脱氧核糖核酸(卷名:生物学)
deoxyribonucleic acid
由成千上万个脱氧核苷酸通过磷酸二酯键连接而成的一类核酸。因含脱氧核糖而得名,简称DNA(见彩图)。
 它是染色体的主要成分。此外,真核生物的线粒体和叶绿体中也含有 DNA,原核生物的质粒全由DNA构成。在不含核糖核酸(RNA)的病毒和噬菌体中,其核酸都是DNA。
它是染色体的主要成分。此外,真核生物的线粒体和叶绿体中也含有 DNA,原核生物的质粒全由DNA构成。在不含核糖核酸(RNA)的病毒和噬菌体中,其核酸都是DNA。除了RNA病毒和噬菌体外,DNA是所有生物的遗传物质基础。生物体亲子之间的相似性和继承性即所谓遗传信息都贮存在 DNA分子中。1944年O.T.埃弗里等人从带荚膜光滑菌落的Ⅲ型肺炎球菌抽提出DNA,加到不带荚膜粗糙菌落的肺炎球菌培养基中,使后者转化为带荚膜光滑菌落的Ⅲ型肺炎球菌,并证明这种转化能力不能为蛋白水解酶或核糖核酸酶破坏,但若用脱氧核糖核酸酶处理,就失去转化能力。1952年A.D.赫尔希和M.蔡斯用同位素分别标记(32P标记DNA,35S标记蛋白质)的T2噬菌体感染大肠杆菌,发现只有DNA进入细菌体内,蛋白质则留在体外。新生成的噬菌体也只含有32P而不带35S,进一步证实DNA是遗传信息的载体。
大小和形状 最小的如病毒和噬菌体 DNA,分子量d也在百万以上,大肠杆菌的DNA分子量为2.5×109,人的DNA为1.5×1012。DNA的大小还可以所含碱基对数目和分子长度来表示,如猴肾病毒40的DNA含有5100碱基对,其分子长为1.7微米,即长度为1微米的DNA相当于3000碱基对,其分子量为3000×660或2×106(每一碱基对分子量以660计)。
DNA分子大多是线性,不分枝,如真核生物染色体中的DNA;有些是环状分子,如大肠杆菌的DNA,线粒体DNA,叶绿体DNA和一些病毒DNA等。绝大多数DNA是双链,只有少数噬菌体和病毒DNA是单链,如ΦX174的DNA是环状单链分子。
碱基组成 由脱氧腺苷酸、脱氧鸟苷酸、脱氧胸苷酸和脱氧胞苷酸等脱氧核苷酸所组成。其中腺、鸟即腺嘌呤(A)和鸟嘌呤(G);胸、胞即胸腺嘧啶(T)和胞嘧啶(C)。 测定这4种碱基的克分子比也就代表了4种脱氧核苷酸的克分子比。分析了许多不同生物材料,如人肝、牛胸腺、 麦胚、酵母、大肠杆菌等的DNA中碱基的克分子比,发现嘌呤碱克分子含量和嘧啶碱克分子含量近似相等,腺嘌呤克分子含量和胸腺嘧啶克分子含量近似相等,鸟嘌呤克分子含量和胞嘧啶克分子含量近似相等(在单链的 DNA中没有这些规律)。例如酵母中鸟嘌呤、腺嘌呤、胞嘧啶、胸腺嘧啶的克分子百分数分别为18.3,31.7,17.4,32.6。此外还发现每一种生物DNA的碱基比例都是恒定的,不因年龄、生长条件、环境因素而有变化,而且在高等生物中,不同器官和不同组织的DNA都含有相同的碱基组成。此外,每个生物DNA的腺+胸与鸟+胞的比例各异,在细菌中这个比例的范围很广,例如藤黄叠球菌为0.35,产气梭状芽孢杆菌为 2.7。但在高等生物中,这个变化就较小,动物一般为13~1.5,植物为1.1~1.7。
一级结构 也称化学结构,即核苷酸的排列顺序。至1983年中期已经测定了许多种含几千个核苷酸的 DNA的一级结构,主要有病毒、噬菌体、质粒和某些基因的DNA。几万对核苷酸的DNA如人的线粒体DNA,牛线粒体DNA和λ噬菌体DNA的一级结构也已测出。这样迅速的发展,是由于:①发现了限制性核酸内切酶,从而可以把大分子DNA在特定的部位切成许多片段,再分别进行结构测定;②测定方法有很大进展,特别是化学降解法(用特殊的化学试剂专一性地作用于A、G、C、T)和末端终止法(用2',3'-双脱氧核苷三磷酸代替正常的3'-脱氧核苷三磷酸以使 DNA链在酶促合成时停止延伸)起了很大作用;③运用重组DNA的方法获得大量的纯DNA;④新技术的应用,如用高比放射性的32P同位素标记DNA片段,使样品大为节省;又如用电子计算机帮助排列各酶解片段的先后。中国学者洪国藩在此基础上,于1982年又创造出一种系统地非随机测定法,进一步提高了效率。
高级结构 1953年,J.D.沃森和F.H.C.克里克根据DNA分子中碱基组成的规律性,DNA的X射线衍射图谱以及其他一些实验结果,提出了DNA的双螺旋模型(见脱氧核糖核酸双螺旋)。
这个模型的特征是:① DNA由两条多核苷酸链构成,这两条链的方向相反,即一条链的5'-末端和另一条链的3'-末端相对应,两条链都以右手螺旋方式盘绕同一中心轴成双螺旋结构,这样的旋转形成了大沟和小沟;②脱氧核糖和磷酸形成的骨架在外侧,碱基在双螺旋内侧,两两配对,A对T,G对C,T对A,C对G,配对碱基之间以氢键联系,从而使两条多核苷酸链形成固定的关系。此外,堆集碱基之间的疏水作用也有助于维持 DNA的双链结构。由于DNA两条链的碱基具有严格的配对关系,通常叫它们为互补链,如果一条链的核苷酸顺序弄清楚了,另外一条链上的核苷酸顺序也就一目了然;③每对碱基处于同一平面,而和中心轴垂直,不同碱基对的平面互相平行,相邻两平面的间距为3.4埃,螺旋每转一圈的螺距为3.4埃,所以每一圈螺旋含10对碱基,螺旋的直径为20埃。DNA的双螺旋结构有3种,即A型,B型和C型,在生物体内和溶液中最常见的为B型,就是具有上述各种特征的一种类型。A型和C型与B型类似,但有微小差异(见表)。
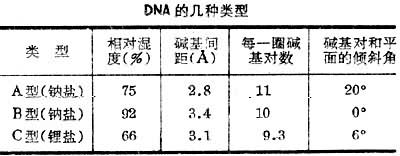
1979年以来对人工合成的dCpGpCpGpCpGp,dpCpGpCpG晶体和聚d(GC)纤维进行X射线衍射研究,发现这些化合物为左旋结构,被命名为Z型DNA。其他嘌呤和嘧啶相间的DNA片段也具有Z型的左旋结构。Z型DNA比B型DNA细而长,碱基对偏离轴心,靠近双螺旋的外侧,呈现一种比较暴露的状态,可能较易受外来因素的影响。Z型DNA还能在异体动物体内诱生抗体,用Z型DNA抗体进行实验,发现它能和天然DNA结合,说明天然DNA分子中有Z型DNA的结构。
DNA还有三级结构,如超螺旋结构,由双螺旋链扭曲而成。在原核生物内长1毫米的 DNA链能压缩在直径1微米的细胞内就是这种盘旋扭曲的结果。在真核生物细胞染色体上,DNA链与组蛋白组成核小体,在电子显微镜下呈念珠状,在一根长丝上附着很多小球,小球直径100埃左右,由长约140碱基对的DNA绕在由组蛋白H2A,H2B,H3和H4各两分子组成的八聚体蛋白质核心外面,小球之间为细丝,由DNA和组蛋白H1构成。核小体进一步旋绕折叠压缩而成染色质(见染色体)。
变性和复性 使刚性的 DNA双链解开成易于柔折的单链的现象叫做变性。凡能破坏氢键和改变堆集碱基的疏水性的试剂和条件,都能使DNA变性,如尿素、甲酰胺等试剂,酸或碱,以及加热等。随着变性,DNA出现一系列性质的变化,如紫外光吸收的增加即增色效应,旋光的减低,粘度的下降,沉降速度增加,浮力密度上升等。由于GC之间可以形成3个氢键,AT之间只能形成2个氢键,所以DNA分子中G+C的克分子含量愈高,变性愈难。利用DNA变性后的增色效应研究变性过程是最常用的手段。以温度对紫外光吸收作图得一 S形曲线,在一个相当窄的温度范围内,增色效应有一个跳跃,类似于结晶的融化。使50%的DNA变性的温度就叫做DNA的融点,通常写作Tm。Tm的高低和DNA分子中G+C的克分子含量有关,可由下式表示:
去除变性条件以后,变性DNA的2条互补链可以重新结合,恢复到原来的双螺旋,这个过程称为复性。复性的速度在一定条件下,和DNA的大小成反比例,如大肠杆菌的DNA比λ噬菌体DNA大80倍,大肠杆菌DNA的复性速度比λ噬菌体DNA小80倍。真核生物DNA更大,变性的真核生物DNA的复性速度相应应该更慢,但小鼠DNA中有约10%的复性速度比已知最小的噬菌体 DNA还要快。原因是原核生物DNA核苷酸顺序是不重复的,而真核生物DNA的核苷酸顺序有不重复的、中度重复的和高度重复的。上述小鼠 DNA的10%复性很快的部分就是高度重复顺序。不重复的顺序大都是编码蛋白质的结构基因。中度重复顺序大部分分散存在于不重复顺序之间,可能对基因表达起着调节控制作用;小部分是连续排列,主要是rRNA的基因,tRNA的基因和某些蛋白质的基因。高度重复顺序的功能还不清楚。
复制 生物为了维持种族的延续,必须把遗传信息稳定不变地传给下一代,这就靠 DNA的忠实复制来实现。以双链DNA为遗传物质的生物在DNA复制时,将亲代DNA的两条链分别作为模板,通过碱基配对(A配T,G配C)合成两条新生链,然后将两个各含一新一旧的双链DNA分配到两个子代细胞中,这种复制方式便称为半保留复制。复制是一个极为复杂的过程,除需DNA聚合酶外,还有多种酶或蛋白质因子参与(见脱氧核糖核酸的复制)。
损伤和修复 环境中化学的或物理的因素以及生物体内代谢过程中产生的自由基均可对 DNA造成损伤。如:酸和热能使嘌呤碱基脱落;碱基脱氨能使DNA复制时产生差错;有些化合物如吖啶嵌入DNA碱基中时,可引起复制和重组的误差(插入或缺失一个核苷酸),而产生移码突变。电离辐射或化学物质(如博莱霉素)能引起磷酸二酯键断裂。紫外光能使 DNA分子中相邻二嘧啶(特别胸腺嘧啶)形成环丁基二聚体,而妨碍了嘧啶通过氢键形成碱基对。
DNA 修复的分子机制目前了解得最多的是对嘧啶二聚体的修复。主要通过两个途径:①酶促光激活作用。生物体内有一个能专一地和嘧啶二聚体结合的酶,当酶和二聚体结合后,在可见光照射下,光能可使二聚体解开恢复成嘧啶单体;②切除修复。在大肠杆菌中,先由一特殊的内切酶在邻近胸腺嘧啶二聚体处切开,然后由聚合酶Ⅰ的5'→3'外切酶活力将带有二聚体的寡核苷酸片段除去,同时通过聚合酶Ⅰ的作用(从切断处的3'-端开始)重新合成缺损的片段,最后再经过DNA连接酶的作用将新合成片段和DNA的其余部分连接起来。
如果缺乏这种修复机制,可造成一些疾病,如人的着色性干皮病,就是因为不能修复由于紫外光造成对DNA的损伤所致。患有这种病的人对阳光非常敏感,容易生皮肤癌(见DNA损伤修复)。
修饰和限制 细菌有一种不降解自身DNA但能降解异源DNA的机制,这是阻止异源DNA在体内起作用的一种保护作用。起这个保护作用的就是限制性核酸内切酶。限制性核酸内切酶有Ⅰ型和Ⅱ型两类:Ⅰ型酶需Mg2+、ATP和S-腺苷甲硫氨酸;Ⅱ型酶只需Mg2+;Ⅰ型酶除有核酸酶活力外,还有甲基化酶和腺三磷酶活力。Ⅱ型酶则只有核酸酶活力;Ⅱ型酶的切点是专一的,它要求一定的核苷酸顺序(具有双重旋转对称性),而Ⅰ型酶则否。
如果碱基被修饰了,如腺嘌呤的6-氨基上的氢原子被甲基取代或胞嘧啶的5位甲基化了,原来能裂解这个核苷酸顺序的限制性核酸内切酶就不能再作用于这个修饰过的DNA。因此,如果细菌本身的DNA是修饰过的(甲基化),则限制性核酸内切酶对自身DNA不起作用,当外来DNA(未修饰过的)进入细菌体内,限制性核酸内切酸可将其降解,这样对自身就起到了保护作用。
由于Ⅱ型限制性核酸内切酶作用于DNA时,需要特定的核苷酸排列顺序,因此它是测定DNA核苷酸排列顺序的非常有用的工具。目前 DNA核苷酸顺序的测定之所以得到极为迅速的发展,Ⅱ型限制性核酸内切酶起了很大作用。
限制性核酸内切酶广泛存在于各种细菌内,但在真核生物中似乎尚未找到过。至1983年已搞清识别顺序的Ⅱ型限制性核酸内切酶达400种。示例如下:
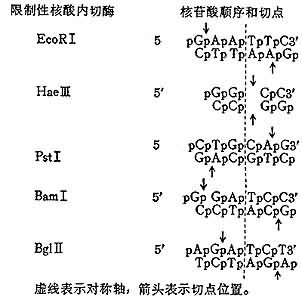 。
。DNA重组 同源DNA在体内重组是经常发生的。这是产生生物遗传性多样化的主要原因。 DNA重组可通过人工方法实现。即将DNA片段在体外连接,然后引入活细胞内进行复制,要完成这项工作须具备下列条件:①DNA运载体(如质粒、噬菌体、病毒等)能在活细胞内复制;②需要复制的DNA片段即DNA乘客(如各种基因);③连接DNA片段和DNA运载体的方法和工具酶(如限制性核酸内切酶、DNA连接酶等);④将连接好的DNA(重组体DNA)引入寄主细胞的方法(如转化、转染等);⑤筛选带有所需重组体DNA的细胞的方法。
重组体 DNA是20世纪70年代才开展起来的研究。短短10年利用这个方法已经生产了许多多肽和蛋白质激素、酶制剂、疫苗、干扰素、抗生素等。
人工合成 由于重组DNA技术的建立,人工合成DNA的工作有了非常迅速的发展。到1983年,已合成了tRNA、胰岛素A、B链、胰岛素原,生长激素释放抑制因子,胸腺素α、干扰素α和γ、表皮生长因子,生长激素类胰岛素生长因子、 促胰激素、 降钙素、内啡肽等的基因(见核酸的人工合成)。
参考书目
王德宝、祁国荣:《核酸结构功能与合成》,科学出版社,北京,1986,1987。
R.L.P.Adams et al., The Biochemistry of Nucleic Acids,9 th ed., Chapman and Hall Ltd.,London,1981.
E.L.Smith et al., Principles of Biochemistry, General Aspects,7th ed., McGraw-Hill Book Co.,New York,1983.