium releases—are also factors in 18 of the 40 extinctions. The introduced species can outcompete native fish, sometimes feed on them, and in some cases hybridize with them. Hybridization may seem a surprising cause of extinction. Populations that have evolved in isolation, however, may no longer retain their morphologic and genetic distinctiveness when mixed with similar species from elsewhere.
The southeastern United States holds a little over half of North America's fish, many of which have small geographic ranges, living in one or just a few rivers. Extensive damming and pollution of southern Appalachian mountain streams have exterminated 4 species and threatened another 80.
Australian mammals
 At least 60 mammals (mammal) have become extinct worldwide in the past two centuries, about a third of them from Caribbean islands. Of the remainder, 18 mammals were native to Australia, where they constituted about 6 percent of the terrestrial animal species prior to the British colonization of the continent beginning in the late 18th century. (Aboriginal peoples reached Australia at least 42,000 years earlier and, like the Polynesians across the eastern Pacific, eliminated many native species.) More than 40 other living mammal species in Australia have lost at least half their former ranges, and some survive only on protected offshore islands.
At least 60 mammals (mammal) have become extinct worldwide in the past two centuries, about a third of them from Caribbean islands. Of the remainder, 18 mammals were native to Australia, where they constituted about 6 percent of the terrestrial animal species prior to the British colonization of the continent beginning in the late 18th century. (Aboriginal peoples reached Australia at least 42,000 years earlier and, like the Polynesians across the eastern Pacific, eliminated many native species.) More than 40 other living mammal species in Australia have lost at least half their former ranges, and some survive only on protected offshore islands. Some of the extinctions have taken place in what is now the wheat belt of the southern tip of Western Australia, which retains only 5 percent of its natural woodland vegetation cover. Others have occurred in the southern arid zone, an area of mostly desert dominated by spinifex grass—and very few people. Nonetheless, this is an area where domestic grazing animals have destroyed the natural vegetation and caused extensive soil erosion. Moreover, European rabbits (Oryctolagus cuniculus) introduced in the mid-19th century are competitors of the native mammals, and the red fox (Vulpes vulpes) introduced about the same time has likely destroyed native small-mammal populations even in remote areas. Where foxes are absent in Australia or where their numbers have been controlled, the native mammals seem to have fared better. In addition, farmers and graziers have greatly altered the natural fire regimes—the roles that fire plays in ecosystems in the absence of human intervention—with consequent changes to the vegetation and the animals that depend on it.
Some of the extinctions have taken place in what is now the wheat belt of the southern tip of Western Australia, which retains only 5 percent of its natural woodland vegetation cover. Others have occurred in the southern arid zone, an area of mostly desert dominated by spinifex grass—and very few people. Nonetheless, this is an area where domestic grazing animals have destroyed the natural vegetation and caused extensive soil erosion. Moreover, European rabbits (Oryctolagus cuniculus) introduced in the mid-19th century are competitors of the native mammals, and the red fox (Vulpes vulpes) introduced about the same time has likely destroyed native small-mammal populations even in remote areas. Where foxes are absent in Australia or where their numbers have been controlled, the native mammals seem to have fared better. In addition, farmers and graziers have greatly altered the natural fire regimes—the roles that fire plays in ecosystems in the absence of human intervention—with consequent changes to the vegetation and the animals that depend on it.Calculating relative rates of extinction
To show how extinction rates are calculated, the discussion will focus on the group that is taxonomically the best-known—birds. The modern process of describing bird species dates from the work of the 18th-century Swedish botanist Carolus Linnaeus (Linnaeus, Carolus) in 1758. Even at that time, two of the species that he described were extinct, including the dodo. The 1800s was the century of bird description—7,079 species, or roughly 70 percent of the modern total, were named. Of those species, 39 became extinct in the subsequent 100 years. The corresponding extinction rate is 55 extinctions per million species per year. More than 100 of those 7,079 species are classified as critically endangered—the most threatened category of species listed by the IUCN (International Union for Conservation of Nature)—or else are dependent on conservation efforts to protect them. When similar calculations are done on bird species described in other centuries, the results are broadly similar.
Estimates of recent and future extinction ratesThe same approach can be used to estimate recent extinction rates for various other groups of plants and animals. One set of such estimates for five major animal groups—the birds discussed above as well as mammals, reptiles, frogs and toads, and freshwater clams—are listed in the table (Estimates of recent and future extinction rates). The calculated extinction rates, which range from 20 to 200 extinctions per million species per year, are high compared with the benchmark background rate of 1 extinction per million species per year, and they are typical of both continents and islands, of both arid lands and rivers, and of both animals and plants.
Although less is known about invertebrates than other species groups, it is clear from the case histories discussed above that high rates of extinction characterize both the bivalves of continental rivers and the land snails on islands. In fact, there is nothing special about the life histories of any of the species in the case histories that make them especially vulnerable to extinction. Indeed, what is striking is how diverse they are. The same is true for where the species live—high rates of extinction occur in a wide range of different ecosystems.
To draw reliable inferences from these case histories about extinctions in other groups of species requires that these be representative and not selected with a bias toward high extinction rates. In reviewing the list of case histories, it seems hard to imagine a more representative selection of samples. With high statistical confidence, they are typical of the many groups of plants and animals about which too little is known to document their extinction. In short, one can be certain that the present rates of extinction are generally pathologically high even if most of the perhaps 10 million living species have not been described or if not much is known about the 1.5 million species that have been described.
Predicting future rates of extinction
Not only do the five case histories demonstrate recent rates of extinction that are tens to hundreds of times higher than the natural rate, but they also portend even higher rates for the future. For every recently extinct species in a major group, there are many more presently threatened species. IUCN (International Union for Conservation of Nature) Red Lists in the early years of the 21st century reported that about 12 percent (i.e., about 1,200 species) of the roughly 10,000 living bird species are at risk of extinction. Furthermore, information in the same source indicates that this percentage is lower than that for mammals, reptiles, fish, flowering plants, or amphibians. For example, 16 percent of flowering plants are deemed threatened among the roughly 300,000 described species. Plant conservationists estimate that 100,000 plant species remain to be described, the majority of which will likely turn out to be rare and very local in their distribution. The latter characteristics explain why these species have not yet been found; they also make the species particularly vulnerable to extinction. Should any of these plants be described, they are likely to be classified as threatened, so the figure of 16 percent is an underestimate.
The Nature Conservancy in 1997 issued a report card on species for the United States that covered 13 groups of animals and plants. Using standards that were different, though broadly comparable, to those of the IUCN, it found that nearly 15 percent of birds were in danger of extinction (again the smallest percentage), between 16 and 18 percent of mammals, butterflies, reptiles, and dragonflies, up to about 40 percent of freshwater fish and amphibians, and more than two-thirds of freshwater mussels.
Once again choosing birds as a starting point, let us assume that the threatened species might last a century—this is no more than a rough guess. The 1,200 species of birds at risk would then suggest a rate of 12 extinctions per year on average for the next 100 years. That translates to 1,200 extinctions per million species per year, or 1,200 times the benchmark rate. This number, uncertain as it is, suggests a massive increase in the extinction rate of birds and, by analogy, of all other species, since the percentage of species at risk in the bird group is estimated to be lower than the percentages in other groups of animals and plants. Improving on this “rough guess” requires a more-detailed assessment of the fates of different sets of species.
Inexorable declines
 Some threatened species are declining rapidly. For a proportion of these, eventual extinction in the wild may be so certain that conservationists may attempt to take them into captivity to breed them (see below Protective custody (conservation)). Recent examples include the California condor (Gymnogyps californianus), which has been reintroduced into the wild with some success, and the alala (or Hawaiian crow, Corvus hawaiiensis), which has not. Other species have not been as lucky. In the early 21st century an exhaustive search for the baiji (Lipotes vexillifer), a species of river dolphin found in the Yangtze River, failed to find any. The dolphin had declined in numbers for decades, and efforts to keep the species alive in captivity were unsuccessful.
Some threatened species are declining rapidly. For a proportion of these, eventual extinction in the wild may be so certain that conservationists may attempt to take them into captivity to breed them (see below Protective custody (conservation)). Recent examples include the California condor (Gymnogyps californianus), which has been reintroduced into the wild with some success, and the alala (or Hawaiian crow, Corvus hawaiiensis), which has not. Other species have not been as lucky. In the early 21st century an exhaustive search for the baiji (Lipotes vexillifer), a species of river dolphin found in the Yangtze River, failed to find any. The dolphin had declined in numbers for decades, and efforts to keep the species alive in captivity were unsuccessful.Some species have no chance for survival even though their habitat is not declining continuously. Those who claim that extraordinary species such as the famous Loch Ness monster (“Nessie”) have long been surviving as solitary individuals or very small mating populations overlook the basics of sexual reproduction. If a species, be it proved or only rumoured to exist, is down to one individual—as some rare species are—then it has no chance. The odds are not much better if there are a few more individuals. If one breeding pair exists and if that pair produces two young—enough to replace the adult numbers in the next generation—there is a 50-50 chance that those young will be both male or both female, whereupon the population will go extinct. Even if they were male and female, they would be brother and sister, and their progeny would likely suffer from a variety of genetic defects (see inbreeding). In the case of two breeding pairs—and four young—the chance is one in eight that the young will all be of the same sex. These are better odds, but if the species plays this game every generation, only replacing its numbers, over many generations the probability is high that one generation will have four young of the same sex and so bring the species to extinction. Thus, for just one Nessie to be alive today, its numbers very likely would have to have been substantial just a few decades ago. These and related probabilities can be explored mathematically, and such models of small populations provide crucial advice to those who manage threatened species.
Field studies of very small populations have been conducted. For example, small islands off the coast of Great Britain have provided a half-century record of many bird species that traveled there and remained to breed. This record shows that most small populations formed by individuals that colonized from the mainland persisted for a few years to decades before going extinct
Surviving but threatened small populations
A broad range of environmental vagaries, such as cold winters, droughts, disease, and food shortages, cause population sizes to fluctuate considerably from year to year. The behaviour of butterfly populations is well studied in this regard. Because their numbers can decline from one year to the next by 99 percent, even quite large populations may be at risk of extinction. In the case of smaller populations, the Nature Conservancy reported that, of about 600 butterfly species in the United States, 16 species number fewer than 3,000 individuals and another 74 species fewer than 10,000 individuals. Butterfly numbers are hard to estimate, in part because they do fluctuate so much from one year to the next, but it is clear that such natural fluctuations could reduce low-population species to numbers that would make recovery unlikely.
The role of population fluctuations has been dissected in some detail in a long-term study of the Bay checkerspot butterfly (Euphydryas editha bayensis) in the grasslands (grassland) above Stanford University in Palo Alto, Calif. In 1960 scientists began following the fate of several local populations of the butterfly at a time when grasslands around San Francisco Bay were being lost to housing developments. The populations were themselves isolated from each other, with only little migration between them. In succeeding decades small populations went extinct from time to time, but immigrants from two larger populations reestablished them. Nonetheless, in 1991 and 1998 first one and then the other larger population became extinct. At their peaks the former had reached almost 10,000 individuals and the latter about 2,000 individuals, although this second population was less variable from year to year. The Bay checkerspot still lives in other places, but the study demonstrates that relatively small populations of butterflies (and, by extension, other insects) whose numbers undergo great annual fluctuations can become extinct quickly.
What are the consequences of these fluctuations for future extinctions worldwide? Simply put, habitat destruction has reduced the majority of species everywhere on Earth to smaller ranges than they enjoyed historically. Where these ranges have shrunk to tiny protected areas, species with small populations have no possibility of expanding their numbers significantly, and quite natural fluctuations (along with the reproductive handicaps of small populations) can exterminate species.
In sum, most of the presently threatened species will likely not survive the 21st century. They may already be declining inexorably to extinction; alternately, their populations may number so few that they cannot survive more than a few generations or may not be large enough to provide a hedge against the risk that natural fluctuations will eventually lead to their extinction.
Because some threatened species will survive through good luck and others by good management of them, estimates of future extinction rates that do not account for these factors will be too high. A factor having the potential to create more serious error in the estimates, however, consists of those species that are not now believed to be threatened but that could become extinct. For example, the 2006 IUCN Red List for birds added many species of seabirds that formerly had been considered too abundant to be at any risk. Over the previous decade or so, the growth of longline fishing, a commercial technique in which numerous baited hooks are trailed from a line that can be kilometres long (see commercial fishing: Drifting longlines (commercial fishing); Bottom longlines (commercial fishing)), has caused many seabirds, including most species of albatross, to decline rapidly in numbers. Albatrosses follow longlining ships to feed on the bait put on the lines' hooks. The birds get hooked and then drown.
Accidentally or deliberately introduced species have been the cause of some quick and unexpected extinctions. For example, about 1960 the unique birds of the island of Guam appeared to be in no danger, for many species were quite common. Yet a reptile, the brown tree snake (Boiga irregularis; see mangrove snake), had been accidentally introduced perhaps a decade earlier, and, as it spread across the island, it systematically exterminated all the island's land birds. The snakes occasionally stow away in cargo leaving Guam, and, since there is substantial air traffic from Guam to Honolulu, Hawaii, some snakes arrived there. None are thought to have survived, but, should the snake establish a population there, the Hawaiian Islands would likely lose all their remaining native birds.
Although anticipating the effect of introduced species on future extinctions may be impossible, it is fairly easy to predict the magnitude of future extinctions from habitat loss, a factor that is simple to quantify and that is usually cited as being the most important cause of extinctions. (For birds, to give an example, some three-fourths of threatened species depend on forests, mostly tropical ones that are rapidly being destroyed.) Habitat destruction is continuing and perhaps accelerating, so some now-common species certainly will lose their habitat within decades. Even so, making specific predictions requires a more-detailed understanding of the factors that cause extinctions, which are addressed in a following section (conservation).
The loss of populations
While much of the concern over the loss of biodiversity centres on the global loss of species, most of the benefits that biodiversity confers depend on the continued existence of sufficient numbers of the local populations of species. An obvious example is loss of the forests that protect a city's watershed. In preventing soil erosion, these forests and their species provide a local service that would cease if the forests were cleared, irrespective of whether their species still existed elsewhere in the world. In addition, local populations supply genetic diversity, because different populations across a species' range differ to varying degrees in their genetic composition. Such genetic diversity has significant value for agricultural plant breeders, who rely on wild relatives of crop plants for novel genes that confer resistance to plant pathogens such as viruses and fungi. Thus, as populations are eliminated locally, potentially beneficial genes may become lost globally.
Scientists estimate than an average species consists of 220 genetically distinct populations, suggesting that there may be more than 2 billion such populations globally. They estimate that 160 million populations (8 percent) are lost each decade. If this trend continues, even though many species may be saved in protected areas (such as national parks), each of them will be merely a remnant of a once geographically extensive and genetically diverse set of populations.
The loss of ecosystems (ecosystem)
 Conservation justifiably prioritizes tropical moist forests (see tropical forest (tropical rainforest)) because they hold such a large fraction of the global total of species, but a comprehensive strategy must also include the conservation of other distinctive ecosystems. For example, major habitat types such as tropical dry forests, tundra, temperate grasslands, lakes, polar seas (see Arctic Ocean; Southern Ocean), and mangroves all contain characteristic species. Importantly, their biological communities house distinctive ecological and evolutionary phenomena. Some of these ecosystems are more threatened than are tropical moist forests and require immediate conservation action.
Conservation justifiably prioritizes tropical moist forests (see tropical forest (tropical rainforest)) because they hold such a large fraction of the global total of species, but a comprehensive strategy must also include the conservation of other distinctive ecosystems. For example, major habitat types such as tropical dry forests, tundra, temperate grasslands, lakes, polar seas (see Arctic Ocean; Southern Ocean), and mangroves all contain characteristic species. Importantly, their biological communities house distinctive ecological and evolutionary phenomena. Some of these ecosystems are more threatened than are tropical moist forests and require immediate conservation action.To provide a more representative selection of Earth's distinctive ecosystems, scientists working in the United States at the World Wildlife Fund (WWF) (WWF) have compiled what they call the world's “ecoregions,” which are drawn from the planet's terrestrial, freshwater, and marine major habitat types. Based on a comparative global analysis, they have identified some 240 ecoregions as global priorities—about 140 terrestrial, 50 freshwater, and 40 marine areas that are the most biologically outstanding, either for their richness of species or for their ecological or evolutionary uniqueness.
 A comparison of the WWF's list of global priorities and a list of priorities based on threatened species alone uncovers many overlaps, predominantly in the tropical moist forests. It is the differences that are interesting, however. The Everglades of Florida and Brazil's Pantanal, for example, do not rank as places with high concentrations of threatened species, but they achieve prominence among the WWF's global ecoregions because they are among the few flooded grasslands (grassland) in the world, all of which are under threat. Other threatened ecosystems identified on WWF's list include the dry forests of lowland Bolivia, southern Mexico, Madagascar, and Hawaii. The temperate grasslands of North America (see Great Plains), another global priority, once sustained vast migrations of large vertebrates such as bison, pronghorn, and elk. Such species are now restricted to small pockets. The same is true in Eurasia, where comparable migrations now occur only in isolated pockets, primarily in the Daurian Steppe (Steppe, the) in China, Mongolia, and Russia and the Tibetan Plateau. And the extraordinary floral communities of the Eurasian steppes and the North American Great Plains have been largely destroyed through the land's conversion to agriculture (agriculture, origins of).
A comparison of the WWF's list of global priorities and a list of priorities based on threatened species alone uncovers many overlaps, predominantly in the tropical moist forests. It is the differences that are interesting, however. The Everglades of Florida and Brazil's Pantanal, for example, do not rank as places with high concentrations of threatened species, but they achieve prominence among the WWF's global ecoregions because they are among the few flooded grasslands (grassland) in the world, all of which are under threat. Other threatened ecosystems identified on WWF's list include the dry forests of lowland Bolivia, southern Mexico, Madagascar, and Hawaii. The temperate grasslands of North America (see Great Plains), another global priority, once sustained vast migrations of large vertebrates such as bison, pronghorn, and elk. Such species are now restricted to small pockets. The same is true in Eurasia, where comparable migrations now occur only in isolated pockets, primarily in the Daurian Steppe (Steppe, the) in China, Mongolia, and Russia and the Tibetan Plateau. And the extraordinary floral communities of the Eurasian steppes and the North American Great Plains have been largely destroyed through the land's conversion to agriculture (agriculture, origins of).Factors that cause extinction
Habitat loss
As mentioned above, habitat loss is widely listed as the predominant cause of extinction. It is not hard to understand why—clear a forest, destroy a coral reef, or dam a river, and the species found there will likely be lost. These are instances of local extinctions, however, and their occurrence does not mean that the species involved will go extinct everywhere, as is the situation for the global extinctions discussed in the case histories above. Some scientists use the term extirpation for local extinctions, reserving extinction to mean global extinction. In this section on factors causing extinction, the term has the global meaning.
On land
In the preceding case histories, the roles of habitat loss in extinction of species are fairly clear-cut. There are seeming counterexamples—Europe and eastern North America, for example—where human actions have extensively modified terrestrial habitats, yet these areas are not centres of extinction. Clearly, habitat destruction causes different numbers of extinctions in different places.
Endemic species
 To learn what makes centres of human-caused extinctions special, one can ask what are their common features. Obviously, many of these places and their species are well known. The 17th-century Dutch artist Rembrandt van Rijn painted birds-of-paradise (bird-of-paradise) and marine cone shells gathered from the early European exploration of the Pacific, testifying to people's fascination for attractive and interesting specimens from “exotic” locales. Victorians filled their cabinets with such curiosities—birds, mammals, marine and terrestrial snail shells, and butterflies—and painted tropical flowers and grew them in their hothouses. Yet the natural history of North America and Europe are also very well known. In fact, what is special about the places with so many extinctions is that each area holds a high proportion of species restricted to it. Scientists call such species endemics.
To learn what makes centres of human-caused extinctions special, one can ask what are their common features. Obviously, many of these places and their species are well known. The 17th-century Dutch artist Rembrandt van Rijn painted birds-of-paradise (bird-of-paradise) and marine cone shells gathered from the early European exploration of the Pacific, testifying to people's fascination for attractive and interesting specimens from “exotic” locales. Victorians filled their cabinets with such curiosities—birds, mammals, marine and terrestrial snail shells, and butterflies—and painted tropical flowers and grew them in their hothouses. Yet the natural history of North America and Europe are also very well known. In fact, what is special about the places with so many extinctions is that each area holds a high proportion of species restricted to it. Scientists call such species endemics.Remote islands have many terrestrial endemics—for instance, more than 90 percent of the plants and land birds of the Hawaiian Islands live nowhere else. Some continental areas are rich in endemic species, too. About 70 percent of the flowering plants in South Africa's fynbos and nearly three quarters of Australia's mammals are endemic. In contrast, many areas have almost no endemic species; only about 1 percent of Europe's birds are found only there.

 The simplest model of extinction would be to assume that within a species group—mammals, for instance—all species had roughly the same risk of extinction. Were this to be true, then the more species that live in a region, the more would likely go extinct. This is not the case, however. For example, the Hawaiian Islands and Great Britain in their pasts held very roughly the same number of breeding land birds. Yet, the former have lost more than 100 species, as discussed in the Pacific Islands case study above, while the latter has lost only a few—and those still survive on the European continent. The difference is that almost all the Hawaiian birds (for instance, honeycreepers (Hawaiian honeycreeper) such as the apapane and iiwi) were endemic, while only one of Great Britain's birds (the Scottish crossbill) is. Thus, the number of extinctions in an area depends only very weakly on its total number of species but strongly on its total number of endemics. Areas rich in endemic species are where extinctions will concentrate, unless they are so remote that human actions do not harm them.
The simplest model of extinction would be to assume that within a species group—mammals, for instance—all species had roughly the same risk of extinction. Were this to be true, then the more species that live in a region, the more would likely go extinct. This is not the case, however. For example, the Hawaiian Islands and Great Britain in their pasts held very roughly the same number of breeding land birds. Yet, the former have lost more than 100 species, as discussed in the Pacific Islands case study above, while the latter has lost only a few—and those still survive on the European continent. The difference is that almost all the Hawaiian birds (for instance, honeycreepers (Hawaiian honeycreeper) such as the apapane and iiwi) were endemic, while only one of Great Britain's birds (the Scottish crossbill) is. Thus, the number of extinctions in an area depends only very weakly on its total number of species but strongly on its total number of endemics. Areas rich in endemic species are where extinctions will concentrate, unless they are so remote that human actions do not harm them.Range size
Given that species differ in their risk of extinction, the size of a species' geographic range is by far the best explanation for the differences. Species with small ranges are much more vulnerable than those with large ranges, simply because it is much easier to destroy the former than the latter.
In regard to range size, the distribution of life on land has several remarkable features. First, many terrestrial species have very small range sizes relative to the average range size. In the Americas, for example, 1 in 10 species of birds and over half the species of amphibians have geographic ranges smaller than the state of Connecticut, and half the bird species have ranges smaller than the states of Washington, Oregon, and California combined. The average range size is very much larger, for some species have huge ranges. The American robin (Turdus migratorius), for example, breeds almost everywhere in the United States from Alaska to Florida to California, across all of continental Canada, and in much of Mexico.
Second, for many kinds of terrestrial organisms, species with small ranges typically have lower local population densities than do widespread species. For example, the American robin is generally a locally common bird across its entire range. But those species with ranges smaller than the size of Connecticut are generally very hard to find even in the midst of their ranges.
 Third, terrestrial species with small ranges are geographically concentrated. North America, for instance, has almost no bird species with small ranges. Such species live almost exclusively in the tropics. The greatest concentrations of bird species are in the Amazon lowlands, with secondary centres in Central America and the forests along the coast of Brazil, as can be seen in the accompanying maps-->, which show distributions of songbird species in the Americas. But it is the Andes and the coastal forests that have the most bird species with small geographic ranges; i.e., these areas are the centres of endemism. And it is in these areas that threatened species are concentrated.
Third, terrestrial species with small ranges are geographically concentrated. North America, for instance, has almost no bird species with small ranges. Such species live almost exclusively in the tropics. The greatest concentrations of bird species are in the Amazon lowlands, with secondary centres in Central America and the forests along the coast of Brazil, as can be seen in the accompanying maps-->, which show distributions of songbird species in the Americas. But it is the Andes and the coastal forests that have the most bird species with small geographic ranges; i.e., these areas are the centres of endemism. And it is in these areas that threatened species are concentrated.Terrestrial hot spots
In the 1990s a team of researchers led by British environmental scientist Norman Myers identified 25 terrestrial “hot spots” of the world—25 areas on land where species with small geographic ranges coincide with high levels of modern human activity (see the map-->). Originally these hot spots encompassed about 17 million square km (6.6 million square miles) of the roughly 130 million square km (50 million square miles) constituting Earth's ice-free land surface. Species ranges are so concentrated geographically in these regions that, out of a total of about 300,000 flowering-plant species described worldwide, more than 133,000 occur only there. The comparable numbers for birds are roughly 2,800 of 10,000 species worldwide (of which roughly two-thirds are restricted to the land); for mammals, 1,300 of roughly 5,000 worldwide; for reptiles, roughly 3,000 of about 8,000 worldwide; and for amphibians, 2,600 of roughly 5,000 worldwide. world's 25 hot spotsThe hot spots have been sites of unusual levels of habitat destruction. Only about one-eighth of the original habitat of these areas has survived to the beginning of the 21st century. Of this remaining habitat, only about two-fifths is protected in any way. Sixteen of the 25 hot spots are forests, most of them tropical forests. For comparison, the relatively less-disturbed forests found in the Amazon, the Congo region, and New Guinea have retained about half their original habitats. As a consequence of these high levels of habitat loss, the 25 hot spots are locations where the majority of threatened and recently extinct species are to be found. For additional details about the hot spots discussed above, see the table (world's 25 hot spots).
Predictions of extinctions based on habitat loss
Worldwide, about 6 percent of the land surface is protected by some form of legislation, though the figure for the 25 hot spots is only 4.5 percent of their original extent. (Such numbers are misleading, however, in that some areas are protected only on paper as their habitats continue to be destroyed.) These statistics lead naturally to the question of how many species will be saved if, say, 4.5 percent of the land worldwide is protected. Will only 4.5 percent of its species be saved? The answer turns out to be closer to 50 percent, a result that needs some explanation.
 The land that countries protect, often as national or regional parks, frequently comprises “islands” of original habitat surrounded by a “sea” of cropland, grazing land, or cities and roads. On a real island the number of species that live there depends on its area, with a larger island housing more species than a smaller one. Many studies involving a wide range of animals and plants show that the relationship between area and species number is remarkably consistent. An island half the size of another will hold about 85 percent of the number of species.
The land that countries protect, often as national or regional parks, frequently comprises “islands” of original habitat surrounded by a “sea” of cropland, grazing land, or cities and roads. On a real island the number of species that live there depends on its area, with a larger island housing more species than a smaller one. Many studies involving a wide range of animals and plants show that the relationship between area and species number is remarkably consistent. An island half the size of another will hold about 85 percent of the number of species.The next question is whether this relationship of species to area holds for “islands” of human-created habitat. If one were to conduct an experiment to test such an idea, one would take a continuous forest, cut it up into isolated patches, and then wait for species to become locally extinct in them. After sufficient time, one could count the numbers of species remaining and relate them to the area of the patches in which they survived. In the last decades of the 20th century, scientists undertook exactly such an experiment, making use of government-approved forest clearing for cattle ranching, in the tropical forests around Manaus in Brazil. More generally, human actions have repeated this experiment across much of the planet in an informal way. Counts of species in areas of different sizes confirm the species-to-area relationship.
 How can these results regarding local habitat “islands” be applied to global extinctions? The mostly deciduous forests of eastern North America provide a case history. The birds of the region have been well-described beginning with the explorations of the naturalist and artist John James Audubon (Audubon, John James) in the first quarter of the 19th century, when the area was still mostly forested. Audubon shot and painted many species including four that are now extinct—the Carolina parakeet (Conuropsis carolinensis; see conure), Bachman's warbler (Vermivora bachmanii; see woodwarbler (wood warbler)), the passenger pigeon (Ectopistes migratorius), and (if not extinct, then very nearly so) the ivory-billed woodpecker (Campephilus principalis). A fifth species, the red-cockaded woodpecker (Picoides borealis), is threatened. Including the above, about 160 species of birds once lived in these eastern forests.
How can these results regarding local habitat “islands” be applied to global extinctions? The mostly deciduous forests of eastern North America provide a case history. The birds of the region have been well-described beginning with the explorations of the naturalist and artist John James Audubon (Audubon, John James) in the first quarter of the 19th century, when the area was still mostly forested. Audubon shot and painted many species including four that are now extinct—the Carolina parakeet (Conuropsis carolinensis; see conure), Bachman's warbler (Vermivora bachmanii; see woodwarbler (wood warbler)), the passenger pigeon (Ectopistes migratorius), and (if not extinct, then very nearly so) the ivory-billed woodpecker (Campephilus principalis). A fifth species, the red-cockaded woodpecker (Picoides borealis), is threatened. Including the above, about 160 species of birds once lived in these eastern forests.European settlement cleared these forests, and, at the low point about 1870, only approximately half the forest remained. The species-to-area relationship predicts that about 15 percent of the 160 bird species—that is, about 24 species—would become extinct. Why is this number much larger than the three to five extinctions or near-extinctions observed? The answer can be found by first supposing that all the eastern forests had been cleared, from Maine to Florida and westward to the prairies. Only those species that lived exclusively within these forests—that is, the endemics—would have gone extinct. Species having much larger ranges, such as the American robin mentioned above, would have survived elsewhere. How many species then were originally endemic to the forests of eastern North America? The answer is about 30, the rest having wide distributions across Canada and, for some, into Mexico. The species-to-area predicts that 15 percent of these 30 species, or 4.5 species, should go extinct, which is remarkably close to the observed number.
Eastern North America clearly is not a place where species with small ranges are concentrated, but the species-to-area predictions work in other places, too. A variety of studies have examined birds and mammals on the islands of Southeast Asia, from Sumatra westward—one of the biodiversity hot spots (Sundaland). Both here and for birds in Brazil's Atlantic Forest, which is another hot spot, the extent of deforestation and the species-to-area relationship accurately predict the number of threatened species rather than extinct ones. Because the deforestation of these areas is relatively recent, many of their species have not yet become extinct.
The close match between the numbers of extinctions predicted by the species-to-area relationship and the numbers of species already extinct (as in eastern North America) or nearly extinct (as in more recently destroyed habitats) allows simple calculations that have worldwide import. As discussed above, the hot spots retain only 12 percent of their original habitats, and only about 4.5 percent of the original habitats are protected. As predicted by the species-to-area relationship, natural habitats that have shrunk to 4.5 percent of their original extent will lose more than half of their species. Since these habitats once supported 30–50 percent of terrestrial species, very roughly one-fourth of all terrestrial species will likely become extinct.
 Species losses will likely be even greater because this calculation does not include nonendemic species—those that live both inside and outside the hot spots. For example, many of the species that live in the relatively less-disturbed tropical moist forests of the Amazon or the Congo region are the same that live in the adjacent hot spots. Human actions are clearing about 10 percent of the original area of these forests every decade, with a half of the area already gone. Species losses in these forests are still relatively few, but the rate will increase rapidly as the last remaining forests dwindle. If only the same percentage of these forests is protected as is the case for the hot spots, then they too will lose half their species.
Species losses will likely be even greater because this calculation does not include nonendemic species—those that live both inside and outside the hot spots. For example, many of the species that live in the relatively less-disturbed tropical moist forests of the Amazon or the Congo region are the same that live in the adjacent hot spots. Human actions are clearing about 10 percent of the original area of these forests every decade, with a half of the area already gone. Species losses in these forests are still relatively few, but the rate will increase rapidly as the last remaining forests dwindle. If only the same percentage of these forests is protected as is the case for the hot spots, then they too will lose half their species.In summary, many scientists believe that habitat destruction will put somewhere between a fourth and a half of all species on an inexorable path to extinction and will do so within the next few decades. If that proves true, extinction rates by the mid-21st century will be several thousand times the benchmark rate.
Fire (forest fire) suppression as habitat loss
Whereas most of the hot spots are tropical moist forests, four areas—the California Floristic Province, the Cape Floristic Province in South Africa, the Mediterranean Basin, and Southwest Australia (see the map-->)—are shrublands. They also are places where people live and grow crops; all four regions are noted for their wines, for example. Not only does this human activity convert land directly, but it also leads to the suppression of fire, especially near people's homes. This alteration of natural fire regimes by the reduction in fire frequencies leads to changes in vegetation, especially to the loss of the native fire-resistant species. Globally, huge areas of grasslands (grassland) and shrublands would become heavily canopied forests were all fires suppressed. The effects of changes in fire frequencies on species losses have not yet been calculated.In the oceans (ocean)
The seas cover more than two-thirds of Earth's surface, yet only 210,000 of the 1,500,000 species that have been described are marine animal and plant species. Because the oceans are still poorly explored, the count of marine species may be even more of an underestimate than that of land species. For example, the Census of Marine Life, a decade-long international program begun at the start of the 21st century, added 13,000 new marine species to the total count over the first four years of the effort. As on land, the peak of marine biodiversity lies in the tropics. coral reefs account for almost 100,000 species, yet their total area represents just 0.2 percent of the ocean surface. Between 4,000 and 5,000 species of fish—perhaps a third of the world's marine fish—live on coral reefs. The frequently cited metaphor that “coral reefs are the rainforests of the sea” underscores their importance for conservation.
When numbers of described marine species are mapped on a worldwide scale, it becomes clear that the global centre of marine biodiversity encompasses the waters of the Philippine and Indonesian islands. Numbers of species drop steeply to the east across the Pacific and less steeply to the west across the Indian Ocean. In the Atlantic Ocean the highest levels of biodiversity are in the Caribbean. fish, corals, mollusks, and lobsters all show similar patterns in the distributions of their species. Again mirroring the patterns on land, the places with the most species are often not the places with the most endemic species. Major centres of endemism for fish, corals, mollusks, and lobsters include the Philippines, southern Japan, the Gulf of Guinea, the Sunda Islands, and the Mascarene Islands.
With the major exception of the Great Barrier Reef of Australia, most coral reefs (coral reef) are off the coasts of developing countries. Rapidly increasing human populations and poverty put increasing fishing pressure on nearshore reefs. In addition, in their efforts to sustain declining fish catches, people resort to extremely damaging fishing methods such as dynamite and poisons. Coral reefs are also threatened by coastal development, pollution, and global warming. Human activities threaten some three-fifths of the world's reefs, with the highest damage being concentrated in areas having high rates of deforestation and high runoff from the land. As the inhabitants of an area destroy their tropical forests, rains erode soils and wash the sediments down rivers into the sea, damaging the local coral reefs. Thus, the destruction of some of the most important terrestrial habitats—in the Caribbean and Southeast Asia—contributes to the destruction of some of the most important marine habitats offshore.
Whereas damage to coral reefs is important for the loss of species, the greatest physical damage to ocean ecosystems involves the effects of bottom trawling, a commercial fishing method making use of a cone-shaped bag of netting that is dragged along the seabed (see commercial fishing: Dragged gear (commercial fishing)). Damage from bottom trawling occurs over larger areas of Earth than does tropical deforestation, and it involves even greater and more-frequent disturbances, albeit ones not easily seen. Bottom trawling disturbs about 15 million square km (6 million square miles) of the world's seafloor each year. This area of ocean is only about 4 percent of the world total, but its small proportion belies its significance. About 90 percent of the ocean consists of deep waters so poor in nutrients that they are the equivalent of the land's deserts. Almost all of the world's fisheries are concentrated in the 30 million square km (12 million square miles) of nutrient-rich waters that are on the continental shelf, plus a few upwellings. On average, the ocean floor of these productive waters is trawled roughly every two years.
The otter trawl is the most widely used bottom-fishing gear. As it is dragged forward, a pair of flat plates called otter boards—one on each side of the trawl net and weighing several tons—spreads horizontally to keep the mouth of the trawl open; at the same time, a long rope with steel weights keeps the mouth open along its bottom edge. This heavy structure plows the ocean floor as it moves, creating furrows and crushing, burying, and exposing marine life. This activity destroys bottom-dwelling species including corals, brachiopods ( lamp shells), mollusks, sponges, sea urchins, and various worms that live on rocks or pebbles on the seabed. It is also damaging to other species, such as polychaete worms that burrow into the seabed. Some species—deep-sea corals, for example—are extremely slow-growing, and they cannot recover before bottom trawls plow the area once again.
In fresh water
Freshwater ecosystems are divided into two major classes—flowing (such as rivers and streams) and static (such as lakes and ponds). Although the distribution of species in freshwater ecosystems is not as well-known as for marine and terrestrial ecosystems, it is still clear that species are similarly concentrated. For fish, the major tropical rivers such as the Amazon River and its tributaries hold a large fraction of the world's freshwater fish species. Tropical lakes, particularly those in the Rift Valley of East Africa (see East African Rift System), also have large numbers of endemic species.
Riverine habitats have been extensively modified by damming and by channelization, the latter being the practice of straightening rivers by forcing them to flow along predetermined channels. A global survey published in the early 21st century revealed just how few of the world's large rivers are natural. In the contiguous United States almost every large river has been modified extensively. (As previously noted, some rivers such as the Tennessee River have been converted almost completely by dams to a series of artificial lakes.) Much the same is true in Europe. In terms of the area of their basins, more than half of the world's rivers are extensively modified. The water of some rivers barely reaches its final destination; this is the case for the Colorado River in the United States, which empties into the Gulf of California, and for the Amu Darya in Central Asia, which empties into the rapidly shrinking Aral Sea. Along their routes the water is used for agriculture (agriculture, origins of) or lost as evaporation from dams. Large wild rivers are typical only of Arctic regions in Alaska, Canada, and Siberia—places so far away from urban centres that there has been no incentive to control their waters. The massive changes to the world's rivers explain why such large fractions of species living in rivers have become extinct or may do so soon (as is described in two of the preceding case histories; see above Freshwater mussels and clams (conservation); Freshwater fish (conservation)).
Pollution
pollution is a special case of habitat destruction; it is chemical destruction rather than the more obvious physical destruction. Pollution occurs in all habitats—land, sea, and fresh water—and in the atmosphere. global warming, which is discussed separately below (see Global change (conservation)), is one consequence of the increasing pollution of the atmosphere by emissions of carbon dioxide and other gases.
Pollution is a global-scale problem, no less so for rivers (river) and marine life. Wastes are often dumped into rivers, and they end up in estuaries and coastal habitats, regions that support the most diverse shallow-water ecosystems and the most productive fisheries. Rivers receive pollution directly from factories that dump a wide variety of wastes into them. They also receive runoff, which is rainwater that has passed over and through the soil while moving toward the rivers. In fact, water entering rivers after it has been used for irrigation has passed through the soil more than once—first as runoff, which is then returned to the land for irrigation, whereupon it soaks through the soil again.
Some polluted river water eventually reaches freshwater wetlands. In the case of the Florida Everglades, runoff from the agricultural areas upstream adds unwanted nutrients to an ecosystem that is naturally nutrient-poor. As it does so, the vegetation changes, and species not common in the Everglades begin to take over the natural habitats.
Other polluted waters reach estuaries on the way to the oceans; estuaries are among the most polluted ecosystems on Earth. On entering the oceans, the polluted waters can harm the ecosystems there. The Mississippi River, for example, drains a basin of more than 3 million square km (1.2 million square miles), delivering its water, sediments, and nutrients and other pollutants into the northern Gulf of Mexico. Fresh water is less dense than salt water and floats on top. This upper layer contains the nitrogen and phosphorus fertilizers that have run off croplands, and they fertilize the ocean's phytoplankton, causing excessive population growth. As the masses of phytoplankton die, sink, and decompose, they deplete the water's oxygen. Bottom dwellers such as shrimp, crabs, starfish, and marine worms suffocate and die, creating a “dead zone.”
Such conditions also occur in Europe's Baltic (Baltic Sea), Adriatic (Adriatic Sea), and Black (Black Sea) seas. The Baltic (Baltic Sea) has gone from being naturally nutrient-poor and diverse in species to being nutrient-rich and degraded in its ecosystems within a few decades. In the Adriatic Sea, rising nutrient levels have generated a large increase in phytoplankton. Nutrients in the runoff flowing into the Black Sea seem to be contributing factors in the invasion and subsequent massive increase since the 1980s of the comb jelly Mnemiopsis leidyi. This has caused the decline of native species and fisheries.
Similar nutrient enrichment has led to increasing frequencies of toxic blooms (water bloom) of microscopic organisms such as Pfiesteria piscicida in the eastern United States, a dinoflagellate that kills fish and has been reported to cause skin rashes and other maladies in humans.
Rising levels of pollution may have also contributed to a wave of outbreaks of diseases affecting marine life. Caribbean coral reefs have been particularly affected, with successive waves of disease propagating throughout the region in recent decades. The result has been large declines in two species of major reef-building corals, Acropora cervicornis and A. palmata, and the herbivorous sea urchin Diadema antillarum. Their combined loss has transformed Caribbean reefs from high-coral, low-algae ecosystems to high-algae, low-coral ones. The latter type of ecosystems support far fewer species.
introduced species
 The case histories previously discussed often implicate introduced species as a cause of species extinctions. Humans have spread species deliberately as they colonized new areas, just one example being the Polynesians as they settled the eastern Pacific Islands. New Yorkers in the 1890s wanted all the birds in Shakespeare (Shakespeare, William)'s works to inhabit the city's Central Park, and they introduced the starling (Sturnus vulgaris) to North America as a consequence. Through the centuries hunters have demanded exotic birds and mammals to shoot, fishermen have wanted challenging fish, and gardeners have wanted beautiful flowers. Nonetheless, the consequences in some cases have been devastating. Cacti (cactus) and the shrub Lantana camara, for example, which were introduced as ornamental plants, have destroyed huge areas of grazing land worldwide.
The case histories previously discussed often implicate introduced species as a cause of species extinctions. Humans have spread species deliberately as they colonized new areas, just one example being the Polynesians as they settled the eastern Pacific Islands. New Yorkers in the 1890s wanted all the birds in Shakespeare (Shakespeare, William)'s works to inhabit the city's Central Park, and they introduced the starling (Sturnus vulgaris) to North America as a consequence. Through the centuries hunters have demanded exotic birds and mammals to shoot, fishermen have wanted challenging fish, and gardeners have wanted beautiful flowers. Nonetheless, the consequences in some cases have been devastating. Cacti (cactus) and the shrub Lantana camara, for example, which were introduced as ornamental plants, have destroyed huge areas of grazing land worldwide.Not all introductions are deliberate. Several species of rats, for instance, were likely unwelcome hitchhikers on ocean voyages. Humans have spread infectious diseases around the world with consequences that dramatically changed human history; a recent example is AIDS, spread from Africa by individuals infected with the human immunodeficiency virus (HIV). Introduction of the tree snake that destroyed Guam's birds was also accidental.
Almost every type of living organism has been moved deliberately or accidentally. A study published in the 1990s surveyed species that traveled in the ballast tanks of oceangoing ships. Such ships take on seawater as ballast in one port and release it in other ports sometimes half a world away. In one example, water taken aboard in Japan and released in Oregon contained 360 species including microscopic single-celled animals and plants, jellyfish and corals, mollusks, various kinds of marine worms, and fish—indeed, an almost complete catalog of major kinds of marine organisms.
 Although not all species devastate the communities they enter, a closer look at what happened on Guam after the introduction of the tree snake Boiga illustrates just how damaging a species can be. Birds started disappearing from the central region of the island in the early 1960s. At the same time, the snakes started causing blackouts as they crawled up power poles and short-circuited the suspended lines, and many people who kept chickens noticed that snakes were killing the birds in their coops. Before Boiga made its appearance, 10 species of native birds lived in the forests of Guam—the Micronesian starling (Aplonis opaca), the Mariana crow (Corvus kubaryi), the Micronesian kingfisher (Halcyon cinnamomina), the Guam flycatcher (Myiagra freycineti), the Guam rail (Rallus owstoni), the rufous fantail (Rhipidura rufifrons), the bridled white-eye (Zosterops conspicillatus), the white-throated ground dove (Gallicolumba xanthonura), the Mariana fruit dove (Ptilinopus roseicapilla), and the cardinal honeyeater (Myzomela cardinalis). An 11th species, the island swiftlet (Aerodramus vanikorensis), nested in caves and fed over the forest. In addition, the island supported a few introduced bird species, birds that nested in wetland habitats, and some seabirds. The 10 forest birds declined following similar patterns—first on the central part of the island, where the main town and port are, and then on southern Guam by the late 1960s. By 1983 scientists could find the birds only in a small patch of forest in the north of the island, and by 1986 they were gone from there as well. The nonnative birds suffered a similar pattern of decline, and many of the waterbirds and seabirds that nested on the island also declined. The starling has survived on a small island, Cocos, off the south coast of Guam, and the swiftlet still nests on the high walls of a cave. The snake has eliminated every species for which it could easily kill the eggs and young in the nest. Of the 10 forest species, the rail and the flycatcher were endemic to the island, while the kingfisher is a distinctive subspecies. The kingfisher and the rail survive in captivity.
Although not all species devastate the communities they enter, a closer look at what happened on Guam after the introduction of the tree snake Boiga illustrates just how damaging a species can be. Birds started disappearing from the central region of the island in the early 1960s. At the same time, the snakes started causing blackouts as they crawled up power poles and short-circuited the suspended lines, and many people who kept chickens noticed that snakes were killing the birds in their coops. Before Boiga made its appearance, 10 species of native birds lived in the forests of Guam—the Micronesian starling (Aplonis opaca), the Mariana crow (Corvus kubaryi), the Micronesian kingfisher (Halcyon cinnamomina), the Guam flycatcher (Myiagra freycineti), the Guam rail (Rallus owstoni), the rufous fantail (Rhipidura rufifrons), the bridled white-eye (Zosterops conspicillatus), the white-throated ground dove (Gallicolumba xanthonura), the Mariana fruit dove (Ptilinopus roseicapilla), and the cardinal honeyeater (Myzomela cardinalis). An 11th species, the island swiftlet (Aerodramus vanikorensis), nested in caves and fed over the forest. In addition, the island supported a few introduced bird species, birds that nested in wetland habitats, and some seabirds. The 10 forest birds declined following similar patterns—first on the central part of the island, where the main town and port are, and then on southern Guam by the late 1960s. By 1983 scientists could find the birds only in a small patch of forest in the north of the island, and by 1986 they were gone from there as well. The nonnative birds suffered a similar pattern of decline, and many of the waterbirds and seabirds that nested on the island also declined. The starling has survived on a small island, Cocos, off the south coast of Guam, and the swiftlet still nests on the high walls of a cave. The snake has eliminated every species for which it could easily kill the eggs and young in the nest. Of the 10 forest species, the rail and the flycatcher were endemic to the island, while the kingfisher is a distinctive subspecies. The kingfisher and the rail survive in captivity.Islands (island) such as Guam seem to be particularly vulnerable to introduced species. People have often brought domestic cat (cat, domestic)s with them to islands, where some of the animals have escaped to form feral populations. Cats are efficient predators on vertebrates that have had no prior experience of them; for example, cats have caused the extinction of some 30 island bird species worldwide. Introduced herbivores, particularly goats, devastate native plant communities, again because the plants have had no chance to adapt to these new threats.
Although islands are particularly vulnerable, introduced species also wreak large-scale changes on continents. An exceptionally clear example is the loss of the American chestnut (Castanea dentata) in eastern North America after the accidental introduction of the fungus, Endothia parasitica, that causes chestnut blight. The chestnut, a once-abundant tree, was removed with surgical precision over its entire range beginning in the early 20th century. All that survive are small individuals that tend to become infected as they get older.
As briefly mentioned above, hybridization is another mechanism by which introduced species can cause extinction. In general, species are considered to be genetically isolated from one another—they cannot interbreed to produce fertile young. In practice, however, the introduction of a species into an area outside its range sometimes leads to interbreeding of species that would not normally meet. If the resident species is extremely rare, it may be genetically swamped by the more abundant alien. One example of a species threatened by hybridization is the white-headed duck (Oxyura leucocephala; see stifftail). The European population of this species lives only in Spain, where habitat destruction and hunting once reduced it to just 22 birds. With protection, it recovered to about 800 individuals, but it is now threatened by a related species, the ruddy duck (O. jamaicensis). This bird is native to North America, was introduced to Great Britain in 1949, and spread to the continent including Spain, where it hybridizes with the much rarer white-headed duck. Similar cases involve other species of ducks, frogs, fish, cats, wolves, and other groups of organisms worldwide.
Overharvesting
Overharvesting, or overfishing in the case of fish and marine invertebrates, depletes some species to very low numbers and drives others to extinction. In practical terms, it reduces valuable living resources to such low levels that their exploitation is no longer sustainable. Whereas the most-familiar cases involve whales and fisheries, species of trees and other plants, especially those valued for their wood or for medicines, also can be exterminated in this way.
whaling
 whaling offers an example of overharvesting that is interesting not only in itself but also for demonstrating how poorly biodiversity has been protected even when it is of economic value. The first whalers likely took their prey close to shore. right whales were the “right” whales to take because they are large and slow-moving, feed near the surface and often inshore, float to the surface when harpooned, and were of considerable commercial value for their oil and baleen (see whalebone). The southern right whale (Eubalaena australis), for example, is often seen in shallow, sheltered bays in South Africa and elsewhere. Such behaviour would make any large supply of raw materials a most tempting target. Whalers had nearly exterminated the North Atlantic species of the northern right whale (Eubalaena glacialis) and the bowhead whale (Greenland right whale; Balaena mysticetus) by 1800. They succeeded in exterminating the Atlantic population of the gray whale (Eschrichtius robustus). Whalers then moved on to species that were more difficult to kill, such as the humpback whale (Megaptera novaeangliae) and the sperm whale (Physeter macrocephalus).
whaling offers an example of overharvesting that is interesting not only in itself but also for demonstrating how poorly biodiversity has been protected even when it is of economic value. The first whalers likely took their prey close to shore. right whales were the “right” whales to take because they are large and slow-moving, feed near the surface and often inshore, float to the surface when harpooned, and were of considerable commercial value for their oil and baleen (see whalebone). The southern right whale (Eubalaena australis), for example, is often seen in shallow, sheltered bays in South Africa and elsewhere. Such behaviour would make any large supply of raw materials a most tempting target. Whalers had nearly exterminated the North Atlantic species of the northern right whale (Eubalaena glacialis) and the bowhead whale (Greenland right whale; Balaena mysticetus) by 1800. They succeeded in exterminating the Atlantic population of the gray whale (Eschrichtius robustus). Whalers then moved on to species that were more difficult to kill, such as the humpback whale (Megaptera novaeangliae) and the sperm whale (Physeter macrocephalus).The Napoleonic Wars gave whales a respite, but with the peace of 1815 came a surge of whalers into the Pacific Ocean, inspired by the stories of James Cook and other explorers. The first whalers arrived in the Hawaiian Islands in 1820, and by 1846 the fleet had grown to nearly 600 ships, the majority from New England. The catch on each whaling voyage averaged 100 whales, though a voyage could last as long as four years.
In the late 1800s, steamships replaced sailing ships, and gun-launched exploding harpoons replaced hand-thrown lances. The new technology allowed whalers to kill what until then had been the “wrong” whales—fast-swimming species such as the blue whale (Balaenoptera musculus) and fin whale (B. physalus). Whalers killed nearly 30,000 blue whales in 1931 alone; World War II gave the whales a break, but the catch of blue whales rose to 10,000 in 1947. The fin whale was next, with the annual catch peaking at 25,000 in the early 1960s; then came the smaller sei whale (B. borealis)—which no one had bothered to kill until the late 1950s—and finally the even smaller minke whale (B. acutorostrata), which whalers still hunt despite an international moratorium in effect since 1986 that seeks to curb commercial whaling.
The story of whaling is, in brief, the rapid depletion and sometimes extermination of one population after another, starting with the easiest species to kill and progressing to the most difficult. That whales are economically valuable raises the obvious question of why there were no attempts to harvest whales sustainably.
Fishing
Overfishing is the greatest threat to the biodiversity of the world's oceans, and contemporary information published for fisheries in the United States can serve as an example of the magnitude of the problem. Congress requires the National Marine Fisheries Service (NMFS) to report regularly on the status of all fisheries whose major stocks are within the country's exclusive economic zone, or EEZ. (Beyond its territorial waters, every coastal country may establish an EEZ extending 370 km 【200 nautical miles】 from shore. Within the EEZ the coastal state has the right to exploit and regulate fisheries and carry out various other activities to its benefit.) The areas involved are considerable, covering portions of the Atlantic, the Caribbean, the Gulf of Mexico, and the Pacific from off San Diego to the Bering Sea out to the west of the Hawaiian island chain along with the islands constituting the western part of the former Trust Territory of the Pacific Islands (Pacific Islands, Trust Territory of the). At the turn of the 21st century, the NMFS deemed some 100 fish stocks to be overfished and a few others close to being so, while some 130 stocks were not thought to be overfished. For about another 670 fish stocks, the data were insufficient to allow conclusions. Thus, a little under half of the stocks that could be assessed were considered overfished. For the major fisheries—those in the Atlantic, the Pacific, and the Gulf of Mexico—two-thirds of the stocks were overfished.
As to the hundreds of stocks about which fisheries biologists know too little, most of them are not considered economically important enough to warrant more investigation. One species, the barn-door skate (Raja laevis), was an incidental catch of western North Atlantic fisheries in the second half of the 20th century. As the name suggests, this is a large fish, too big to go unrecorded. Its numbers fell every year until, by the 1990s, none were being caught.
logging and collecting
Similar cases of overharvested species are found in terrestrial ecosystems. For example, even when forests are not completely cleared, particularly valuable trees such as mahogany may be selectively logged from an area, eliminating both the tree species and all the animals that depend on it. Another example is the coast sandalwood (Santalum ellipticum), a tree endemic to the Hawaiian Islands that was almost completely eliminated from its habitats for its wood and fragrant oil.
Some species are overharvested not to be killed but to be kept alive and sold as pets or ornamental plants. Many species of parrots worldwide, for example, are in danger because of the pet trade, and the survival of cacti (cactus) and orchid species is threatened by collectors.
Secondary extinctions
Once one species goes extinct, there will likely be other extinctions or even an avalanche of them. Some cases of these secondary extinctions are simple to understand—e.g., for every bird or mammal that goes extinct, one or several species of parasite (parasitism) also will likely disappear. From well-studied species, it is known that bird and mammal species tend to have parasites on or inside them that can live on no other host.
 Other extinctions cause changes that can be quite complicated. Species are bound together in ecological communities to form a food web (see food chain) of species interactions. Once a species is lost, those species that fed on it, were fed on by it, or were otherwise benefited or harmed by that species will all be affected, for better or worse. These species, in turn, will affect yet other species. Ecological theory suggests that the patterns of secondary extinction are quite complicated and thus may be difficult to demonstrate.
Other extinctions cause changes that can be quite complicated. Species are bound together in ecological communities to form a food web (see food chain) of species interactions. Once a species is lost, those species that fed on it, were fed on by it, or were otherwise benefited or harmed by that species will all be affected, for better or worse. These species, in turn, will affect yet other species. Ecological theory suggests that the patterns of secondary extinction are quite complicated and thus may be difficult to demonstrate. The most easily recognizable secondary extinctions should be seen in species that depend closely on each other. In the Hawaiian Islands, anecdotal evidence for secondary extinctions comes from a consideration of nectar-feeding birds. Before modern human activity on the islands, there were three nectar-feeding Hawaiian honeycreepers—the mamo (Drepanis pacifica), the black mamo (Drepanis funerea), and the iiwi (Vestiaria coccinea)—that had long decurved (downward-curving) beaks, the kind adapted to inserting into appropriately long and curved flowers. The first two birds are extinct, whereas the third is extinct on two islands, is very rare on a third, and has declined on others.
The most easily recognizable secondary extinctions should be seen in species that depend closely on each other. In the Hawaiian Islands, anecdotal evidence for secondary extinctions comes from a consideration of nectar-feeding birds. Before modern human activity on the islands, there were three nectar-feeding Hawaiian honeycreepers—the mamo (Drepanis pacifica), the black mamo (Drepanis funerea), and the iiwi (Vestiaria coccinea)—that had long decurved (downward-curving) beaks, the kind adapted to inserting into appropriately long and curved flowers. The first two birds are extinct, whereas the third is extinct on two islands, is very rare on a third, and has declined on others.Extensive habitat destruction is likely the cause of many species of Hawaiian birds, as previously discussed. In addition, native Hawaiians hunted some species for their feathers. In the case of the three honeycreepers described above, however, their extinctions may have followed the destruction of important nectar-producing plants by introduced goats and pigs. Many of the native lobelias, such as those of the genera Trematolobelia and Clermontia, have clearly evolved to be pollinated by the three honeycreepers, and the plants were once important components of the forest's understory. About a quarter of these plant species are now extinct, a rate that plainly exceeds those for the rest of the flora, perhaps because they were so vulnerable to introduced mammalian herbivores. It is not certain, however, whether the plants disappeared first and then their bird pollinators or vice versa.
Similarly, some surviving Hawaiian birds seem to be unusually specialized feeders and to be threatened as a consequence of the loss of their food sources. For example, another rare honeycreeper, the akiapolaau (Hemignathus munroi), is an insectivore that feeds on insects mainly on large koa (Acacia koa) trees (see acacia). Today, however, few koa forests remain, because the trees have been overharvested for their attractive wood. Yet another Hawaiian honeycreeper, a seed-eating species called the palila (Loxioides bailleui), is endangered because it depends almost exclusively on the seeds of one tree, the mamane (Sophora chrysophylla), which is grazed by introduced goats and sheep.
Stories of secondary extinctions are nearly always unsatisfactory anecdotes because of the difficulty of teasing apart the various explanations for what happened in the past. There is abundant evidence from small-scale ecological experiments that a change in one species' numbers (including its complete elimination) will cause cascading effects in the abundance of other species. The stories are plausible enough that particular attention should be paid to the future consequences of contemporary extinctions.
Global change
 The global effects of flooding the atmosphere with carbon dioxide and other greenhouse gases (see greenhouse effect) created as by-products of human activity are many and complex. global warming, an increase in global average surface temperature, is but one of them. Both the land and the oceans are increasing in temperature at different rates in different places. The Arctic and Antarctic seem to be heating up the most, while temperature changes in the tropics are more modest. In addition, the heating is melting glaciers and ice sheets, causing sea levels to rise, and it may be increasing the frequency of the most intense hurricanes and perhaps the intensity of other storms and extreme weather, with consequences to the flow of rivers. Some areas and their ecosystems are becoming wetter, while others are becoming drier and thus likely to suffer more wildfires; some areas may even become colder. Scientific knowledge is constantly increasing and changing what is known about the way Earth works, and knowledge of global change and its effects on species is expanding particularly quickly.
The global effects of flooding the atmosphere with carbon dioxide and other greenhouse gases (see greenhouse effect) created as by-products of human activity are many and complex. global warming, an increase in global average surface temperature, is but one of them. Both the land and the oceans are increasing in temperature at different rates in different places. The Arctic and Antarctic seem to be heating up the most, while temperature changes in the tropics are more modest. In addition, the heating is melting glaciers and ice sheets, causing sea levels to rise, and it may be increasing the frequency of the most intense hurricanes and perhaps the intensity of other storms and extreme weather, with consequences to the flow of rivers. Some areas and their ecosystems are becoming wetter, while others are becoming drier and thus likely to suffer more wildfires; some areas may even become colder. Scientific knowledge is constantly increasing and changing what is known about the way Earth works, and knowledge of global change and its effects on species is expanding particularly quickly.Since the turn of the 21st century, the relationship between changes to Earth's physics and chemistry and biodiversity has been clarified significantly. Although the precise effects of global change on species extinction rates are still uncertain, they almost certainly will be large.
It is now clear that most species are shifting their geographic ranges toward cooler places and are starting important events such as breeding, migration, and flowering earlier in the year. One study, published in the early 21st century, found that more than 80 percent of nearly 1,500 species of animals and plants from a wide variety of habitats worldwide were changing in the direction expected from global warming. Another study, published about the same time, found consistent northward movement of the northern boundaries of animal and plant species in Europe and North America. Such geographic shifts of species alter important ecological interactions with their prey, predators, competitors, and diseases. Some species can benefit; others can be harmed—for example, when migratory insect-eating birds arrive too late to exploit the emergence of moth larvae that they typically feed to their young.
Even if scientists know that species likely will move in the direction of cooler habitats—toward the poles or up mountainsides—they will find the exact changes difficult to predict because so many different factors are involved. For example, one contemporary study examined the changes in the ranges and feeding ecology of butterflies (butterfly) in Great Britain in the last decades of the 20th century. Two species were found to have increased in range, as expected, but by unexpected amounts. One species lived in isolated habitat patches and could not fly the distance to some of the patches that were suitable for populating. Rising temperatures increased the number and density of suitable patches, allowing the butterfly to reach the distant patches by making use of intervening newly available ones as “stepping-stones.” The second butterfly species was able to exploit a previously unused food plant that grew in shady places that formerly had been too cool. With that change in diet, the butterfly was able to greatly expand its range. In both cases, the species benefited from the warming climate, at least at the northern edges of their ranges. It takes little imagination to foresee the consequences if, for example, the mosquito Aedes aegypti, which carries dengue hemorrhagic fever, were to expand its range to an unexpected degree across the southern United States, which is its present northern limit.
Can anything useful be inferred about the likely consequences of climate change to species extinction? Assuming that the change has simple effects, scientists can predict where a species should be in the future if it is to live in the same range of climatic conditions that it does now. The real concern is that this range of suitable conditions, or the species' climate envelope, may shrink to nothing as conditions change—i.e., there may be no suitable conditions for a species in the future.
Other things being equal, one would expect that species with small geographic ranges will more likely be affected than species with large ranges. For a species with a large range, global change may cause the species to disappear from the south and appear farther north, but the change in the size of the range may be quite small. The species may be present in many of the same areas both before and after global warming. The American robin, whose large range size is discussed above, could be expected to behave this way.
 Of the species that have very small ranges, many live in mountainous areas, as can be seen in the map-->. To date, human activity has had relatively small effects on such species because of the impracticalities of cultivating land in mountains, especially on steep slopes. But it is these species—living on cool mountaintops that are now becoming too warm for them—that have nowhere else to go. Such arguments lead scientists to believe that extinctions caused by rising temperatures will be additional to those caused by land-use changes such as deforestation. Rough calculations suggest that rising temperatures may threaten about a quarter of the species in hot spots.
Of the species that have very small ranges, many live in mountainous areas, as can be seen in the map-->. To date, human activity has had relatively small effects on such species because of the impracticalities of cultivating land in mountains, especially on steep slopes. But it is these species—living on cool mountaintops that are now becoming too warm for them—that have nowhere else to go. Such arguments lead scientists to believe that extinctions caused by rising temperatures will be additional to those caused by land-use changes such as deforestation. Rough calculations suggest that rising temperatures may threaten about a quarter of the species in hot spots.Which species are most vulnerable to extinction?
Before a species becomes extinct, it must first be rare. Some species are naturally rare, while others have that rarity imposed upon them by one or more of the factors discussed above. Some species are much more vulnerable than others to becoming dangerously rare, and other species, when rare, are more likely than others of equal rarity to succumb. These and other factors affecting a species' susceptibility to destruction are discussed in this section.
Endemism and rarity
As previously discussed, a small geographic range makes a species particularly vulnerable to global extinction. Many of the threats to species are geographically restricted, so species with large ranges will survive somewhere even if they are locally extirpated. Species with small ranges do not have this “reserve.” Moreover, as also was mentioned above, species with small geographic ranges tend to be rare within them—they have low population densities.
The collections of plants and animals on islands are often rich in endemic species; those species that are restricted to particular islands must inevitably have small ranges. Since 1600 the great majority of known bird extinctions have been on islands. The problem of extinction is not just limited to islands, however, as the previous sections amply describe.
In sum, combined endemism and rarity are the factors that confer the greatest risk of extinction.
Body size and related reproductive characteristics
 Generally, the larger the body size of an animal, the longer it lives and the fewer offspring it produces each year. Relatively large animals also tend to have relatively low population densities; thus, a viable population of, say, elephants occupies considerably more space than an equal-sized population of rabbits. Large predators such as tigers (Panthera tigris) have lower population densities than the herbivores on which they feed. A tiger has a home range that may occupy 100 square km (40 square miles), while a rabbit may survive in 1 hectare (0.004 square mile).
Generally, the larger the body size of an animal, the longer it lives and the fewer offspring it produces each year. Relatively large animals also tend to have relatively low population densities; thus, a viable population of, say, elephants occupies considerably more space than an equal-sized population of rabbits. Large predators such as tigers (Panthera tigris) have lower population densities than the herbivores on which they feed. A tiger has a home range that may occupy 100 square km (40 square miles), while a rabbit may survive in 1 hectare (0.004 square mile).Large animals, by virtue of their low population densities, are at increased risk of extinction. Moreover, an animal species that produces few offspring each year and that suffers a major loss in numbers from human activity will need more time to recover than a species with high reproductive rates. Again taking the tiger as an example, a tiger population large enough to avoid all the perils of low population size (including the chance that all offspring will be the same sex or that genetic inbreeding will occur) requires extremely large areas. Although the tiger is not considered an endemic species—it has a very large geographic range, stretching over much of eastern and southern Asia—its rarity in that range means that it is nonetheless vulnerable to extinction from human activity.
Behaviour (animal behaviour)
Size of home range
In the case of lions, wolves, and wild dogs, population size alone is a poor predictor of their local extinction, even when the animals live in protected areas. For these species, conflict with people on the borders of the protected areas is the major cause of mortality. Border areas are population “sinks,” areas where the death rate of the population exceeds the birth rate and into which individuals enter through migration from more-central portions of the protected area. This explains, for example, why African wild dogs (African hunting dog) (Lycaon pictus; see African hunting dog) are under particular threat—more so than other species, such as cheetahs and leopards, that also are afforded protected areas. Wild dogs live in large packs that roam very large areas, in contrast to the two cat species, which are mostly solitary and which have smaller home ranges. The more widely a species moves (i.e., the larger its home range), the more likely it is to move beyond areas where it is protected.
Concentration
Some species have aggregation behaviours that make them vulnerable to disturbance or hunting. For example, bats may congregate in large numbers in particular caves to have their young, making significant portions of their total population especially susceptible when their habitat is disturbed by human visitation or damaged by the cave's commercialization or flooding. The various species of groupers often come together to spawn on a few nights of each year tied to phases of the moon and at traditional mating sites. Fishermen who know these sites and the timing of spawning can devastate large populations of these species by concentrating their efforts during this most vulnerable time in the fish's life cycle.
Low dispersal
Because small populations are so much more at risk than large ones, individuals of species that can readily disperse can rescue local populations on the verge of extinction. The previously discussed example of the checkerspot butterfly (see above Surviving but threatened small populations (conservation)) illustrates, first, the rescue of some small populations by the dispersal of individuals from larger “reservoir” populations nearby and, second, the subsequent extinction of the reservoirs as urban development isolated them from still other butterfly populations that could have rescued them. Species that have low dispersal rates are at a disadvantage because it is unlikely that one population can save another.
Mating systems
Small populations suffer from inbreeding, an inevitable tendency of mating individuals in a small isolated population to be more closely related than they would be in a larger one. When population size is severely reduced, inbreeding may be the final insult that will cause the remaining population to go extinct. The likelihood that this will happen, however, seems related to the social structure of the species involved.
 A comparison of two species, a seal and a rhinoceros, serves to explore the issue in more detail. The northern elephant seal (Mirounga angustirostris) of the Pacific Coast of North America was thought to have been hunted to extinction in the late 1800s, though it later became apparent that perhaps 20–30 individuals persisted locally for a couple of decades before the population began to recover gradually under protection. The Indian rhinoceros (Rhinoceros unicornis) in the early 20th century was reduced to two isolated populations—one numbering between 12 and 100, the other between 60 and 80—before protection allowed it to make a limited recovery. Moreover, not all of the rhinoceros males in the reduced population were likely to have bred. Today the elephant seal is genetically uniform, suggesting that a high degree of inbreeding occurred during the time its population was at a minimum, whereas the rhino has probably lost little of its genetic variability. The population histories of the two species are similar, so why the differences in their genetic variability?
A comparison of two species, a seal and a rhinoceros, serves to explore the issue in more detail. The northern elephant seal (Mirounga angustirostris) of the Pacific Coast of North America was thought to have been hunted to extinction in the late 1800s, though it later became apparent that perhaps 20–30 individuals persisted locally for a couple of decades before the population began to recover gradually under protection. The Indian rhinoceros (Rhinoceros unicornis) in the early 20th century was reduced to two isolated populations—one numbering between 12 and 100, the other between 60 and 80—before protection allowed it to make a limited recovery. Moreover, not all of the rhinoceros males in the reduced population were likely to have bred. Today the elephant seal is genetically uniform, suggesting that a high degree of inbreeding occurred during the time its population was at a minimum, whereas the rhino has probably lost little of its genetic variability. The population histories of the two species are similar, so why the differences in their genetic variability?The social structures of elephant seals and rhinos are dramatically different. Each year, the one dominant bull seal that guards the harem is likely to father all of the young. An isolated seal population thus may become genetically uniform relatively quickly because very few males father each generation of young. Rhinos, on the other hand, are largely monogamous, so a group of them will have a greater number of fathers than a comparably sized group of seals.
Insularity
The vulnerability of island species is likely a combination of two factors previously discussed—their endemism and rarity and their ecological naivete, the latter being exemplified by the greater effect of domestic cat introductions on unwary island bird species than on more “streetwise” mainland species. Nevertheless, some island bird species are less likely to be threatened than similar bird species found on continents. The reason lies in the abundance of island species—they are often quite numerous on their islands, for they have fewer competitors than do mainland species.
Human use
Many species are hunted for meat and other products, including whales and various fish, as discussed above. Less familiar is the widespread trade in bushmeat, which is essentially everything that can be hunted—from mice (mouse) to chimpanzees and gorillas—and is especially prevalent in West and Central Africa. Yet other species are harvested for body parts, such as tiger bones and rhino horns, which are used in Asian medicines. A wide variety of plants are harvested too, again often for medicinal purposes. Simply put, any species that is used for food, wood, or medicine or as pets or houseplants, that is collected (such as butterflies or invertebrate shells), or that attracts attention for any other reason suffers an increased risk of extinction.
Preventing the loss of biodiversity
A thorough knowledge of the factors that cause extinction and the vulnerability of different species to them is an essential part of conserving species. In large part, conservation is about removing or reducing those factors and doing so for the most vulnerable species and in the places where species are most vulnerable. Much of the task of conservation professionals is to protect habitats large enough to house viable populations of species, first deciding where the priorities should be and sometimes restoring habitats that already have been destroyed. Local conservation groups often spend time removing introduced species, which can mean physically weeding invasive plants or trapping invasive animals. These activities must be accompanied by efforts to prevent introductions of new threats. Others work to reduce harvesting directly or to reduce the incidental catch of nontarget species. Nonetheless, there are a variety of specific tools that can be applied to different circumstances, as categorized below and illustrated by case histories. Sometimes, when working with very rare species, scientists may not know the exact causes of threat, which can lead to intense arguments about exactly how to proceed.
Species interventions
Protective custody
Some species become so rare that there are doubts about whether they will be able to survive in the wild. Under such circumstances, the species may be brought into protective custody until areas can be made suitable for their release back into the wild.
Protective custody is an important tool in plant conservation, where a large number of seeds can be easily stored. In addition, botanic gardens can grow rare plants, protecting the species until such times as they can be planted in the wild. Of a total of about 300,000 described species of flowering plants, Botanic Gardens Conservation International has estimated that about 80,000 species are protected in botanic gardens and a few thousand additional species in other facilities—together about one-third of the total.
For animals, zoos provide an important refuge for some vertebrate species but not for the vast majority of animals, which are invertebrates. Of species protected in zoos, a number of them have later been returned to the wild. To do so, however, substantial problems need to be overcome.
Determining that a species should be brought into captivity and then deciding what to do with the individuals are illustrated by the California condor. The first key decision was whether to bring the birds into protective custody or to manage their small population in the wild. There was no dispute that the condors were once widespread, ranging across the southern and western United States and northern Mexico. As long ago as the turn of the 20th century, however, they were restricted to the mountains of southern California. The widespread practice of setting out poisoned carcasses to kill livestock predators was likely the major cause of their decline. Exactly how many condors still survived was at the core of the debate. Some thought their numbers had been declining constantly, from 150 in the 1950s to 60 in 1970. Others posited constant numbers, and, according to one opinion, if there was no decline, there was need for neither explanation nor intervention—the condors, though rare, should be left alone.Eventually, photographic surveys completed the catalog of individuals, removing doubts about which individuals were alive, which were dead, and why. Studies of nesting showed that those birds that bred did so with reasonable success. Radio-collared birds showed that the species foraged far beyond the remote areas of its nesting sites, so simply leaving the birds alone and protecting the habitat near the nest would not be sufficient. Birds died from ingesting lead in animals that had been shot or the toxic substances in poison-laced carrion. In the 1980s all the remaining birds were brought into captivity, although not without a lawsuit over the issue.
Because captive populations are almost always small, there is a high risk of inbreeding. Thus, the condors were carefully screened genetically to ensure that as much as possible of the genetic variability of the species was preserved (see below). The captive-breeding program was ultimately a success, and between 12 and 20 fledglings were produced each year after 1991. Earlier production had been lower, likely as a consequence of the birds' inexperience in nesting and rearing. Efforts to teach captive-reared birds through artificial means—some nestlings were fed by puppets resembling condor heads—were much less successful. These birds were particularly inept when released, as real parents teach their young many important things about living in the wild. Eventually, scientists released birds in Arizona, California, and Baja California, and some of these birds have reared young of their own in the wild.
Genetic intervention
In small populations, inbreeding can cause genetic (genetics) variability to be lost quite quickly. A simple example is provided by the Y chromosome in humans (and other mammals), which confers maleness and which behaves like human surnames do in large parts of the world. If every human couple had just two children each generation, then by chance alone 25 percent of the couples would have two sons, each with one Y chromosome from the father; 25 percent would have two daughters, each with no Y chromosome; and the remaining 50 percent would have one son and one daughter. The surnames of the fathers with two daughters would be lost as they married and had their own children, as indeed would be those fathers' Y chromosomes. In a small breeding population, after just a few generations, every individual would have the last name of the same male ancestor—perhaps a great-great-grandfather—and the same Y chromosome.
Many genetic mutations are deleterious, reducing the individual's chances of survival, but are also recessive (see recessiveness), requiring the inheritance of one mutant gene from each parent to manifest their effect. This means that in a large population, their effect will be masked by the overwhelming numerical superiority of the normal dominant gene ( allele; see dominance). For the reasons explained above, in a small population chance events cause the loss of genetic variability and so increase the likelihood that individuals will suffer harmful genetic diseases.
Studies on mammals kept in zoos illustrates the harmful effects of inbreeding. In the past, to maintain sufficient productivity, zookeepers often bred animals that proved to be good at producing young. Because of this practice, some breeding pairs quickly came to have the same grandparents and, in some cases, the same parents. The studies showed that such pairs produced young that were much less likely to survive than young from pairs of unrelated individuals.
The practical problem for conservation is whether to place efforts on genetic intervention—bringing in “new blood,” that is, individuals and so genes, from the outside—or to concentrate on the factors causing the initial decline.
 This issue was at the heart of the management dilemma posed by the Florida panther (Puma concolor coryi), a distinct subspecies of puma (P. concolor) confined to a small, isolated, and inbred population in southern Florida. The specific question was whether to introduce pumas from Texas into the Florida population. Florida panthers once had been part of a continuous widespread population. In the 19th century they became isolated in southern Florida. As their numbers declined, the occurrence of genetic defects increased, including sperm and heart defects and undescended testicles in males. Conservation scientists hoped that the introduction of pumas from outside Florida would reverse the genetic damage. This proposal was highly controversial; as with the example of the California condor, some argued that the population was doing well in its limited range and did not need additional animals. Despite concerns, in the mid-1990s eight females from Texas were released, and scientists closely followed the fates of them, of their young, and of the young of cats with only Florida parents. It was found that, although hybrid cats—those with both Florida and Texas parents—do not seem to live longer than pure Florida cats and hybrid females do not produce more kittens than pure Florida females, hybrid kittens survive about twice as well. Since the introduction of the Texas females, numbers of Florida panthers have increased, and hybrid cats were expanding the known range of their habitats.
This issue was at the heart of the management dilemma posed by the Florida panther (Puma concolor coryi), a distinct subspecies of puma (P. concolor) confined to a small, isolated, and inbred population in southern Florida. The specific question was whether to introduce pumas from Texas into the Florida population. Florida panthers once had been part of a continuous widespread population. In the 19th century they became isolated in southern Florida. As their numbers declined, the occurrence of genetic defects increased, including sperm and heart defects and undescended testicles in males. Conservation scientists hoped that the introduction of pumas from outside Florida would reverse the genetic damage. This proposal was highly controversial; as with the example of the California condor, some argued that the population was doing well in its limited range and did not need additional animals. Despite concerns, in the mid-1990s eight females from Texas were released, and scientists closely followed the fates of them, of their young, and of the young of cats with only Florida parents. It was found that, although hybrid cats—those with both Florida and Texas parents—do not seem to live longer than pure Florida cats and hybrid females do not produce more kittens than pure Florida females, hybrid kittens survive about twice as well. Since the introduction of the Texas females, numbers of Florida panthers have increased, and hybrid cats were expanding the known range of their habitats.Associated difficulties and costs
Previous discussions have covered the release of individuals into the wild to reestablish populations that have gone extinct or to rescue inbred populations. They also have noted that protecting species from human harvesting and removing invasive species are major techniques in saving biodiversity. These and other species-by-species interventions are often high-profile actions that attract considerable media attention. This does not mean that they are easy or cheap, as the following examples demonstrate.
Protecting species
 For species that are hunted or collected, direct protection may be an essential conservation tool. National laws, such as the Endangered Species Act in the United States, make collecting or killing an endangered species or threatened species illegal. An example of such a protected species in the United States is the country's national bird, the bald eagle (Haliaeetus leucocephalus). International laws protect whales of various species, and such agreements as the Convention on International Trade in Endangered Species prohibit commercial trade in designated species. Enforcing these laws and conventions is another matter, however.
For species that are hunted or collected, direct protection may be an essential conservation tool. National laws, such as the Endangered Species Act in the United States, make collecting or killing an endangered species or threatened species illegal. An example of such a protected species in the United States is the country's national bird, the bald eagle (Haliaeetus leucocephalus). International laws protect whales of various species, and such agreements as the Convention on International Trade in Endangered Species prohibit commercial trade in designated species. Enforcing these laws and conventions is another matter, however.
 The problems of implementing protection are illustrated by the conservation of the two African species of rhinoceros. The population of the black rhinoceros (Diceros bicornis) fell to about 2,400 individuals in 1995, down from a likely number of several hundred thousand at the start of the 20th century, when it ranged over most of southern Africa. The white rhinoceros (Ceratotherium simum) historically had a smaller geographic range. Today its northern subspecies occurs only in the Democratic Republic of the Congo, where it is very rare. The southern subspecies lives almost entirely in South Africa, Namibia, Zimbabwe, and Kenya. Together, they numbered under 12,000 in 2001, again likely a small fraction of their original numbers.
The problems of implementing protection are illustrated by the conservation of the two African species of rhinoceros. The population of the black rhinoceros (Diceros bicornis) fell to about 2,400 individuals in 1995, down from a likely number of several hundred thousand at the start of the 20th century, when it ranged over most of southern Africa. The white rhinoceros (Ceratotherium simum) historically had a smaller geographic range. Today its northern subspecies occurs only in the Democratic Republic of the Congo, where it is very rare. The southern subspecies lives almost entirely in South Africa, Namibia, Zimbabwe, and Kenya. Together, they numbered under 12,000 in 2001, again likely a small fraction of their original numbers.Although conversion of habitat land to agricultural use and sport hunting caused the earlier rhino declines, the major threat is now poaching—entering reserves where the animals are protected and killing them for their horns, which are in high demand in parts of the world for dagger handles and, in powdered form, as an ingredient of traditional medicines. The costs of protecting rhinos are considerable. WWF has estimated that it costs $1,400 per square km ($3,600 per square mile) per year to detect and deter poaching. Reserves that hold rhinos often cover tens of thousands of square kilometres; the multimillion-dollar budgets required are beyond the means of many African governments.
Removing invasive species
Many conservation programs have tackled invasive species, and, once again, the message from the two case studies that follow is that, although these programs can be successful, they are often expensive.
As previously described, introduced domestic cats have caused the extinction of many island species. In the 1970s scientists estimated that the cat population on Marion Island, one of the two Prince Edward Islands in the subantarctic Indian Ocean, was killing 450,000 seabirds each year, jeopardizing the birds' survival. The cats, which were estimated to number at least 2,000, were descended from five animals brought to the island in 1949 to deal with a mouse problem at a meteorological station there. Islands are small and isolated, and because cats are predators, they tend to have much smaller populations than their prey. Nonetheless, it took 19 years of an intensive eradication program to remove the last cats.
The second case study is the purple loosestrife (Lythrum salicaria), a plant that has overrun thousands of square kilometres of North American wetlands, replacing the naturally diverse vegetation of grasses, sedges, and other wetland plants. It is native to Europe and was introduced into North America in the early 1800s. It now occurs across most of the continental United Sates and is most prevalent in the northeastern and north-central part of the country and in Canada.
Like other exotic weeds, purple loosestrife can spread rapidly. Control measures have included the use of herbicides and the release of herbivorous insects, including different species of beetles that eat the leaves, roots, and seeds of the plant. The latter strategy, an instance of biological control, is not risk-free. It involves introducing a second nonnative species to control a nonnative species introduced earlier. The effects of the insect introductions have not always been clear, and, as with other examples of biological control, there is a danger that the introduced herbivorous insects will harm native plant species.
Introducing species (introduced species)
The introductions of Texas pumas into the Florida panther population and of captive-reared condors back into parts of their original habitat were successful, as discussed above. So were the introductions to North America of the starling, also discussed above, and the house sparrow (Passer domesticus), which was introduced to New York City from Europe in the 1850s. What is often overlooked, however, is that many other attempts have failed. This in fact is the typical result.
For example, after extensive efforts in the 19th and early 20th centuries to introduce nonnative game birds to the United States, it was found that, even with releases of large numbers of birds, most of the attempts failed. An exception has been the common, or ring-necked, pheasant (Phasianus colchicus), native to China and introduced to the United States in the 1890s. This low rate of success has an important implication—even when it is known from hindsight that an individual introduction can succeed, as did the pheasant, most such introductions still will fail. When returning species to the wild that have already gone extinct there, the prognosis is even bleaker. One reason in some cases is that the cause of the extinctions—for instance, the brown tree snake responsible for bird extinctions on Guam—is still present.
habitat protection
Because the loss of habitat is the primary reason that species are lost both locally and globally, protecting more habitat emerges as the most important priority for conservation. This simple idea raises difficult questions. Which habitats should be protected? And because it seems unlikely that all habitats can be protected, which ones should receive priority?
If reserves (nature reserve) were judiciously placed over the identified hot spots of biodiversity, the special places where vulnerable species are concentrated, a large fraction of species might be saved. Presently, the allocation of reserves around the world is poor. Reserves larger than 100,000 square km (40,000 square miles) are generally in high mountains, tundras, and the driest deserts, areas that are not particularly species-rich. On the other hand, hot spots such as Madagascar and the Philippines protect less than 2 percent of their land.
The same kinds of questions hold on smaller scales, as illustrated by a study reported in the late 1990s. The Agulhas Plain at the southern tip of Africa is one of the world's “hottest” spots for concentrations of vulnerable plant species. An area only some 1,500 square km (600 square miles) in size was found to house 1,751 plant species, 99 of them endemic. Whereas most of the state forests and private nature reserves in the area are coastal, most of the hot spot's endemic plants live inland. Given that new reserves must be created if these plants are to survive, where should they be situated to encompass the maximum number of species at minimum cost?
Fortunately, the data available to make these decisions included a knowledge of the distribution of plant species over the Agulhas Plain in fairly good detail—the kind of information not likely to be available in most hot spots. This allowed the plain's plant-species composition to be divided into a grid of cells, each 3 × 3 km on a side. Computer algorithms (systematic problem-solving methods) were then used to select sets of cells from the grid according to their complementary species composition—that is, the aim was to encompass as many species or as many endemics as possible in as small an area (as few grid cells) as possible.
Naively applied, these algorithms will not give useful results. For example, the sites they select may not be available for reserves. Also, the choice of too small a cell size can lead to the selection of protected areas containing populations so small and widely scattered that they would be unlikely to persist. This is fittingly dubbed the “Noah's Ark effect,” because the ark held only two individuals of each species for a short time. Reserves (nature reserve) need to be large enough to support species indefinitely. The choice of a cell size of 3 × 3 km is politically feasible because reserves of this size already have been established in the region and are probably ecologically sensible for many plant species. Other factors had to be included in the final selection of cells. Some areas are unsuitable for various reasons—for instance, some are overrun by alien weeds, while others are mostly in urban areas or croplands. In contrast, other areas are particularly desirable—for example, they may be adjacent to existing reserves, and it is easier to expand such reserves than to create new ones. The results of this study thus provided advice for establishing reserves that combined ecological information on species distribution with practical and political considerations.
Saving the most species for the least money likewise was the consideration that motivated another study published in the late 1990s, of which counties in the United States should be conservation priorities. An earlier study that attempted to locate sites for new reserves in the United States had equated efficiency with the minimum number of counties needed to achieve a given coverage of endangered species. That approach would have been sensible if land were much the same price everywhere. Unfortunately, the study's targets had included counties encompassing San Diego, Santa Cruz, and San Francisco in California, Honolulu in Hawaii, and certain counties in Florida, all of which are among the highest-priced land in the country. The later study asked how many species could be protected for a given total cost. It found that considerable savings in cost per species accrue from selecting larger, more-complementary areas and lower total costs and that, as a consequence of this approach, the places identified for protection were often quite different from those recommended in the earlier study.
Habitat connections
Habitats that are not completely destroyed may be fragmented to the degree that individual fragments are too small to hold viable populations of many species—they may suffer from inbreeding or the increased demographic risks previously discussed. Yet in total the fragments may actually be of sufficient area to support these species. An obvious conservation intervention is to find ways of connecting fragments by corridors. These corridors can be created from currently unprotected land between existing reserves or by restoring land between existing habitat fragments.
Many contemporary efforts to create corridors are small-scale; they can be as simple as hedgerows that connect woodlots, a strategy that likely works for some small species. Other efforts are far more ambitious. One of the earliest large efforts involved a plan to connect various parks and other protected areas in Florida by corridors of land that would have to be purchased or otherwise protected from development. From this initial effort, a conservation group, the Wildlands Project, has developed an extensive set of plans for many areas in North America intended to set priorities for the acquisition of land for a mosaic of corridors that would eventually link together large parts of the continent.
A regional example of corridor creation is in Costa Rica. Situated in the Caribbean lowlands, La Selva Biological Station is one of the major centres for research on tropical forests. Occupying an area of about 16 square km (6 square miles), the station is bordered on the south by Braulio Carrillo National Park, a much larger area of forest covering 460 square km (180 square miles). The national park extends to La Selva through a forest corridor that descends in elevation from nearly 3,000 metres (nearly 10,000 feet) at Barva Volcano down to 35 metres (115 feet) above sea level at La Selva. The corridor had been threatened by agricultural development until conservation groups, realizing that La Selva would become an isolated forest “island,” purchased the corridor to protect it.
Habitat management
Once protected, areas must often be managed in order to maintain the threatened species within them. Management may involve the removal of alien species, as previously discussed. It can also involve restoring natural ecological processes to the area. Original fire and flooding regimes are examples of such processes, and they are often controversial because human actions can alter them significantly.
Fire control
Despite the often valid reasons for suppressing wildfires (forest fire), the practice can change vegetation dramatically and sometimes harm species in the process. As previously noted, human activities have changed fire regimes across large areas of the planet, including some biodiversity hot spots. Getting the fire regimes right can be essential for conserving species.
An example of a species for which the control of fire regimes has proven both possible and essential is Kirtland's warbler (Dendroica kirtlandii; see woodwarbler (wood warbler)). This endangered species nests only in the Upper Peninsula of Michigan, an exceptional case of a bird species with a tiny geographic range well outside the tropics. The bird places its nest in grasses and shrubs below living branches of jack pines (Pinus banksiana) that are between 5 and 20 years old. The region's natural wildfires originally maintained a sufficient area of young jack pines. As elsewhere, modern practices suppressed fires, and the habitat declined. Active management of fires to ensure that there are always jack pines of the right age has steadily increased the population of warblers since the early 1990s, when it had included only about 200 singing males. By the early years of the 21st century, there were about 1,300 singing males.
Flood control
In much the same way that human actions suppress fire regimes, they also control water levels, and the resulting changes can have important consequences for endangered species. An example of a species so affected is the Cape Sable seaside sparrow (Ammodramus maritimus mirabilis) found in the Florida Everglades. The Everglades once stretched from Lake Okeechobee in the north to Florida Bay in the south. Water flowed slowly over a wide area, and its levels varied seasonally: summer rains caused the levels to rise in late summer and early fall; then the dry season dropped the levels to their lowest in late May. Under this natural regime, some prairies were continuously flooded for many years, drying out in only the driest years, whereas others were flooded for only a few months each year. It is in these drier prairies that the sparrows nest from about the middle of March until the water floods their nests in the summer.
Water-management actions have diverted the flow of water to the west of its natural path, making the western part of the Everglades unnaturally wet in some years during the bird's nesting season. Unnatural flooding during the four years that followed 1992 reduced the bird population in the west to less than 10 percent of its 1992 level. In the east the prairies have become unnaturally dry and so have become subject to an increased number of fires, which has jeopardized the sparrow population there. It is clear that rescuing the Cape Sable seaside sparrow requires the restoration of the natural water levels and flows within the Everglades and the consequent return to natural fire regimes.
Habitat restoration
Once a habitat has been destroyed, the only remaining conservation tool is to restore it. The problems involved may be formidable, and they must include actions for dealing with what caused the destruction. Restorations are massive ecological experiments; as such, they are likely to meet with different degrees of success in different places. Restoration of the Everglades, for example, requires restoring the natural patterns of water flow to thousands of square kilometres of southern Florida.
A case history of habitat restoration comes from the Midwestern United States. In Illinois, natural ecosystems cover less than 0.1 percent of the state, so restoration is almost the only conservation tool available. North Branch is a 20-km (12-mile) strip of land running northward from Chicago along the north branch of the Chicago River. Early in the 20th century, it was protected from building but later abandoned. Beginning in the 1970s, a group of volunteers first cleared out introduced European buckthorn (Rhamnus cathartica), removed abandoned cars, and planted seeds that they had gathered from combing the tiny surviving remnants of original prairies in places such as along railways and in cemeteries. The initial effort to replant the prairie species failed, however; animals ate the small growing plants. The solution was to restore the natural fire regime, although controlled burns—setting fires safely near homes—posed a difficult technical challenge. But once it was accomplished, the results were immediate and dramatic. The original prairie plants flourished and the weeds retreated, although with an important exception. Under the native trees grew nonnative thistles and dandelions.
The original habitats locally called barrens constituted a visually striking and ecologically special habitat. Restoring them was a particular challenge, and the main conservation problem was finding the right mix of species. One recommendation was to use remnant barrens as models, but the North Branch volunteers rejected them as being too degraded. In the early 20th century, naturalists had speculated that barrens were special because they lacked some characteristic prairie species and, at the same time, had their own distinctive species. The volunteers examined habitat descriptions in old treatises of local plants looking for such species, many of which certainly would now be scarce. The key discovery was a list of barrens plants published by a country doctor in 1846. The scientific names had changed in 150 years, but, by tracking them through the local literature, the volunteers found many of their putative barrens-specific plants on the doctor's list. In possession of this vital information, the group succeeded in restoring barrens sites that by 1991 held 136 native plants, including whole patches consisting of species that, until restoration, had been locally rare.
The example above illustrates two rules of ecological restoration. The first is that one must be able to save all the component species of the original ecological community. The second, which is illustrated by the doctor's list, is that one needs to know which species belong and which do not. Without the right species mix, restorers must constantly weed and reseed.
Putting a price on conservation
 Estimating the economic (economics) value of biodiversity is controversial. The value of whales, for example, once would have been just the price that traders would have paid for their parts, mostly oil and baleen. Today whales have an obvious economic value that comes from what tourists spend to watch them from boats—vessels that often now sail from the same ports that once supported whaling. Less obvious as an economic factor are the deeply held beliefs that hunting whales is cruel or that whales have a right to exist. Many people throughout the world object to whaling even though they may rarely or never have seen whales. Economists incorporate these ideas into their calculations of value by asking how much the public would pay to protect whales.
Estimating the economic (economics) value of biodiversity is controversial. The value of whales, for example, once would have been just the price that traders would have paid for their parts, mostly oil and baleen. Today whales have an obvious economic value that comes from what tourists spend to watch them from boats—vessels that often now sail from the same ports that once supported whaling. Less obvious as an economic factor are the deeply held beliefs that hunting whales is cruel or that whales have a right to exist. Many people throughout the world object to whaling even though they may rarely or never have seen whales. Economists incorporate these ideas into their calculations of value by asking how much the public would pay to protect whales.A relevant real-world example is found in the aftermath of the oil spill from the tanker Exxon Valdez, which ran aground in Prince William Sound, Alaska, in 1989. When the spill polluted the spectacular scenery of Alaska's coast, many Americans felt that they had suffered a personal loss. Exxon (Exxon Corporation) was forced to pay for cleaning up the mess and was fined not only for harming people's livelihoods but also for incurring that loss.
Whereas the propriety of decisions that put monetary values on moral issues is contentious, in 1997 a team of ecologists and economists attempted to do just that on a grand scale when they reported their calculations of how much the diverse services provided by Earth's ecosystems are worth. Their approach was to group these services into 17 categories and to classify the planet's surface into 16 ecosystem types. Many combinations of services and ecosystem types were not estimated. Mountains, the Arctic, and deserts certainly have cultural values and devoted ecotourists, but these were absent, as were urban areas with city parks. Nonetheless, the total value of $33 trillion per year at which the team arrived is roughly twice the value of the global economy.
A challenge for today's society is to comprehend the economic values of Earth's ecosystems, for often they are taken for granted. One effort to do this is found in the steps taken by New York City to improve its water supply by protecting the watersheds that supply that water. The cost of this protection was small compared with the alternative of greatly expanding the city's system of water-treatment plants. Another example, aimed at slowing the rate of deforestation, can be seen in the Costa Rican government's willingness to pay for protecting forest watersheds. A global example comprises efforts to reduce carbon emissions worldwide by rewarding those who protect existing forests or restore forests on deforested land.
Governments and international agencies often encourage the destruction of biodiversity by providing financial subsidies (subsidy) in the interests of economic growth. For example, many countries massively support their national fishing fleets by subsidizing the price of fish. The Soviet Union was such a country, but on its breakup the Russian government withdrew the subsidies, with the result that much of the fishing fleet has rusted in harbour. A study conducted in the late 20th century by the Food and Agriculture Organization of the United Nations estimated that the total annual value of the world fish catch was about $70 billion. Yet the cost of catching these fish was $92 billion, reflecting a loss of $22 billion. The cost included routine maintenance, fuel, insurance, supplies, and labour. The magnitude of the loss was certainly an underestimate, for any private company would also include in its total cost the annual costs of paying off the loans for the fishing boats and other facilities. Those loans, the study concluded, would account for another $32 billion, making the annual deficit $54 billion per year.
Agricultural subsides worldwide are even larger—a global review of all subsidies estimates that they total trillions of dollars per year. The review suggests that a powerful strategy for conservation would be to examine whether these subsidies are sensible given concerns about the environment and the need to be fiscally responsible.
Additional Reading
General works
Martha J. Groom, Gary K. Meffe, and C. Ronald Carroll, Principles of Conservation Biology, 3rd ed. (2006); and Richard B. Primack, Essentials of Conservation Biology, 4th ed. (2006), are introductions to the subject. Michael E. Soulé (ed.), Conservation Biology: The Science of Scarcity and Diversity (1986); and David Western and Mary C. Pearl (eds.), Conservation for the Twenty-first Century (1989), compile the ideas and experiences of several dozen leading conservation scientists.
Assessments of biodiversity
Stuart L. Pimm, The World According to Pimm: A Scientist Audits the Earth (2001; also published as A Scientist Audits the Earth, 2004), gives an overview of the state of human activity on Earth's terrestrial, freshwater, and marine regions and discusses the effects of this activity on biodiversity. Stuart L. Pimm et al., “The Future of Biodiversity,” Science, 269(5222):347–350 (July 21, 1995), estimates past, present, and future rates of extinction. Extensive tables and figures on the distribution and status of biodiversity are provided in Brian Groombridge and Martin D. Jenkins, World Atlas of Biodiversity: Earth's Living Resources in the 21st Century (2002). N.J. Collar, M.J. Crosby, and A.J. Stattersfield, Birds to Watch 2: The World List of Threatened Birds (1994), describes the fate of the roughly 1,100 species of birds that are globally threatened; because birds are particularly well-known, the book represents the most complete discussion of species threatened with extinction and the underlying causes of the threat. Sandra Postel, Last Oasis: Facing Water Scarcity (1992); and Norman Myers, The Primary Source: Tropical Forests and Our Future, 2nd ed. (1992), analyze the fate of freshwater ecosystems and of tropical forests, respectively. The state of fish stocks within the United States and its territories is evaluated in National Marine Fisheries Service, Report to Congress: Status of Fisheries of the United States (annual). Callum M. Roberts et al., “Marine Biodiversity Hotspots and Conservation Priorities for Tropical Reefs,” Science, 295(5558):1280–84 (Feb. 15, 2002), maps out the biodiversity of coral reefs and the factors involved in their harm.
Specific causes of species loss
Robert L. Peters and Thomas E. Lovejoy (eds.), Global Warming and Biological Diversity (1992), explores the likely consequences of global warming on biodiversity. The damage to marine ecosystems by overfishing receives major treatment in Mark Kurlansky, Cod: A Biography of the Fish that Changed the World (1997); and Carl Safina, Song for the Blue Ocean: Encounters Along the World's Coasts and Beneath the Sea (1998). Daniel Simberloff, Don C. Schmitz, and Tom C. Brown (eds.), Strangers in Paradise: Impact and Management of Nonindigenous Species in Florida (1997); and Stuart L. Pimm, “Species that Need No Introduction,” Yearbook of Science and the Future, pp. 200–219 (1994), are reviews of introduced species and their role in reducing biodiversity.
Vulnerabilities of particular species
Rosie Woodroffe and Joshua R. Ginsberg, “Edge Effects and the Extinction of Populations Inside Protected Areas,” Science, 280(5372):2126–28 (June 26, 1998), shows that the foraging behaviour of some large predators, such as African wild dogs, puts them at particular risk of local extinction. Ronald L. Westemeier et al., “Tracking the Long-Term Decline and Recovery of an Isolated Population,” Science, 282(5394):1695–98 (Nov. 27,1998), demonstrates that the decline of genetically inbred small populations can be reversed by introducing individuals from genetically diverse larger populations.
Priority setting and habitat restoration
Stuart L. Pimm and John H. Lawton, “Planning for Biodiversity,” Science, 279(5359):2068–69 (March 27, 1998), reviews technical approaches to selecting the economically and ecologically most-effective areas to be protected for their biodiversity. William K. Stevens, Miracle Under the Oaks: The Revival of Nature in America (1995), describes the efforts to restore prairies in the American Midwest.
Economic and legal issues in the conservation of biodiversity
Robert Costanza et al., “The Value of the World's Ecosystem Services and Natural Capital,” Nature, 387(6630):253–260 (May 15, 1997), calculates the annual value of the ecosystem services provided by the environment. Norman Myers and Jennifer Kent, Perverse Subsidies: How Tax Dollars Can Undercut the Environment and the Economy (2001); Geoffrey Heal, Nature and the Marketplace: Capturing the Value of Ecosystem Services (2000); and Gretchen C. Daily and Katherine Ellison, The New Economy of Nature: The Quest to Make Conservation Profitable (2002), discuss the economic factors that harm biodiversity and the development of economic tools to save it. National Research Council (U.S.), Committee on Scientific Issues in the Endangered Species Act, Science and the Endangered Species Act (1995), examines the factors involved in U.S. law in protecting endangered species.
- Schönborn, Friedrich Karl, Graf (count) von
- Schönbrunn, Schloss
- Schönbrunn, Treaty of
- Schönemann, Johann Friedrich
- Schönerer, Georg, Ritter von
- Schönherr, Karl
- Schönlein, Johann Lukas
- Schücking, Levin
- Schütz, Heinrich
- Sciacca
- Sciascia, Leonardo
- sciatica
- sciatic nerve
- Scicli
- Scidmore, Eliza Ruhamah
- science
- Science and Industry, Museum of
- science fiction
- science, history of
- Science Museum
- Science, Museum of
- science, philosophy of
- Sciences, Academy of
- Scientific American
- bunting