细胞周期(卷名:生物学)
cell cycle
以有丝分裂方式增殖的细胞从一次分裂结束到下一次分裂结束所经历的过程。这一过程周而复始。
细胞周期是50年代细胞学上重大发现之一。在这之前认为有丝分裂期是细胞增殖周期中的主要阶段,而把处于分裂间期的细胞视为细胞的静止阶段。
1951 年霍华德等用32P-磷酸盐标记了蚕豆根尖细胞,通过放射自显影研究根尖细胞DNA合成的时间间隔,观察到32P之掺入不是在有丝分裂期,而是在有丝分裂前的间期中的一段时间内。发现间期内有一个DNA合成期(S期),32P只在这时才掺入到DNA;S期和分裂期(M期)之间有一个间隙无32P掺入,称为G2期,在M期和S期之间有另一个间隙称为G1期,G1期也不能合成DNA。
于是他们提出了细胞周期的概念,并首先证明间期是细胞周期中极为重要的一个阶段,发生着许多与细胞分裂有关的特殊生化事件。这一发现被以后学者们用3H-胸腺嘧啶核苷进行的类似研究所证实。
细胞生命活动大部分时间是在间期度过的,如大鼠角膜上皮细胞的细胞周期内,间期占14000分钟。分裂期仅占70分钟。细胞周期各阶段都有复杂的生化变化。间期是细胞合成DNA、RNA、蛋白质和各种酶的时期,是为细胞分裂准备物质基础的主要阶段。
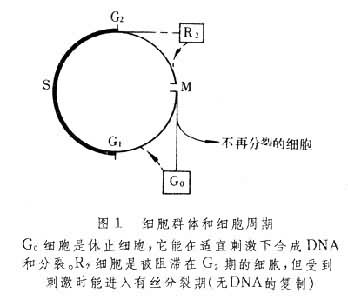
在一个增殖的细胞群中,所有细胞并非是同步增殖的,它们在细胞周期运行中,可能有四种命运(图1):①细胞经M期又开始第二次周期;②停止于G2期,称为G2期细胞(R2),它受某种刺激后可进入周期;③停止在G1期,称为休止细胞或名G0期细胞,这类细胞受某种刺激仍能进入周期,并开始DNA合成和有丝分裂;④丧失生命力近于死亡的细胞,称为丢失细胞,或称不再分裂的细胞。继续分裂的细胞沿着细胞周期从一个有丝分裂期到下一个分裂期。不再分裂的细胞离开了细胞周期不再分裂,最终死亡。
G1期 进行大量物质合成时期。细胞体积逐渐增大,制造RNA(包括tRNA,mRNA,rRNA以及核糖体等)。RNA的合成又导致结构蛋白和酶蛋白的形成,这些酶又控制着形成新细胞成分的代谢活动。G1又分为G1早期和G1晚期两个阶段;细胞在G1早期中合成各种在G1期内所特有的RNA和蛋白质,而在G1晚期至S期则转为合成DNA复制所需要的若干前体物和酶分子,包括胸腺嘧啶激酶、胸腺嘧啶核苷酸激酶、脱氧胸腺嘧啶核苷酸合成酶等,特别是DNA聚合酶急剧增高。这些酶活性的增高对于充分利用核酸底物在S期合成DNA是不可少的条件。
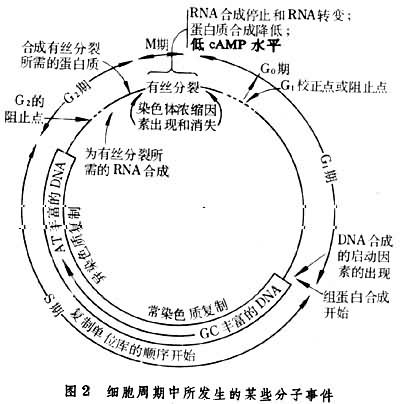
G1期持续时间变异很大,多数细胞的G1期较长,是与细胞需要增加质量有关。但在某些单细胞生物如大变形虫、四膜虫和多细胞生物的某些细胞(如海胆胚胎,小鼠胚胎细胞)则无G1期,中国仓鼠卵巢细胞的变异株无G1和G2期,以致M期和S期连接在一起。G1期的长短之所以变化很大,与G1期内存在一个校正点或阻止点(简称R点)有关。R点主要控制 G1期时间的长短。通过了此点,细胞就能以正常速度不受外界条件的影响而完成细胞周期的其他时期。因此,有人认为细胞的生长是在G1期R点上停止的,例如当细胞内环腺苷酸(cAMP)水平增高,细胞密度增加时,可阻止细胞从G1期向S期过渡,用嘌呤霉素抑制蛋白质合成或用放射线菌素D抑制RNA合成,也能延缓细胞从G1期进入S期(图2)。
有人发现 G1期内能合成一种有触发作用的蛋白质;它是不稳定的,极易被分解,故称为v蛋白。v蛋白在G1细胞中达到一定水平时,细胞便可通过R点进入S期。
G0期 细胞周期的调节主要是通过G1期的阻留而实现的,G0期即指细胞处于阻留的状态。细胞通过M期一分为二,有的可继续分裂进行周期循环,有的转入G0期。G0期是脱离细胞周期暂时停止分裂的一个阶段。但在一定适宜刺激下,又可进入周期(图1),合成DNA与分裂。G0期的特点为:①在未受刺激的G0细胞,DNA合成与细胞分裂的潜力仍然存在;②当G0细胞受到刺激而增殖时,又能合成DNA和进行细胞分裂。
S期 在这一阶段完成DNA的合成以及合成与DNA组装构成染色质等有关的组蛋白。 DNA含量在此时期增加一倍。S期终结时,每一染色体复制成两个染色单体(Hole,1979)。 生成的两个子代DNA分子与原来DNA分子的结构完全相同。一个人体细胞核直径10~20微米,其中DNA含量为10-11克,如拉成一根DNA链,长度可达3米。哺乳类动物细胞S期一般为6~8小时。DNA的复制能在几小时内完成,主要是由于DNA链分成许多的复制单位(复制子)(可多达10000个左右),它们可在S期的不同时间分别复制。另外,在S期内还有组蛋白的合成──组蛋白基因在G1-S期之间活化,组蛋白mRNA的转录增大,并在整个S期内连续进行。 已合成的组蛋白使新合成的DNA很快转为核组蛋白复合体。
S期细胞含有一种因素能诱导DNA合成,用细胞融合实验证明,G1细胞在与S期细胞融合后能加速其核内DNA复制的起点启动。S期不同阶段复制的DNA碱基组成是不同的,早期复制的DNA富有G-C碱基,晚期复制的DNA富有A-T碱基,即常染色质比异染色质复制较早(图2)。
G2期 是DNA复制结束和开始有丝分裂之间的间隙,在这期间细胞合成某些蛋白质和RNA分子,为进入有丝分裂提供物质条件。 用放射标记的RNA前体和蛋白质前体示踪,表明G2期进行着强烈的RNA和蛋白质的合成。假如破坏这些合成过程,细胞就不能过渡到M期。G2期合成的是染色体浓缩以及形成有丝分裂器所需的成分。有人认为G2期继续完成从S期就开始的微管蛋白的合成,为M期纺锤丝的组装提供原料。在G2晚期开始合成有丝分裂因子。在某些缺少G1期细胞中,G2期更为复杂,还要担负起其他细胞G1期中所要完成的事件。也有少数情况,S期结束后立即开始有丝分裂,而不存在G2期。
M期 有丝分裂时期,是细胞形态结构发生急速变化的时期,包括一系列核的变化、染色质的浓缩、纺锤体的出现,以及染色体精确均等地分配到两个子细胞中的过程,使分裂后的细胞保持遗传上的一致性。M期分为前期、中期、后期和末期(见有丝分裂)。 M期虽是形态变化最为显著的时期,但其呼吸作用反而降低,蛋白质合成明显下降,RNA合成及其他代谢周转停止,这是由于有丝分裂期所需要的能量和其他基本物质均在间期内合成和贮备好了有关。
细胞周期中,细胞形态也发生一系列变化,从光学显微镜下可看到G1期细胞最小,细胞扁平而光滑,随着向S→G2→M期的发展细胞逐渐增大,从扁平变成球形。扫描电镜下可明显看到各时期内细胞表面形态的变化,如微绒毛逐渐增加,这些变化和细胞内各种生化的和生理的周期性变化是有关的。
调控 细胞周期中的许多生化事件是按一定顺序,有条不紊地进行着,这和基因按一定顺序表达密切相关。
有人认为,细胞周期内有两个阶段最为重要:G1到S和G2到M;这两个阶段正处在复杂活跃的分子水平变化的时期,容易受环境条件的影响,如果能够人为的进行调控,将对深入了解生物的生长发育和控制肿瘤生长等有重要意义。
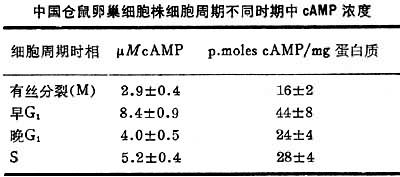
已发现很多体内因素可以激发或抑制细胞的增殖,例如多种激素、血清因子、多胺、蛋白水解酶、神经氨酶、cAMP、cGMP以及甘油二脂(DG)、 三磷酸肌醇(1P3)和Ca2+信使系统等等。 细胞内cAMP浓度增加对细胞增殖有抑制作用,凡能使细胞内cAMP增高的因素都能抑制细胞的增殖,降低细胞生长速度;反之,凡能使细胞内cAMP含量下降的因素都能促进DNA的合成与细胞的增殖。 细胞周期的各期中的cAMP含量也不相同(见表)。在中国仓鼠卵巢细胞株中,M期cAMP含量最低,M期后cAMP的水平增高三倍,从G1早期至G1晚期,cAMP水平降低到中等水平,直至S期仍维持低的水平(图3)。
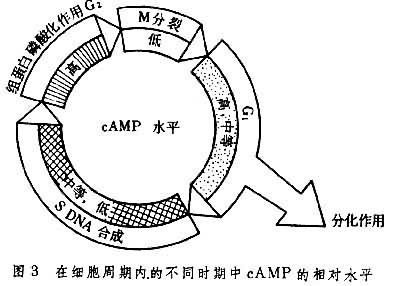
还有许多实验指出cGMP 也对细胞增殖起调控作用,如将cGMP或双丁酰cGMP加到休止在G1期的 3T3 细胞时,能诱导DNA含量的增加, 促进细胞的分裂。如提高细胞cGMP水平,就可促进细胞的有丝分裂,反过来,促进有丝分裂的药物也能增加cGMP的浓度。
cAMP能抑制细胞的分裂,促进细胞的分化,cGMP则能抑制细胞分化,促进细胞增殖,在正常生长的细胞中,cAMP和cGMP维持在适当的水平,调节控制细胞周期的运转。
抑素是细胞产生的一种小分子蛋白质或多肽,有的还含有糖或RNA。它无种属特异性,但有细胞特异性,对同类细胞增殖有抑制作用并且可逆。当抑素含量达到一定浓度时可抑制同类细胞的增殖,抑素浓度下降则细胞增殖活跃。有人认为抑素作用的机制,在于它能激活细胞膜上的腺苷环化酶活性,提高细胞内cAMP的浓度,因而抑制细胞的增殖,也可能通过cAMP-依赖性蛋白激酶对蛋白质的磷酸化作用来影响调节基因的活动。
细胞周期也受机体调节系统的影响,例如肝再生就是由调节系统的作用加速肝细胞增殖。但是肿瘤细胞,由于宿主失去对它的调控,因而恶性增殖。在肿瘤治疗中可应用细胞周期的原理,如G0期细胞对化疗不敏感,往往成为日后癌症复发的根源,因而可通过调控机理的研究,诱发G0期癌细胞进入细胞周期,再合理用抗癌药物加以杀灭,是防止癌转移和扩散的重要调控措施,也是细胞动力学中有理论意义和实践意义的研究问题。
总之,目前所了解的细胞增殖调控的分子基础还少,尚待进一步探索。