致死突变型(卷名:生物学)
lethal mutant
导致个体死亡的突变型,在高等动植物中常导致个体在性成熟前死亡。致死突变型对于致死的个体来说是不利的,但有利于维持自然群体的杂合状态。某些致死突变型的生物品系可用于个体发育的研究及育种实践,所以受到人们的重视。
简史 1905年法国学者L.居埃诺首先报道小鼠的黄色突变型从来没有育成过纯种。这种使小鼠体毛呈黄色的突变基因AY对黑色基因 a虽然是显性的,但所有黄鼠近交所产生的后代都是2黄色∶1黑色,而不是通常应出现的3∶1分离比(见孟德尔定律)。这种黄色小鼠与黑色小鼠杂交的子代则是1黄色∶1黑色。这表明黄色的小鼠是杂合体。后来的研究证明了含两个AY基因的纯合体黄色小鼠的胚胎曾在母体内出现过,但都在发育早期死亡了。此外又发现将黄色小鼠自交产生的子代个体数同黄色小鼠和黑色小鼠杂交所产生的子代个体数相比平均约少1/4。这些事实证明 AY基因有多效性:它既是一个影响毛色的显性基因,又是一个影响生活力的隐性致死基因。AY基因纯合体的胚泡植入子宫壁后不久便死亡,所以这种小鼠的黄色变种不可能育成纯种。上述杂交结果可以表示如下:
此后在异体交配的动、植物群体及人类中普遍地发现了隐性致死突变型。这种突变型在杂合状态下一般不影响个体的生活力,但纯合状态则会引起死亡。例如在植物中的隐性白苗致死突变型在纯合情况下幼苗缺乏合成叶绿素的能力,子叶中的养料耗尽就会死亡。又如人类中的一种致死性大疱性表皮松解症是常染色体隐性遗传疾病,如果隐性致死基因呈纯合状态则个体在胚胎期死亡。
类别 致死突变型按表现程度可分为全致死的(纯合体全部死亡或至少90%死亡),亚致死的(纯合体死亡率达50~90%),半致死的(纯合体死亡率在10~50%之间)和弱致死的(即类正常的,纯合体死亡率在10%以下) 几种。按环境条件对致死效应的影响又可分为条件致死的(例如温度敏感突变型在较低温度下不出现致死效应在较高温度下出现致死效应)和非条件致死的(在已知任何条件下都有致死效应)两类。按致死作用发生的阶段还可分为配子致死的(花粉死亡)和合子致死的(受精后合子不能发育)、胚胎致死的和幼体致死的。按照个体发育中致死作用时相来分,又可以分为单相致死的(只在某一时期致死)、多相致死的(致死期有两个或两个以上)、无相致死的(致死作用可以发生在任何一个时期)。
致死突变可发生在任何染色体上;发生在常染色体上的称常染色体致死,发生在性染色体上的称为伴性致死。在果蝇等性染色体属XY型的生物中,如果隐性致死突变发生在 X染色体上,对雄性果蝇即可产生致死效应;但对雌性果蝇则只有两个隐性致死突变基因纯合才会造成死亡。如果在雌性果蝇的一个 X染色体上带有致死基因1,在另一个X染色体上带有它的等位显性基因 L,雌果蝇就可以生活和产生后代。所以雌性果蝇可以是隐性致死基因的长期携带者。这种雌性杂合体(L1)果蝇和正常的雄性果蝇交配所产生的子代的雌雄比不是1♀∶1♂而是2♀∶1♂。
显性的致死突变型在杂合状态下即可致死,因此显性致死突变型只有条件致死的,半致死的或弱致死的。例如人类的Ⅱ型家族性高脂蛋白血症是常染色体显性突变型,它的主要临床特征是高血脂和多发黄色瘤,部分患者常早年死于心肌梗塞。
检测 观察杂交的子代是否出现不正常分离比、异常的雌雄比和有规律的早期夭折等都有助于发现致死突变型。1927年美国遗传学家H.J.马勒首创 C1B法用来检测果蝇伴性致死突变基因。这里1(lethal)代表X染色体上的一个隐性致死基因;B代表同一X染色体上的显性棒眼基因(bareye,B);C(crossover suppressor)代表 X染色体上一段包括1和B的倒位,它能抑制交换的发生(见染色体畸变),从而保证1和B在连续的世代中稳定地联系在一起,所以棒眼性状的存在就表明这一隐性致死基因的存在。检测方法是用诱变因子(如 X射线)处理雄性果蝇,然后将它和 C1B雌性果蝇交配,子一代会出现棒眼雌蝇,把子一代中的棒眼雌蝇与野生型雄蝇作单对交配,子二代雌雄蝇数目之比应是2∶1。但是如果在子一代的雌蝇中来自雄性亲体的 X染色体上发生了一个隐性致死突变,那么子二代中就看不到雄性果蝇(图1)。应用这一方法可以检测 X连锁隐性致死基因突变率。马勒用这种方法测得的果蝇 X连锁隐性致死自发突变率大约是0.1~0.2%。以后又发展起来的Muller-5法、骈连X染色体法等可以用来更有效地检测果蝇的X连锁隐性致死突变率。

平衡致死 马勒在1918年发现果蝇的蛎壳翅 (Bea-ded,Bd) 基因是一个隐性致死的显性形态基因。经过选择,可以得到一个只产生蛎壳翅果蝇的真实遗传品系。以后证明在这一品系中还有另一个隐性致死基因1,Bd基因与1基因分别位于一对同源染色体的不同位置上,而且还有一个能抑制Bd基因与 1基因发生交换的因子。这一真实遗传的蛎壳翅品系的基因型可以写作Bd+/+1,由于Bd和1之间不发生交换,所以Bd+/+1果蝇的后裔只可能是Bd+/Bd+、+1/+1和Bd+/+1三种。可是Bd+/Bd+和+1/+1两种是致死的,因此实际上只有后一种能够生存,而且都具有蛎壳翅表型。所以在这一平衡致死系统中,每一存活的个体都是杂合体,无须选择而能保持真实遗传(图2)。杂合状态一般有利于物种的生存(见杂种优势),而致死基因虽然对个体有害却有利于杂合性的保持。

应用 二倍体生物的致死突变型的纯合体不能生存或生活能力较低,所以一般难以保持致死基因的真实遗传品系。平衡致死系统可以用来保持致死突变型的真实遗传品系。在单倍体生物(如病毒、细菌)中,致死突变型往往无法保存。所以在这些生物中条件致死突变型对于遗传学研究有特别重要的意义,因为可以在非致死条件下保存它们,在致死条件下对它们进行研究。例如在T4噬菌体中发现的几十种温度敏感致死突变型在低温下可以在寄主细菌中增殖,然后以完整颗粒的形式释放出来。但在较高温度下某些突变型不能产生头部,另一些不能产生尾或尾丝等部分。对于这些突变型研究的结果表明T4噬菌体是通过三条装配线而形成一个成熟噬菌体的。又如在大肠杆菌中已经发现十几个在高温中不能进行DNA复制的温度敏感突变型。由于DNA是生命活动中的必要物质,所以这些突变型都是致死的,只在某一种条件下才能生存。对于这些条件致死突变型进行深入研究,将有助于对 DNA复制过程的了解。高等动植物必须通过复杂的胚胎发育过程才能成为一个个体。对各类致死突变型的研究也将有助于对发育机制的了解。
致死突变型还可以应用于育种实践,1975年B.A.斯特鲁尼科夫利用性连锁平衡致死原理育成了几乎只产雄蚕的家蚕品系。雄性家蚕的性染色体是ZZ,雌性家蚕是ZW。11和12是两个Z染色体上的致死基因,将不带有致死基因的雌蛾WZ和两个 Z染色体上分别带有致死基因11和12的雄蛾进行杂交,雌蛾所产的蚕卵中雌性蚕卵WZ(11)或WZ(12)全部不能孵化, 而雄性蚕卵++/11+或++/+12则可以孵化(图3)。
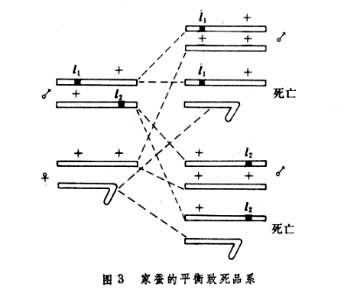
由于雄性的幼虫期短,消耗桑叶较少,出丝率较高,丝的质量较好,所以能达到增产的效果。